Anti-Inflammatory Effect of Chloroform Fraction of Pyrus Ussuriensis Maxim. Leaf Extract on 2, 4-Dinitrochlorobenzene-Induced Atopic Dermatitis in nc/nga Mice


Abstract
:1. Introduction
2. Material and Methods
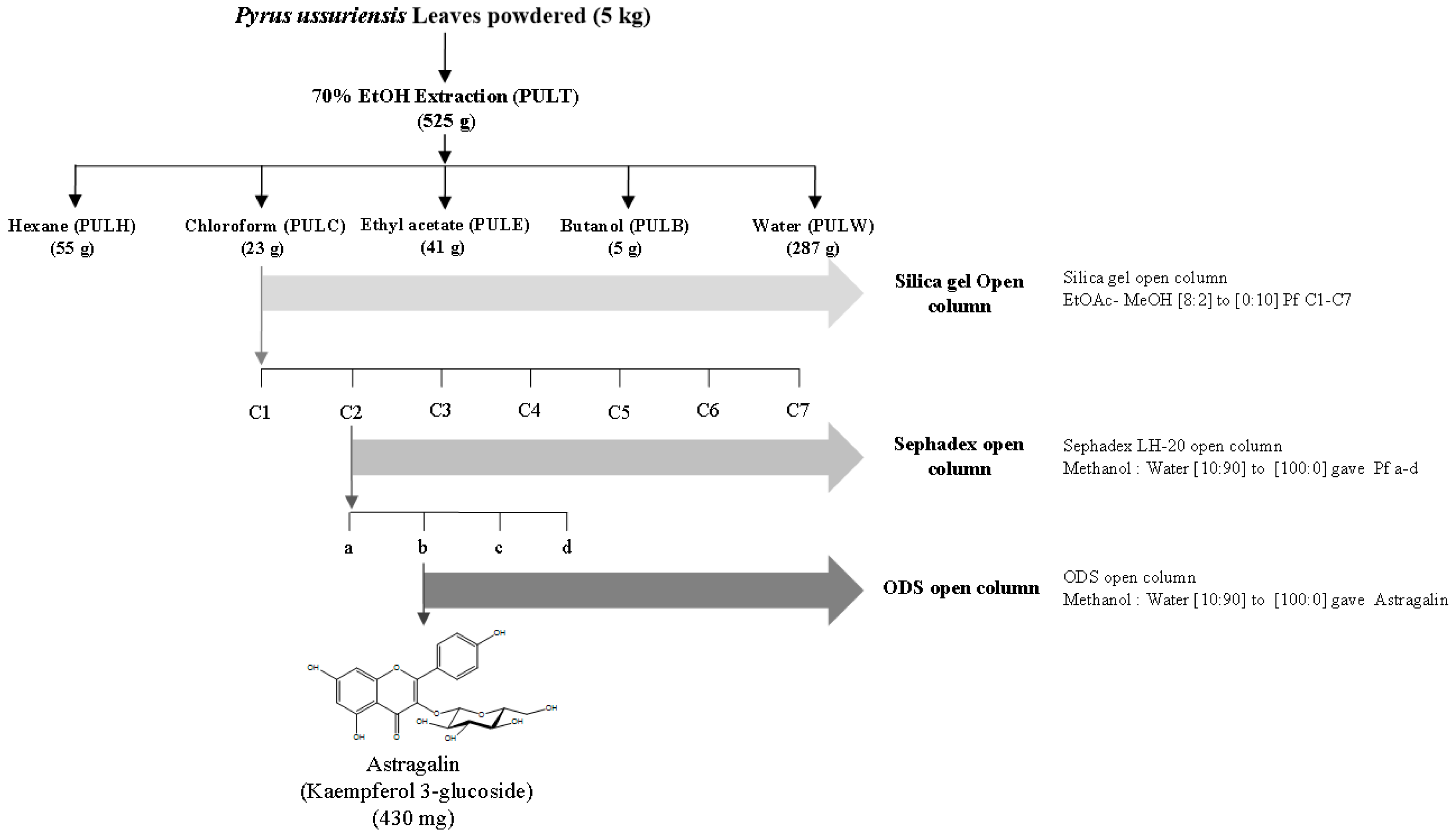
2.1. Fractionation and Isolation of Pul Extract
2.2. HPLC Analysis of PULC
2.3. Cell Culture
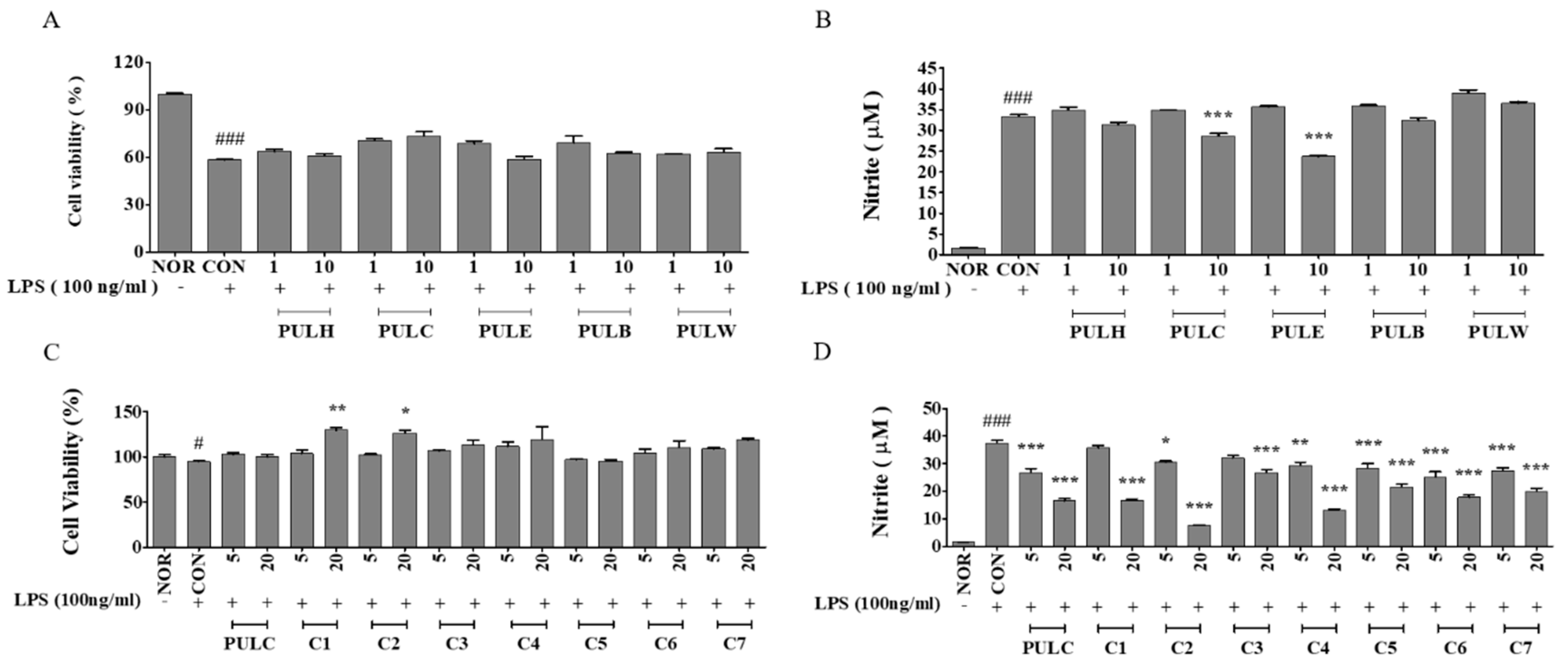
2.4. MTT Assay
2.5. Assessment of NO Production in RAW 264.7 Cells
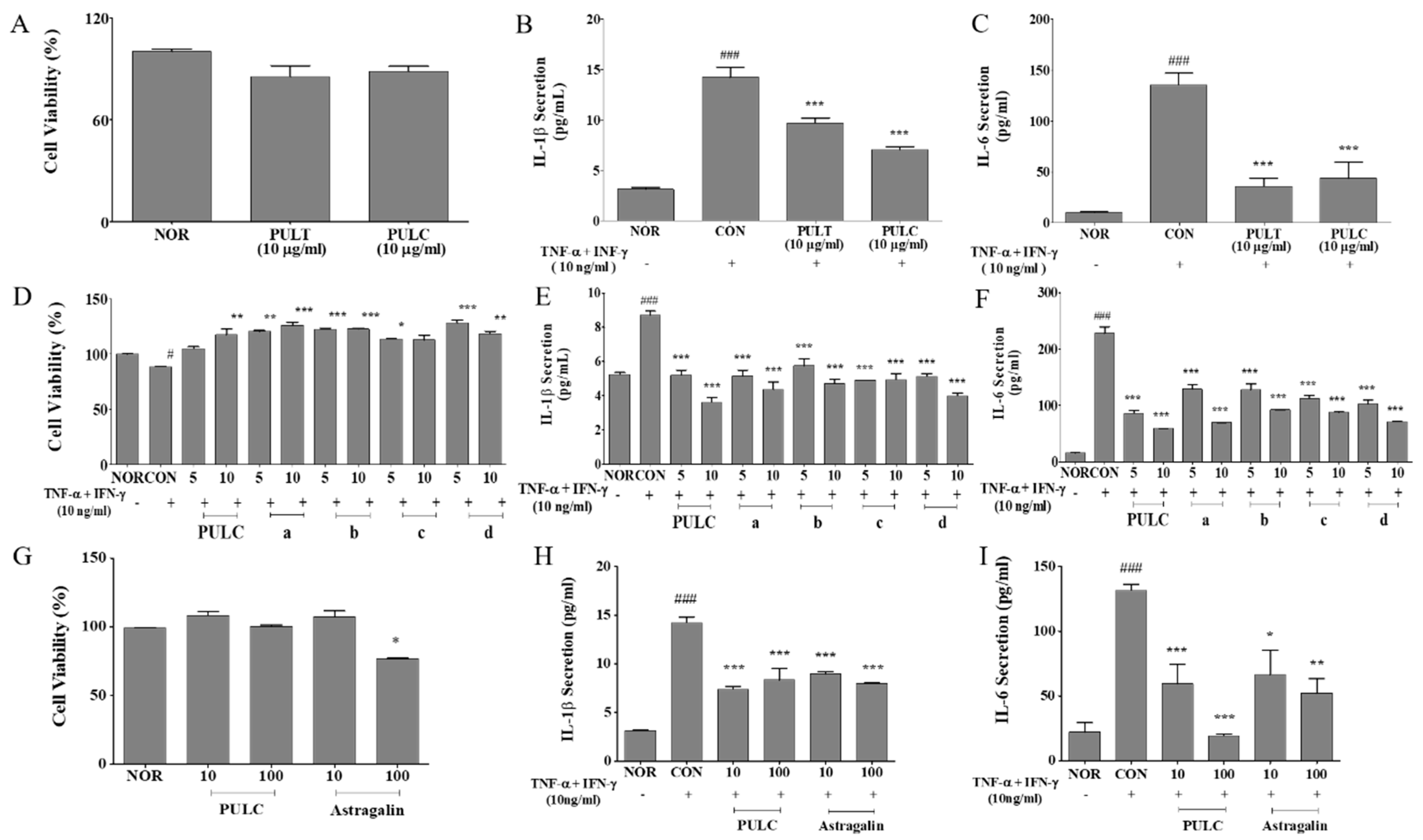
2.6. Evaluation of IL-6 and IL-1β in TNF-α/ IFN-γ-Induced HaCaT Cells
2.7. DNCB-Induced AD model
2.8. Measurement of Trans-Epidermal Water Loss (TEWL), Hydration, and Erythema
2.9. Evaluation of Skin Lesions and Dermatitis Score
2.10. Scratching Behavior Evaluation
2.11. Serum IgE
2.12. Histological Examination
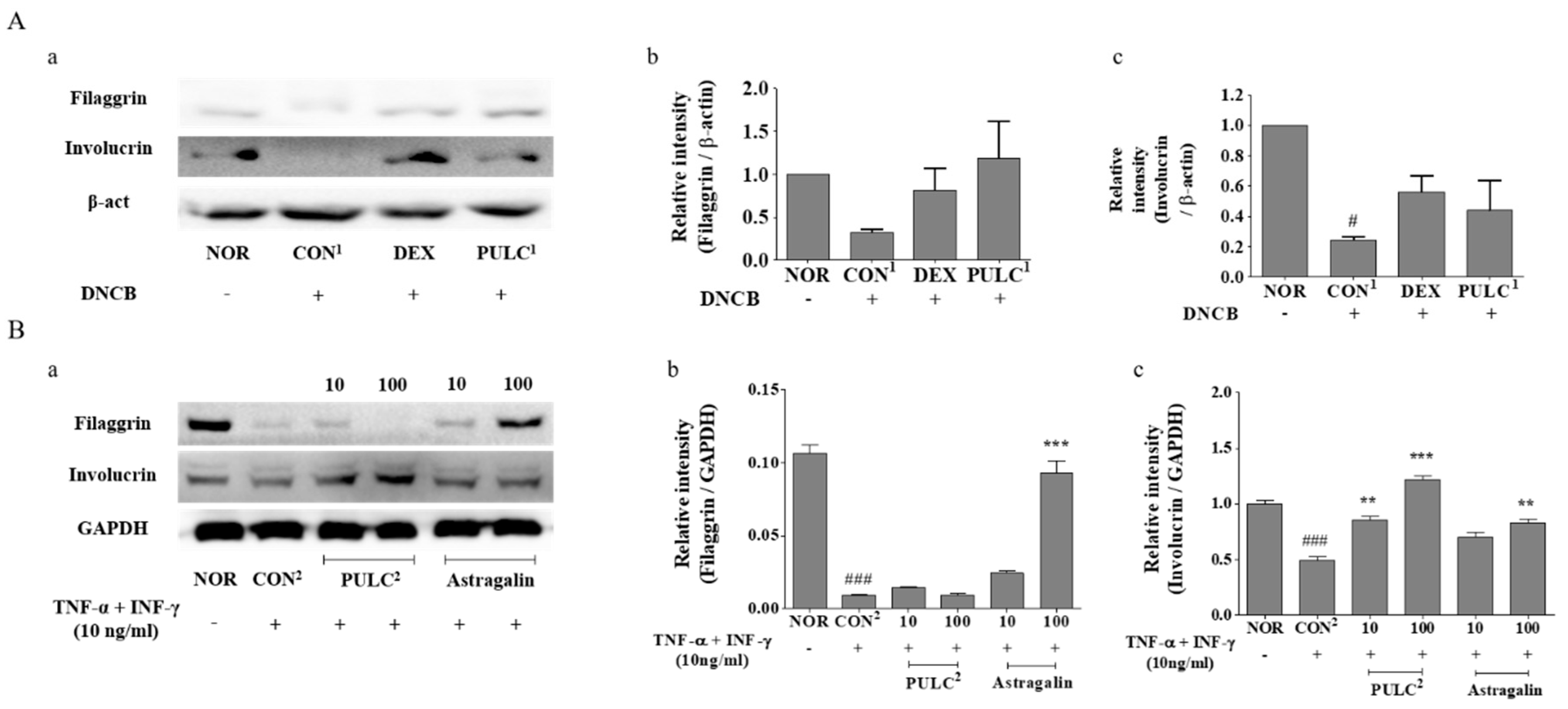
2.13. Western Blotting
2.14. Statistical Analysis
3. Results
3.1. Measurement of Astragalin Content from PULC by HPLC Analysis
3.2. Bioactivity-Guided Fractionation of PULC Based on Its Anti-Inflammatory Effects
3.3. PULC Reduced IL-6 and IL-1β Levels in TNF-α/IFN-γ Treated HaCaT Cells
3.4. Effect of PULC on Nc/Nga Mice
3.5. PULC Alleviates DNCB-Induced AD-Like Symptoms in NC/Nga Mice
3.6. PULC Alleviates DNCB-Induced Scratching Behavior in NC/Nga Mice
3.7. PULC Decreased Serum IgE Levels in DNCB-Induced NC/Nga Mice
3.8. Histological Examination
3.9. Effects of PULC and Astragalin on Protein Expressions of Filaggrin and Involucrin
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pastore, S.; Mascia, F.; Giustizieri, M.L.; Giannetti, A.; Girolomoni, G. Pathogenetic mechanisms of atopic dermatitis. Arch. Immunol. Ther. Exp. 1999, 48, 497–504. [Google Scholar]
- Novak, N.; Bieber, T.; Leung, D.Y. Immune mechanisms leading to atopic dermatitis. J. Allergy Clin. Immunol. 2003, 112, S128–S139. [Google Scholar] [CrossRef] [PubMed]
- Leung, D. Atopic dermatitis: Immunobiology and treatment with immune modulators. Clin. Exp. Immunol. 1997, 107, 25–30. [Google Scholar] [PubMed]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soter, N.A. Morphology of atopic eczema. Allergy 1989, 44, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Saklatvala, J.; Davis, W.; Guesdon, F.; Karin, M.; Marshall, C. Interleukin 1 (IL1) and tumour necrosis factor (TNF) signal transduction and discussion. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 151–157. [Google Scholar]
- Sumimoto, S.; Kawai, M.; Kasajima, Y.; Hamamoto, T. Increased plasma tumour necrosis factor-alpha concentration in atopic dermatitis. Arch. Dis. Child. 1992, 67, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.-Y.; Liu, W.; Zhang, P.; Wang, R.-Y.; Guo, J.-Y. Effects and mechanisms of aloperine on 2, 4-dinitrofluorobenzene-induced allergic contact dermatitis in BALB/c mice. Eur. J. Pharmacol. 2010, 629, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Toshitani, A.; Ansel, J.C.; Chan, S.C.; Li, S.-H.; Hanifin, J.M. Increased interleukin 6 production by T cells derived from patients with atopic dermatitis. J. Investig. Dermatol. 1993, 100, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Rowe, A.; Bunker, C. Interleukin-4 and the interleukin-4 receptor in allergic contact dermatitis. Contact Dermat. 1998, 38, 36–39. [Google Scholar] [CrossRef]
- Blume-Peytavi, U.; Wahn, U. Optimizing the treatment of atopic dermatitis in children: A review of the benefit/risk ratio of methylprednisolone aceponate. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Hengge, U.R.; Ruzicka, T.; Schwartz, R.A.; Cork, M.J. Adverse effects of topical glucocorticosteroids. J. Am. Acad. Dermatol. 2006, 54, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Gelfand, E.W. Mouse models of allergic diseases. Curr. Opin. Immunol. 2009, 21, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.G.; Cho, S.J.; Koh, H.B.; Han, D.U.; Lee, B.J. Fermented Maesil (Prunus mume) with probiotics inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice. Vet. Dermatol. 2010, 21, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Takeuchi, H.; Sakuma, S.; Sengoku, T.; Takakura, S. Characterization of a 2, 4-dinitrochlorobenzene-induced chronic dermatitis model in rats. Skin Pharmacol. Physiol. 2009, 22, 240–247. [Google Scholar] [CrossRef]
- Inagaki, N.; Nagai, H. Analysis of the mechanism for the development of allergic skin inflammation and the application for its treatment: Mouse models for the development of remedies for human allergic dermatitis. J. Pharmacol. Sci. 2009, 110, 251–259. [Google Scholar] [CrossRef]
- Takano, N.; Arai, I.; Kurachi, M. Analysis of the spontaneous scratching behavior by NC/Nga mice: A possible approach to evaluate antipruritics for subjects with atopic dermatitis. Eur. J. Pharmacol. 2003, 471, 223–228. [Google Scholar] [CrossRef]
- Ziment, I.; Tashkin, D.P. Alternative medicine for allergy and asthma. J. Allergy Clin. Immunol. 2000, 106, 603–614. [Google Scholar] [CrossRef]
- Lee, J.C.; Pak, S.C.; Lee, S.H.; Na, C.S.; Lim, S.C.; Song, C.H.; Bai, Y.H.; Jang, C.H. Asian pear pectin administration during presensitization inhibits allergic response to ovalbumin in BALB/c mice. J. Altern. Complement. Med. 2004, 10, 527–534. [Google Scholar] [CrossRef]
- Lee, C.-E.; Kim, Y.-H.; Lee, B.-G.; Lee, D.-H. Antioxidant effect of the fraction isolated from Pyrus ussuriensis Leaves. J. Korean For. Soc. 2010, 99, 885–890. [Google Scholar]
- Cho, K.; Parveen, A.; Kang, M.C.; Subedi, L.; Lee, J.H.; Park, S.Y.; Jin, M.R.; Yoon, H.; Son, Y.K.; Kim, S.Y. Pyrus ussuriensis Maxim. leaves extract ameliorates DNCB-induced atopic dermatitis-like symptoms in NC/Nga mice. Phytomed. Int. J. Phytother. Phytopharm. 2018, 48, 76–83. [Google Scholar] [CrossRef]
- Hwang, K.-A.; Hwang, Y.-J.; Song, J. Antioxidant activities and oxidative stress inhibitory effects of ethanol extracts from Cornus officinalis on raw 264.7 cells. BMC Complement. Altern. Med. 2016, 16, 196. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-K.; Kim, Y.-S.; Natarajan, S.B.; Kim, W.-S.; Hwang, J.-W.; Jeon, N.-J.; Jeong, J.-H.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Antioxidant and anti-inflammatory effects of Chaenomeles sinensis leaf extracts on LPS-stimulated RAW 264.7 cells. Molecules 2016, 21, 422. [Google Scholar] [CrossRef]
- Noguchi, A.; Tominaga, M.; Takahashi, N.; Matsuda, H.; Kamata, Y.; Umehara, Y.; Ko, K.C.; Suga, Y.; Ogawa, H.; Takamori, K. Differences in therapeutic effects of topically applied corticosteroid and tacrolimus on atopic dermatitis-like symptoms in NC/Nga mice. J. Dermatol. Sci. 2017, 86, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; An, D.; Lee, M.-H.; Lee, K.; Kim, B.; Suman, C.K.; Ham, I.; Choi, H.-Y. Effect of Acer tegmentosum bark on atopic dermatitis-like skin lesions in NC/Nga mice. J. Ethnopharmacol. 2016, 177, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Arellano, F.M.; Wentworth, C.E.; Arana, A.; Fernandez, C.; Paul, C.F. Risk of lymphoma following exposure to calcineurin inhibitors and topical steroids in patients with atopic dermatitis. J. Investig. Dermatol. 2007, 127, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Aioi, A.; Tonogaito, H.; Suto, H.; Hamada, K.; Ra, C.R.; Ogawa, H.; Maibach, H.; Matsuda, H. Impairment of skin barrier function in NC/Nga Tnd mice as a possible model for atopic dermatitis. Br. J. Dermatol. 2001, 144, 12–18. [Google Scholar] [CrossRef]
- Abramson, S.B. Nitric oxide in inflammation and pain associated with osteoarthritis. Arthritis Res. Ther. 2008, 10, S2. [Google Scholar] [CrossRef] [PubMed]
- Nedoszytko, B.; Sokołowska-Wojdyło, M.; Ruckemann-Dziurdzińska, K.; Roszkiewicz, J.; Nowicki, R.J. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: Atopic dermatitis, psoriasis and skin mastocytosis. Adv. Dermatol. Allergol. Postepy. Dermatol. Alergol. 2014, 31, 84–91. [Google Scholar] [CrossRef]
- Choi, Y.A.; Choi, J.K.; Jang, Y.H.; Lee, S.; Lee, S.R.; Choi, J.H.; Park, J.H.; Shin, T.Y.; Kim, S.H. Antiinflammatory effect of Amomum xanthioides in a mouse atopic dermatitis model. Mol. Med. Rep. 2017, 16, 8964–8972. [Google Scholar] [CrossRef]
- Vestergaard, C.; Bang, K.; Gesser, B.; Yoneyama, H.; Matsushima, K.; Larsen, C.G. A Th2 chemokine, TARC, produced by keratinocytes may recruit CLA+CCR4+ lymphocytes into lesional atopic dermatitis skin. J. Investig. Dermatol. 2000, 115, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.K.; Nakamura, N.; Fuseda, K.; Tanaka, H.; Inagaki, N.; Nagai, H. Establishment of allergic dermatitis in NC/Nga mice as a model for severe atopic dermatitis. Biol. Pharm. Bull. 2004, 27, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Chuang, H.Y.; Shih, C.C.; Jong, S.B.; Chang, C.H.; Yu, H.S. Transepidermal water loss, serum IgE and beta-endorphin as important and independent biological markers for development of itch intensity in atopic dermatitis. Br. J. Dermatol. 2006, 154, 1100–1107. [Google Scholar] [CrossRef]
- Kawakami, T.; Ando, T.; Kimura, M.; Wilson, B.S.; Kawakami, Y. Mast cells in atopic dermatitis. Curr. Opin. Immunol. 2009, 21, 666–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Choi, C.H.; Lee, K.; Lee, M.; Ham, I.; Choi, H.Y. Effects of Catalpa ovata stem bark on atopic dermatitis-like skin lesions in NC/Nga mice. J. Ethnopharmacol. 2013, 145, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Lin, Z.C.; Hu, S.C.; Chiang, Y.C.; Hsu, L.F.; Lin, Y.C.; Lee, I.T.; Tsai, M.H.; Fang, J.Y. Urban particulate matter down-regulates filaggrin via COX2 expression/PGE2 production leading to skin barrier dysfunction. Sci. Rep. 2016, 6, 27995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Choi, M.J.; Bak, D.H.; Lee, B.C.; Ko, E.J.; Ahn, G.R.; Ahn, S.W.; Kim, M.J.; Na, J.; Kim, B.J. Topical administration of EGF suppresses immune response and protects skin barrier in DNCB-induced atopic dermatitis in NC/Nga mice. Sci. Rep. 2018, 8, 11895. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content of Astragalin in the Pulc Fraction (Μg/100 Μg of Powdered Pulc Fraction) | |
|---|---|
| Sample | Astragalin |
| PULC | 0.03 ± 0.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, K.; Kang, M.C.; Parveen, A.; Yumnam, S.; Kim, S.Y. Anti-Inflammatory Effect of Chloroform Fraction of Pyrus Ussuriensis Maxim. Leaf Extract on 2, 4-Dinitrochlorobenzene-Induced Atopic Dermatitis in nc/nga Mice. Nutrients 2019, 11, 276. https://doi.org/10.3390/nu11020276
Cho K, Kang MC, Parveen A, Yumnam S, Kim SY. Anti-Inflammatory Effect of Chloroform Fraction of Pyrus Ussuriensis Maxim. Leaf Extract on 2, 4-Dinitrochlorobenzene-Induced Atopic Dermatitis in nc/nga Mice. Nutrients. 2019; 11(2):276. https://doi.org/10.3390/nu11020276
Chicago/Turabian StyleCho, KyoHee, Min Cheol Kang, Amna Parveen, Silvia Yumnam, and Sun Yeou Kim. 2019. "Anti-Inflammatory Effect of Chloroform Fraction of Pyrus Ussuriensis Maxim. Leaf Extract on 2, 4-Dinitrochlorobenzene-Induced Atopic Dermatitis in nc/nga Mice" Nutrients 11, no. 2: 276. https://doi.org/10.3390/nu11020276