The Safety and Tolerability of a Potential Alginate-Based Iron Chelator; Results of A Healthy Participant Study

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Healthy Participant Study Design
2.2. Ethics Statement
2.3. Faecal Microbiota Assessment (Healthy Participant Study and SHIME)
2.4. Healthy Volunteer Faecal Iron and Calcium Assessment
2.5. Faecal Reactive Oxygen Species Assessment
2.6. Intestinal Model (SHIME)
2.7. SHIME Iron Assessments
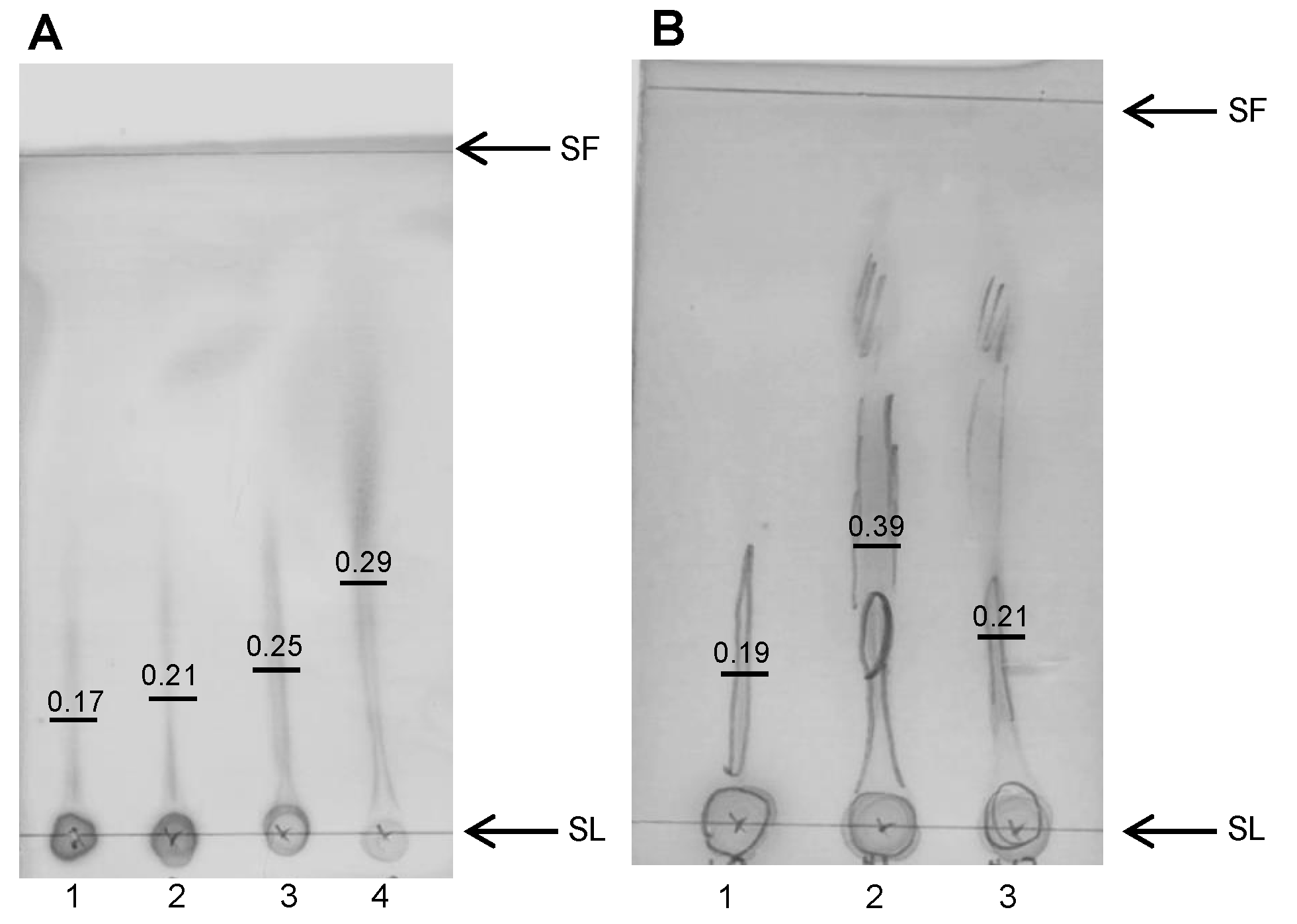
2.8. Thin Layer Chromatography (TLC)
2.9. Statistical Methods
3. Results
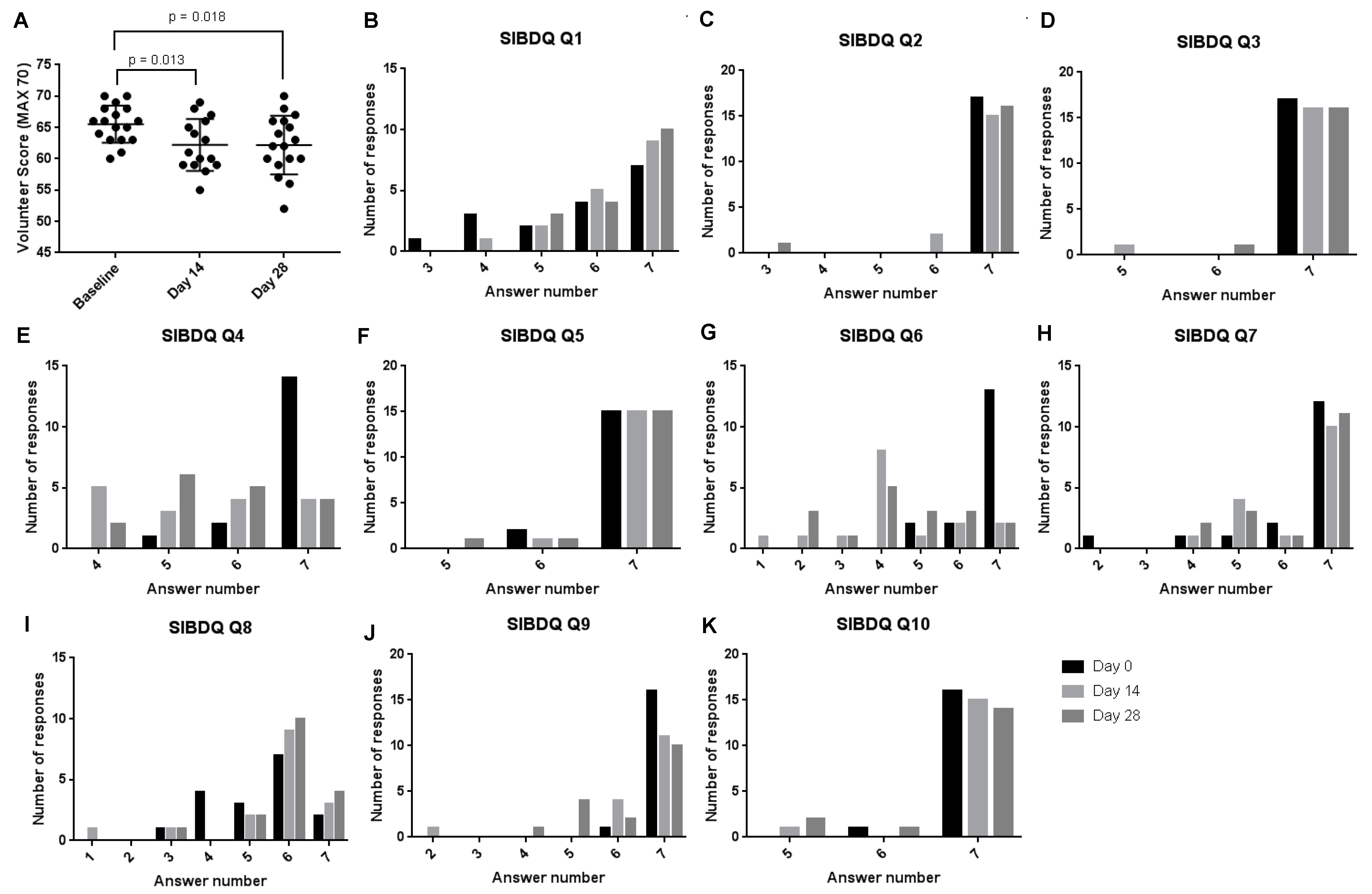
3.1. Study Participant Baseline Demographics
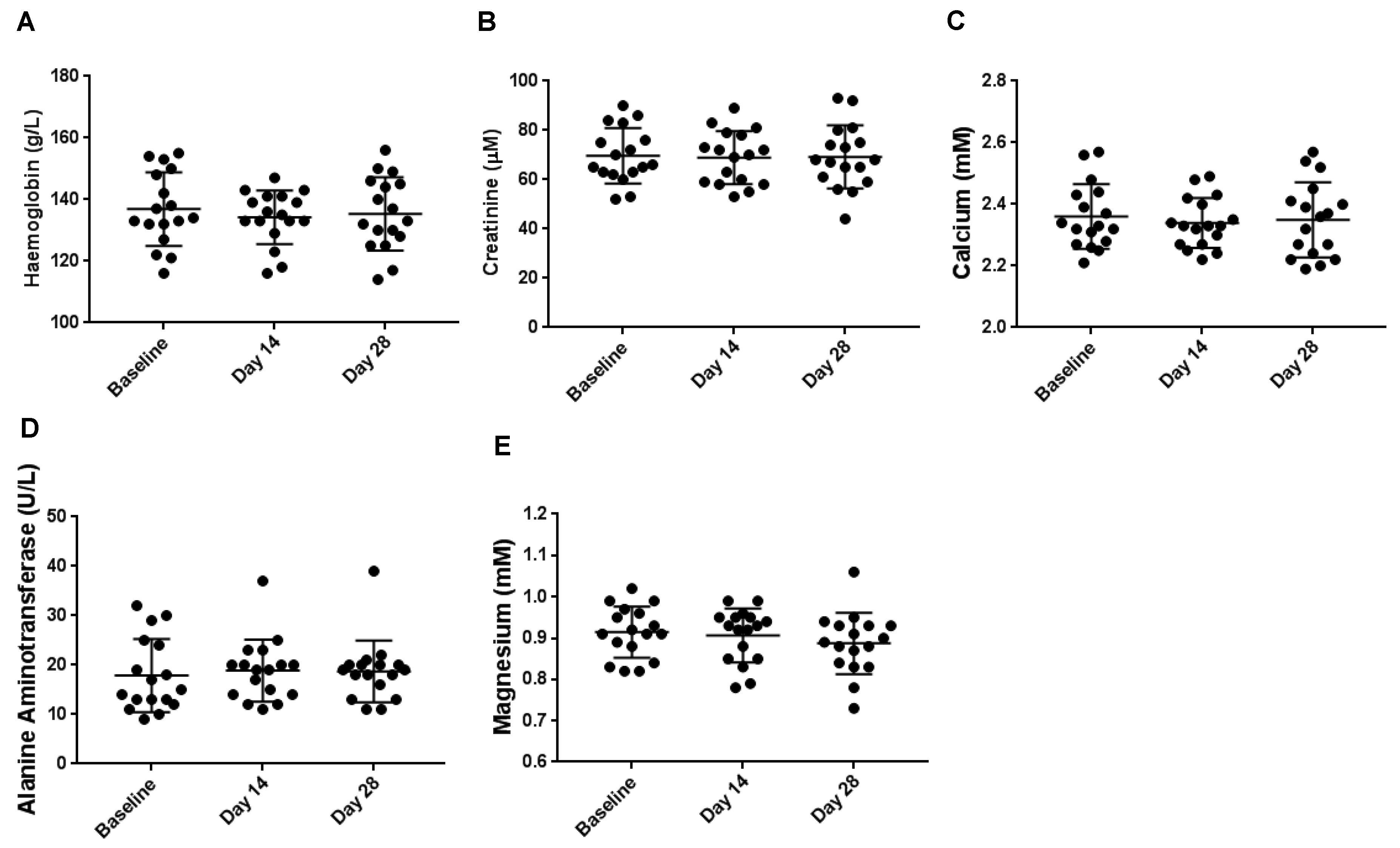
3.2. Haematological Biochemical Analyses
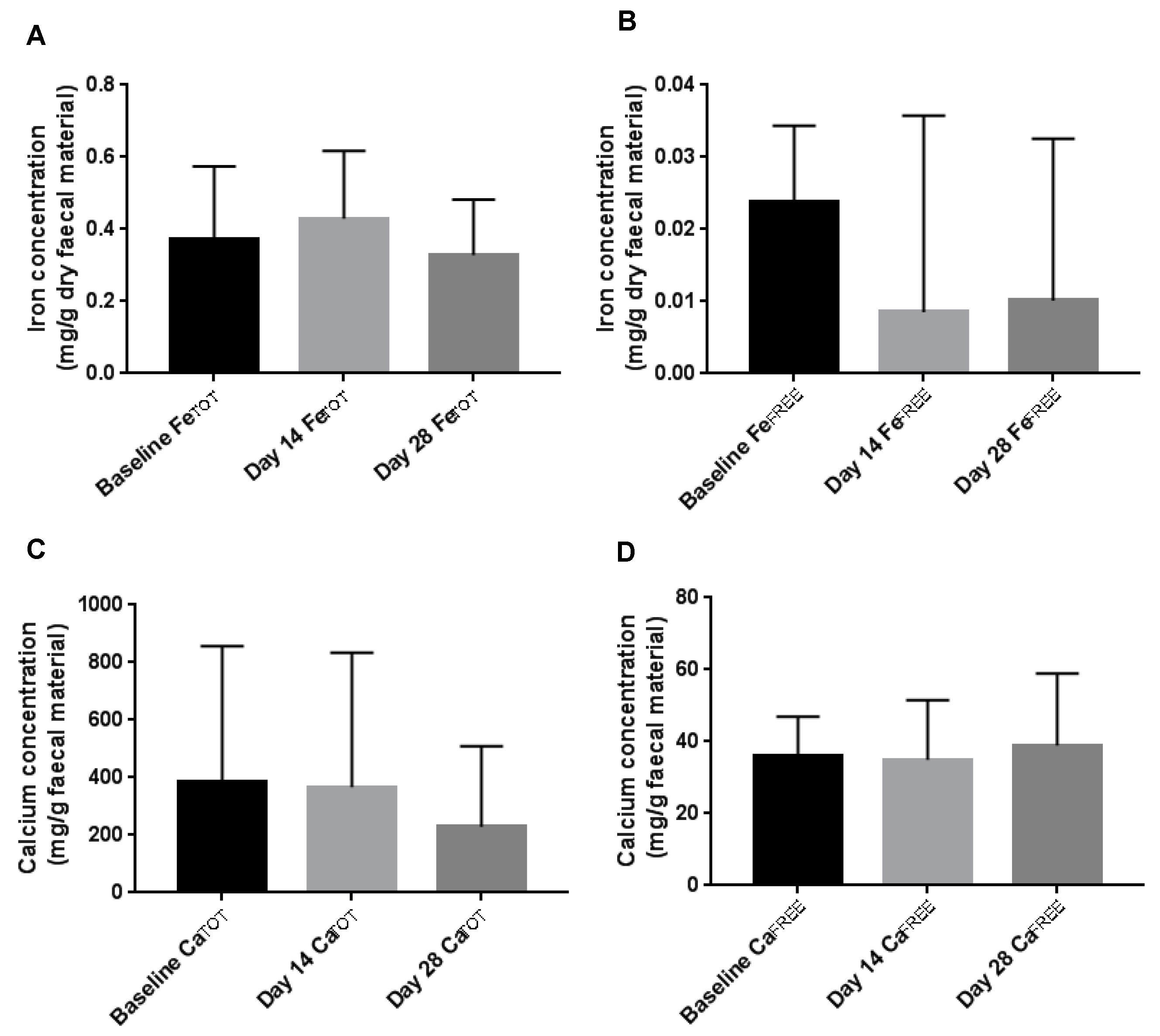
3.3. Faecal Iron and Calcium Concentration
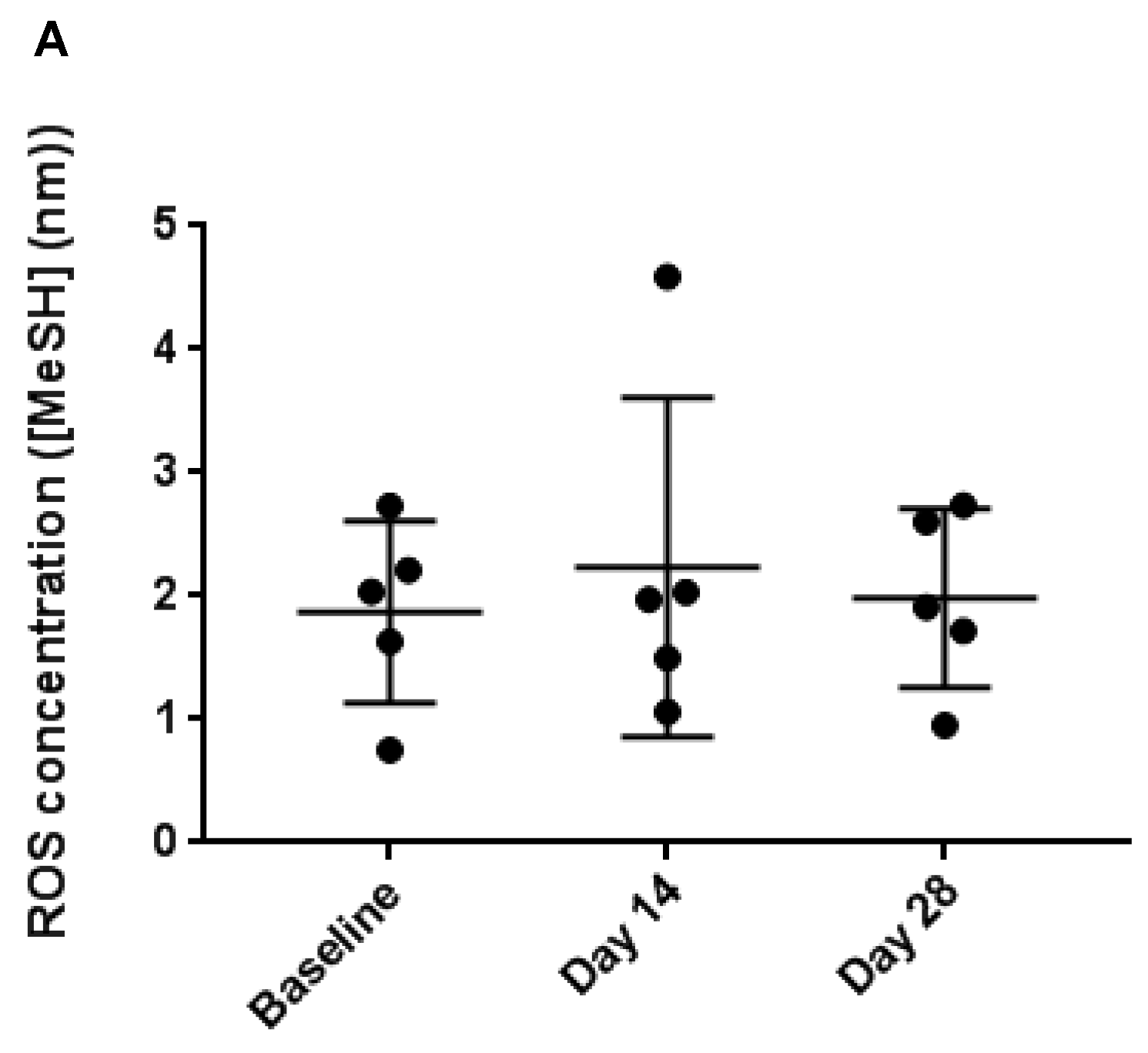
3.4. Faecal Reactive Oxygen Species
3.5. Healthy Participant Microbiome Analysis
3.6. Iron, ROS and Ammonia Concentrations within SHIME
3.7. SHIME SCFA Analysis
3.8. SHIME Microbiome Analysis
3.9. Breakdown of Alginate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kortman, G.A.; Raffatellu, M.; Swinkels, D.W.; Tjalsma, H. Nutritional iron turned inside out: Intestinal stress from a gut microbial perspective. FEMS Microbiol. Rev. 2014, 38, 1202–1234. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.V.; Torti, F.M. Iron and cancer: More ore to be mined. Nat. Rev. Cancer 2013, 13, 342. [Google Scholar] [CrossRef]
- Seril, D.N.; Liao, J.; Yang, G.-Y.; Yang, C.S. Oxidative stress and ulcerative colitis-associated carcinogenesis: Studies in humans and animal models. Carcinogenesis 2003, 24, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Wagner, S.J.; Martínez, I.; Walter, J.; Chang, J.-S.; Clavel, T.; Kisling, S.; Schuemann, K.; Haller, D. Depletion of luminal iron alters the gut microbiota and prevents Crohn’s disease-like ileitis. Gut 2011, 60, 325. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Naito, Y.; Mizuhima, K.; Hirai, Y.; Harusato, A.; Okayama, T.; Katada, K.; Kamada, K.; Uchiyama, K.; Handa, O. Heme oxygenase-1 prevents murine intestinal inflammation. J. Clin. Biochem. Nutr. 2018, 17–133. [Google Scholar] [CrossRef] [PubMed]
- Carrier, J.; Aghdassi, E.; Platt, I.; Cullen, J.; Allard, J. Effect of oral iron supplementation on oxidative stress and colonic inflammation in rats with induced colitis. Aliment. Pharmacol. Ther. 2001, 15, 1989–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glei, M.; Latunde-Dada, G.O.; Klinder, A.; Becker, T.W.; Hermann, U.; Voigt, K.; Pool-Zobel, B.L. Iron-overload induces oxidative DNA damage in the human colon carcinoma cell line HT29 clone 19A. Mt. Res./Genet. Toxicol. Environ. Mutagen. 2002, 519, 151–161. [Google Scholar] [CrossRef]
- Lund, E.K.; Wharf, S.G.; Fairweather-Tait, S.J.; Johnson, I.T. Oral ferrous sulfate supplements increase the free radical–generating capacity of feces from healthy volunteers. Am. J. Clin. Nutr. 1999, 69, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M.B.; Chassard, C.; Rohner, F.; N’Goran, E.K.; Nindjin, C.; Dostal, A.; Utzinger, J.; Ghattas, H.; Lacroix, C.; Hurrell, R.F. The effects of iron fortification on the gut microbiota in African children: A randomized controlled trial in Côte d’Ivoire. Am. J. Clin. Nutr. 2010, 92, 1406–1415. [Google Scholar] [CrossRef]
- Fang, S.; Zhuo, Z.; Yu, X.; Wang, H.; Feng, J. Oral administration of liquid iron preparation containing excess iron induces intestine and liver injury, impairs intestinal barrier function and alters the gut microbiota in rats. J. Trace Elem. Med. Biol. 2018, 47, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Paganini, D.; Zimmermann, M.B. The effects of iron fortification and supplementation on the gut microbiome and diarrhea in infants and children: A review. Am. J. Clin. Nutr. 2017, 106, 1688S–1693S. [Google Scholar] [CrossRef]
- Constante, M.; Fragoso, G.; Calvé, A.; Samba-Mondonga, M.; Santos, M.M. Dietary heme induces gut dysbiosis, aggravates colitis, and potentiates the development of adenomas in mice. Front. Microbiol. 2017, 8, 1809. [Google Scholar] [CrossRef]
- Jaeggi, T.; Kortman, G.A.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Chua, A.C.; Klopcic, B.R.; Ho, D.S.; Fu, S.K.; Forrest, C.H.; Croft, K.D.; Olynyk, J.K.; Lawrance, I.C.; Trinder, D. Dietary iron enhances colonic inflammation and IL-6/IL-11-Stat3 signaling promoting colonic tumor development in mice. PLoS ONE 2013, 8, e78850. [Google Scholar] [CrossRef] [PubMed]
- Radulescu, S.; Brookes, M.J.; Salgueiro, P.; Ridgway, R.A.; McGhee, E.; Anderson, K.; Ford, S.J.; Stones, D.H.; Iqbal, T.H.; Tselepis, C. Luminal iron levels govern intestinal tumorigenesis after Apc loss in vivo. Cell Rep. 2012, 2, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Collings, R.; Harvey, L.J.; Hooper, L.; Hurst, R.; Brown, T.J.; Ansett, J.; King, M.; Fairweather-Tait, S.J. The absorption of iron from whole diets: A systematic review. Am. J. Clin. Nutr. 2013, 98, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.J.; Cook, W.B.; Hutchinson, C.; Tolkien, Z.; Chatfield, M.; Pereira, D.I.; Lomer, M.C. Dietary fortificant iron intake is negatively associated with quality of life in patients with mildly active inflammatory bowel disease. Nutr. Metab. 2013, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Kaitha, S.; Bashir, M.; Ali, T. Iron deficiency anemia in inflammatory bowel disease. World J. Gastrointest. Pathophysiol. 2015, 6, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Debon, S.J.; Tester, R.F. In vitro binding of calcium, iron and zinc by non-starch polysaccharides. Food Chem. 2001, 73, 401–410. [Google Scholar] [CrossRef]
- Berner, L.A.; Hood, L.F. Iron binding by sodium alginate. J. Food Sci. 1983, 48, 755–758. [Google Scholar] [CrossRef]
- Wölbling, R.; Becker, G.; Forth, W. Inhibition of the intestinal absorption of iron by sodium alginate and guar gum in rats. Digestion 1980, 20, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed]
- Hampson, F.; Farndale, A.; Strugala, V.; Sykes, J.; Jolliffe, I.; Dettmar, P. Alginate rafts and their characterisation. Int. J. Pharm. 2005, 294, 137–147. [Google Scholar] [CrossRef]
- Horniblow, R.D.; Latunde-Dada, G.O.; Harding, S.E.; Schneider, M.; Almutairi, F.M.; Sahni, M.; Bhatti, A.; Ludwig, C.; Norton, I.T.; Iqbal, T.H. The chelation of colonic luminal iron by a unique sodium alginate for the improvement of gastrointestinal health. Mol. Nutr. Food Res. 2016, 60, 2098–2108. [Google Scholar] [CrossRef] [Green Version]
- Wawer, A.A.; Harvey, L.J.; Dainty, J.R.; Perez-Moral, N.; Sharp, P.; Fairweather-Tait, S.J. Alginate inhibits iron absorption from ferrous gluconate in a randomized controlled trial and reduces iron uptake into caco-2 cells. PLoS ONE 2014, 9, e112144. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Roos, S.; Eeckhaut, V.; MacKenzie, D.A.; Derde, M.; Verstraete, W.; Marzorati, M.; Possemiers, S.; Vanhoecke, B.; Van Immerseel, F. Incorporating a mucosal environment in a dynamic gut model results in a more representative colonization by lactobacilli. Microb. Biotechnol. 2012, 5, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, C.; Marzorati, M.; Innocenti, M.; Vilchez-Vargas, R.; Vital, M.; Pieper, D.H.; Van de Wiele, T.; Mulinacci, N. Dietary supplement based on stilbenes: A focus on gut microbial metabolism by the in vitro simulator M-SHIME®. Food Funct. 2016, 7, 4564–4575. [Google Scholar] [CrossRef]
- Li, M.; Li, G.; Shang, Q.; Chen, X.; Liu, W.; Pi, X.; Zhu, L.; Yin, Y.; Yu, G.; Wang, X. In vitro fermentation of alginate and its derivatives by human gut microbiota. Anaerobe 2016, 39, 19–25. [Google Scholar] [CrossRef]
- Lee, T.; Clavel, T.; Smirnov, K.; Schmidt, A.; Lagkouvardos, I.; Walker, A.; Lucio, M.; Michalke, B.; Schmitt-Kopplin, P.; Fedorak, R. Oral versus intravenous iron replacement therapy distinctly alters the gut microbiota and metabolome in patients with IBD. Gut 2017, 66, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Mahalhal, A.; Williams, J.M.; Johnson, S.; Ellaby, N.; Duckworth, C.A.; Burkitt, M.D.; Liu, X.; Hold, G.L.; Campbell, B.J.; Pritchard, D.M. Oral iron exacerbates colitis and influences the intestinal microbiome. PLoS ONE 2018, 13, e0202460. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Frank, D.N.; Hendricks, A.E.; Ir, D.; Esamai, F.; Liechty, E.; Hambidge, K.M.; Krebs, N.F. Iron in micronutrient powder promotes an unfavorable gut microbiota in Kenyan infants. Nutrients 2017, 9, 776. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharm. Rev. 2010, 4, 118. [Google Scholar] [CrossRef] [PubMed]
- Additives, E.P.O.F.; Food, N.S.A.t.; Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; et al. Re-evaluation of alginic acid and its sodium, potassium, ammonium and calcium salts (E 400–E 404) as food additives. EFSA J. 2017, 15, e05049. [Google Scholar]
- Cummings, J.H.; Macfarlane, G.T. Gastrointestinal effects of prebiotics. Br. J. Nutr. 2002, 87, S145–S151. [Google Scholar] [CrossRef] [PubMed]
- Coudray, C.; Bellanger, J.; Castiglia-Delavaud, C.; Remesy, C.; Vermorel, M.; Rayssignuier, Y. Effect of soluble or partly soluble dietary fibres supplementation on absorption and balance of calcium, magnesium, iron and zinc in healthy young men. Eur. J. Clin. Nutr. 1997, 51, 375. [Google Scholar] [CrossRef]
- PLAlT, S.R.; Clydesdale, F.M. Mineral binding characteristics of lignin, guar gum, cellulose, pectin and neutral detergent fiber under simulated duodenal pH conditions. J. Food Sci. 1987, 52, 1414–1419. [Google Scholar] [CrossRef]
- Powell, J.; Jugdaohsingh, R.; Thompson, R. The regulation of mineral absorption in the gastrointestinal tract. Proc. Nutr. Soc. 1999, 58, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.I.; Bruggraber, S.F.; Faria, N.; Poots, L.K.; Tagmount, M.A.; Aslam, M.F.; Frazer, D.M.; Vulpe, C.D.; Anderson, G.J.; Powell, J.J. Nanoparticulate iron (III) oxo-hydroxide delivers safe iron that is well absorbed and utilised in humans. Nanomed. Nanotechnol. Biol. Med. 2014, 10, 1877–1886. [Google Scholar] [CrossRef]
- Vanlancker, E.; Vanhoecke, B.; Stringer, A.; Van de Wiele, T. 5-Fluorouracil and irinotecan (SN-38) have limited impact on colon microbial functionality and composition in vitro. PeerJ 2017, 5, e4017. [Google Scholar] [CrossRef]
- Wang, Y.; Han, F.; Hu, B.; Li, J.; Yu, W. In vivo prebiotic properties of alginate oligosaccharides prepared through enzymatic hydrolysis of alginate. Nutr. Res. 2006, 26, 597–603. [Google Scholar] [CrossRef]
- Chen, X.; Sun, Y.; Hu, L.; Liu, S.; Yu, H.; Li, R.; Wang, X.; Li, P. In vitro prebiotic effects of seaweed polysaccharides. J. Oceanol. Limnol. 2018, 36, 926–932. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adverse Effect | Number of Participants Reported |
|---|---|
| Flatulence | 16 |
| Bloating | 7 |
| Heartburn | 2 |
| Abdominal Pain | 8 |
| Loose Motion | 4 |
| Urgency to Defecate | 1 |
| Nausea and Vomiting | 3 |
| Headache | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horniblow, R.D.; Mistry, P.; Quraishi, M.N.; Beggs, A.D.; Van de Wiele, T.; Iqbal, T.H.; Tselepis, C. The Safety and Tolerability of a Potential Alginate-Based Iron Chelator; Results of A Healthy Participant Study. Nutrients 2019, 11, 674. https://doi.org/10.3390/nu11030674
Horniblow RD, Mistry P, Quraishi MN, Beggs AD, Van de Wiele T, Iqbal TH, Tselepis C. The Safety and Tolerability of a Potential Alginate-Based Iron Chelator; Results of A Healthy Participant Study. Nutrients. 2019; 11(3):674. https://doi.org/10.3390/nu11030674
Chicago/Turabian StyleHorniblow, Richard D., Pritesh Mistry, Mohammed N. Quraishi, Andrew D. Beggs, Tom Van de Wiele, Tariq H. Iqbal, and Chris Tselepis. 2019. "The Safety and Tolerability of a Potential Alginate-Based Iron Chelator; Results of A Healthy Participant Study" Nutrients 11, no. 3: 674. https://doi.org/10.3390/nu11030674
APA StyleHorniblow, R. D., Mistry, P., Quraishi, M. N., Beggs, A. D., Van de Wiele, T., Iqbal, T. H., & Tselepis, C. (2019). The Safety and Tolerability of a Potential Alginate-Based Iron Chelator; Results of A Healthy Participant Study. Nutrients, 11(3), 674. https://doi.org/10.3390/nu11030674