Central Role of Cell Cycle Regulation in the Antitumoral Action of Ocoxin


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. OOS Composition
2.2. Cells Culture and Animal Experiments
2.3. Functional Genomic and Transcriptomic Profiling
2.4. Evaluation of Transcription Factor Association with Deregulated Genes
2.5. E2F Pathway Map Development and Activation Score
2.6. TCGA Patients Gene Expression Comparison and Outcome Analyses
2.7. Statistical Analyses
3. Results
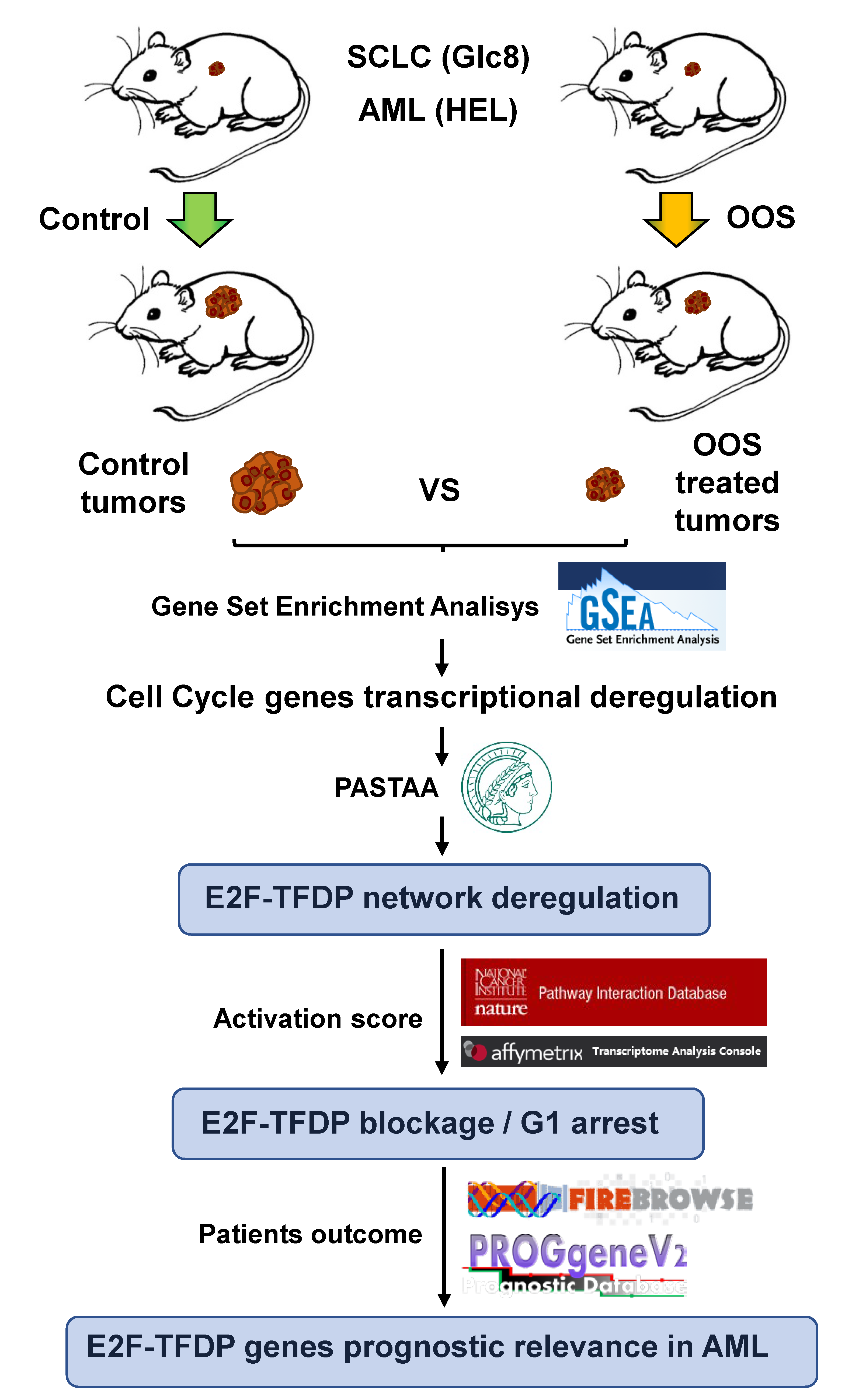
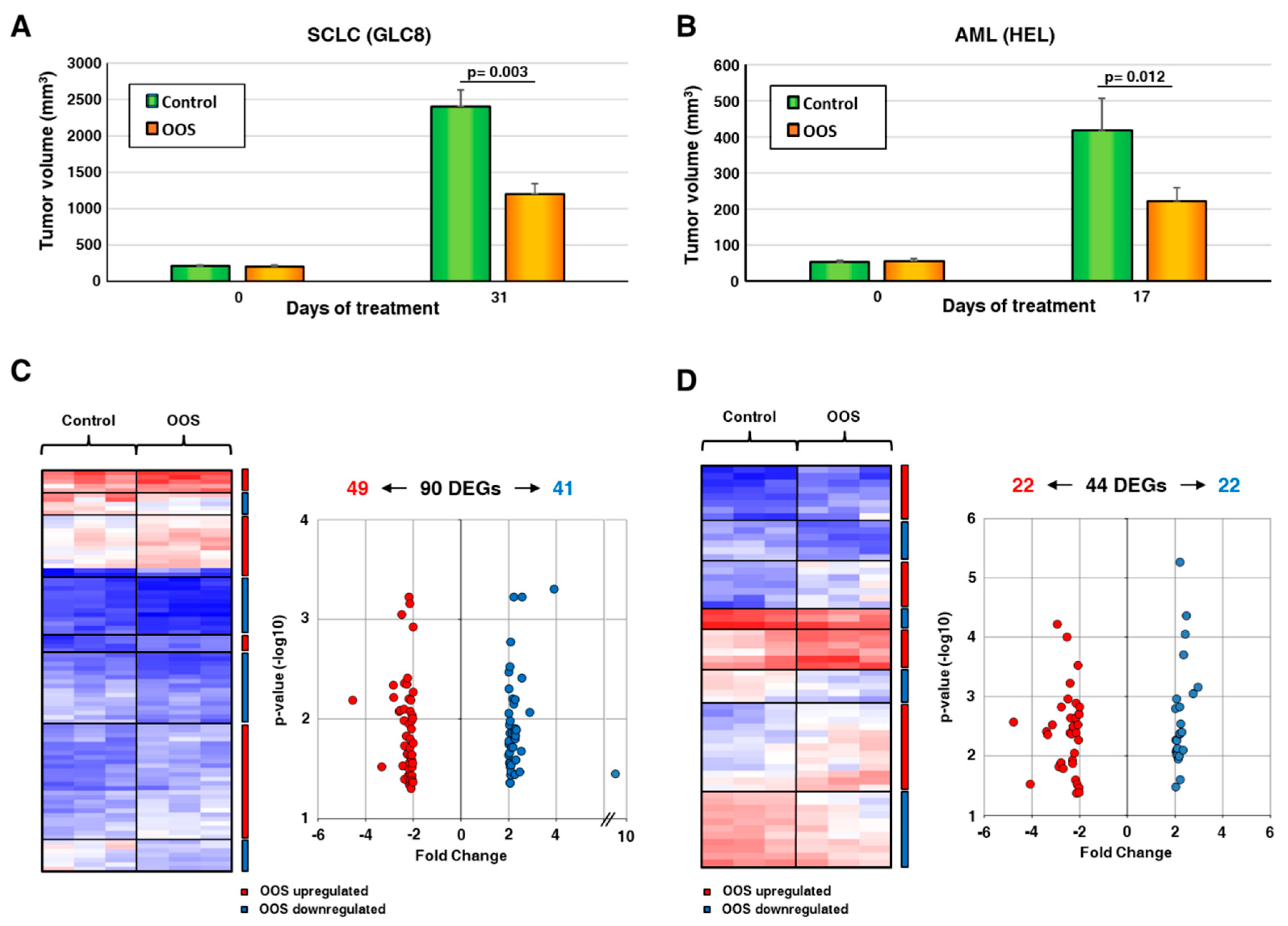
3.1. Genes Deregulated by OOS In Vivo
3.2. Functional Transcriptomic Analyses in AML and SCLC Tumors
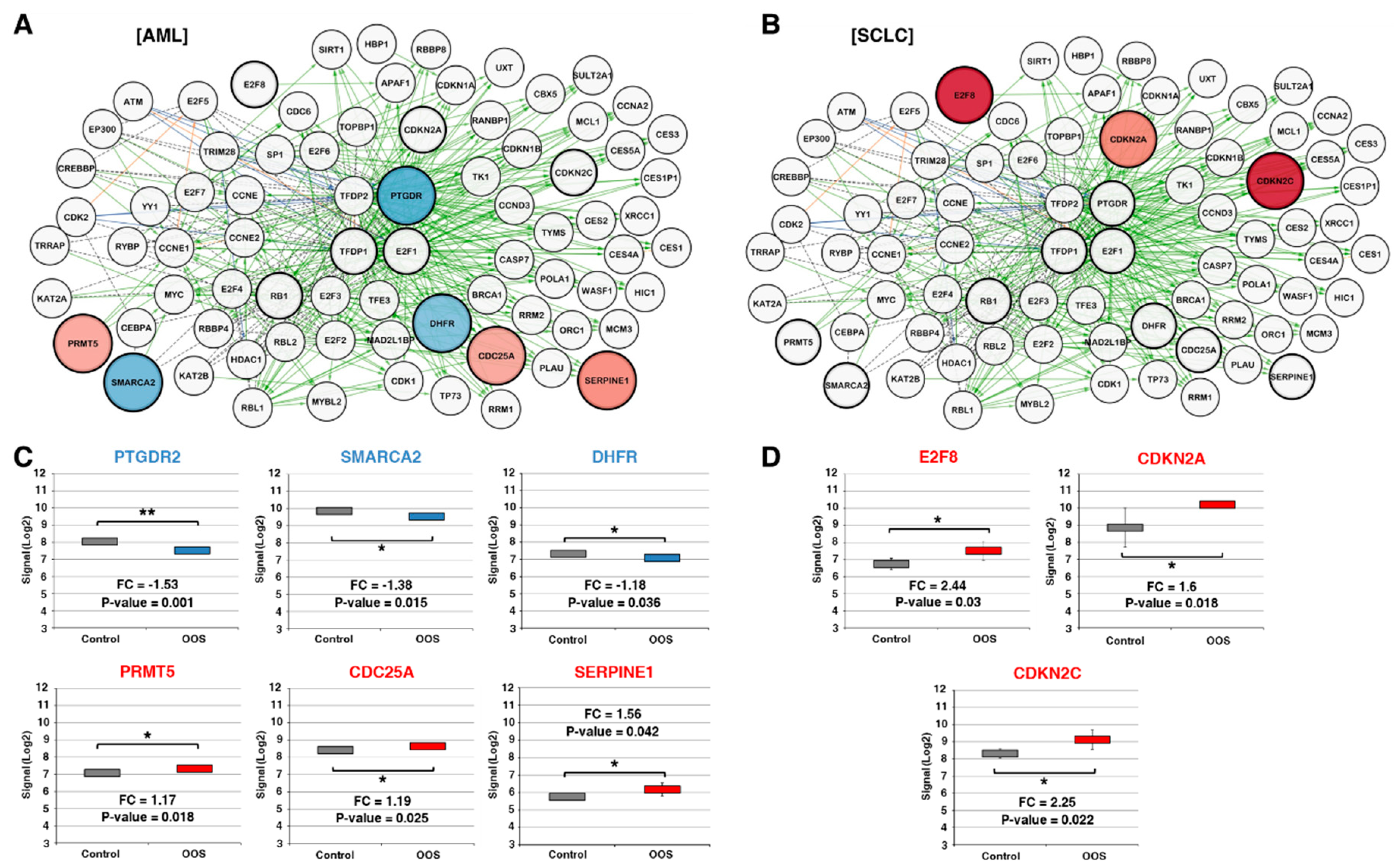
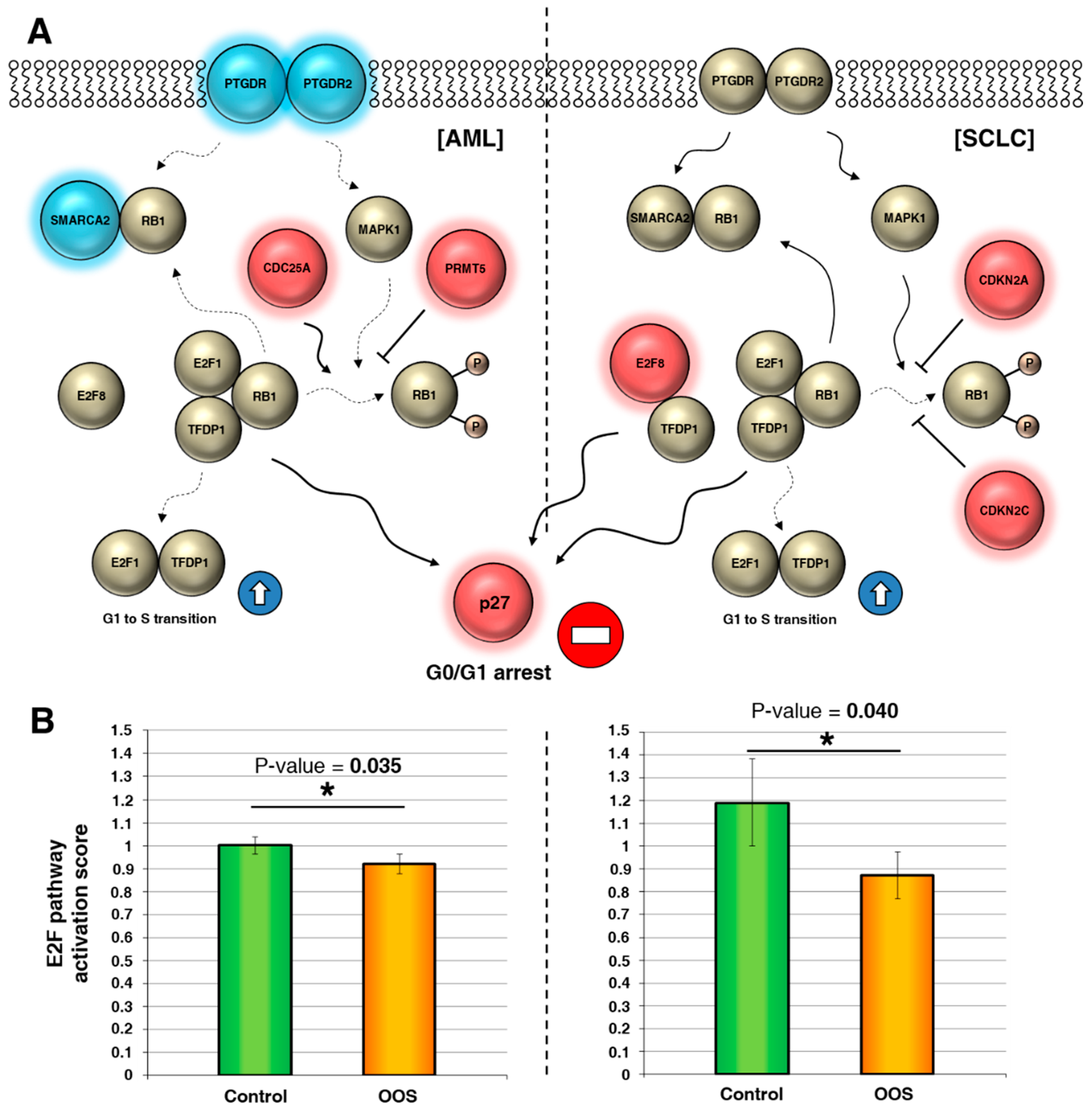
3.3. Action of OOS on the E2F–TFDP Transcriptional Network
3.4. Prognostic Relevance of the E2F–TFDP Route
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fridlender, M.; Kapulnik, Y.; Koltai, H. Plant derived substances with anti-cancer activity: From folklore to practice. Front Plant Sci. 2015, 6, 799. [Google Scholar] [CrossRef] [PubMed]
- Greenwell, M.; Rahman, P.K. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [Green Version]
- Moudi, M.; Go, R.; Yien, C.Y.; Nazre, M. Vinca alkaloids. Int. J. Prev. Med. 2013, 4, 1231–1235. [Google Scholar]
- Mukhtar, E.; Adhami, V.M.; Mukhtar, H. Targeting microtubules by natural agents for cancer therapy. Mol. Cancer Ther. 2014, 13, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Slichenmyer, W.J.; Von Hoff, D.D. New natural products in cancer chemotherapy. J. Clin. Pharmacol. 1990, 30, 770–788. [Google Scholar] [CrossRef] [PubMed]
- Slichenmyer, W.J.; Von Hoff, D.D. Taxol: A new and effective anti-cancer drug. Anticancer Drugs 1991, 2, 519–530. [Google Scholar] [CrossRef]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef]
- Al-Mahtab, M.; Akbar, S.; Khan, M.; Rahman, S. Increased survival of patients with end-stage hepatocellular carcinoma due to intake of ONCOXIN((R)), a dietary supplement. Indian J. Cancer 2015, 52, 443–446. [Google Scholar]
- Dayem Uddin, M.A.I.; Mahmood, I.; Ghosha, A.K.; Khatuns, R.A. Findings of the 3-Month supportive treatment with ocoxin solution beside the standard modalities of patients with different neoplastic diseases. TAJ 2009, 22, 172–175. [Google Scholar]
- Hollman, P.C.; Feskens, E.J.; Katan, M.B. Tea flavonols in cardiovascular disease and cancer epidemiology. Proc. Soc. Exp. Biol. Med. 1999, 220, 198–202. [Google Scholar]
- Liu, L.; Hudgins, W.R.; Shack, S.; Yin, M.Q.; Samid, D. Cinnamic acid: A natural product with potential use in cancer intervention. Int. J. Cancer 1995, 62, 345–350. [Google Scholar] [CrossRef]
- Yang, C.S.; Chung, J.Y.; Yang, G.; Chhabra, S.K.; Lee, M.J. Tea and tea polyphenols in cancer prevention. J. Nutr. 2000, 130, 472S–478S. [Google Scholar] [CrossRef]
- Sankaranarayanan, R.; Valiveti, C.K.; Kumar, D.R.; Van Slambrouck, S.; Kesharwani, S.S.; Seefeldt, T.; Scaria, J.; Tummala, H.; Bhat, G.J. The Flavonoid Metabolite 2,4,6-Trihydroxybenzoic Acid Is a CDK Inhibitor and an Anti-Proliferative Agent: A Potential Role in Cancer Prevention. Cancers 2019, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.V.; Perez, Y.M.; Sanchez, H.V.; Forment, G.R.; Soler, E.A.; Bertot, L.C.; Garcia, A.Y.; del Rosario Abreu Vazquez, M.; Fabian, L.G. Antioxidant and immunomodulatory effects of Viusid in patients with chronic hepatitis C. World J. Gastroenterol. 2010, 16, 2638–2647. [Google Scholar] [CrossRef]
- Hernandez-Garcia, S.; Gonzalez, V.; Sanz, E.; Pandiella, A. Effect of Oncoxin Oral Solution in HER2-Overexpressing Breast Cancer. Nutr. Cancer 2015, 67, 1159–1169. [Google Scholar] [CrossRef]
- Hernandez-Unzueta, I.; Benedicto, A.; Olaso, E.; Sanz, E.; Viera, C.; Arteta, B.; Marquez, J. Ocoxin oral solution((R)) as a complement to irinotecan chemotherapy in the metastatic progression of colorectal cancer to the liver. Oncol. Lett. 2017, 13, 4002–4012. [Google Scholar] [CrossRef]
- Marquez, J.; Mena, J.; Hernandez-Unzueta, I.; Benedicto, A.; Sanz, E.; Arteta, B.; Olaso, E. Ocoxin(R) oral solution slows down tumor growth in an experimental model of colorectal cancer metastasis to the liver in Balb/c mice. Oncol. Rep. 2016, 35, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Rodriguez, E.; El-Mallah, A.M.; Sanz, E.; Pandiella, A. Antitumoral effect of Ocoxin in hepatocellular carcinoma. Oncol. Lett. 2017, 14, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Rodriguez, E.; Sanz, E.; Pandiella, A. Antitumoral effect of Ocoxin, a natural compound-containing nutritional supplement, in small cell lung cancer. Int. J. Oncol. 2018, 53, 113–123. [Google Scholar] [PubMed] [Green Version]
- Diaz-Rodriguez, E.; Hernandez-Garcia, S.; Sanz, E.; Pandiella, A. Antitumoral effect of Ocoxin on acute myeloid leukemia. Oncotarget 2016, 7, 6231–6242. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Rodriguez, E.; Perez-Pena, J.; Rios-Luci, C.; Arribas, J.; Ocana, A.; Pandiella, A. TRAIL receptor activation overcomes resistance to trastuzumab in HER2 positive breast cancer cells. Cancer Lett. 2019. [Google Scholar] [CrossRef] [PubMed]
- Roider, H.G.; Kanhere, A.; Manke, T.; Vingron, M. Predicting transcription factor affinities to DNA from a biophysical model. Bioinformatics 2007, 23, 134–141. [Google Scholar] [CrossRef]
- Schaefer, C.F.; Anthony, K.; Krupa, S.; Buchoff, J.; Day, M.; Hannay, T.; Buetow, K.H. PID: The Pathway Interaction Database. Nucleic Acids Res. 2009, 37, D674–D679. [Google Scholar] [CrossRef]
- Dyson, N. The regulation of E2F by pRB-family proteins. Genes Dev. 1998, 12, 2245–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harbour, J.W.; Dean, D.C. The Rb/E2F pathway: Expanding roles and emerging paradigms. Genes Dev. 2000, 14, 2393–2409. [Google Scholar] [CrossRef]
- Trimarchi, J.M.; Lees, J.A. Sibling rivalry in the E2F family. Nat. Rev. Mol. Cell Biol. 2002, 3, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H. Green tea: Health benefits as cancer preventive for humans. Chem. Rec. 2005, 5, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Hostetler, B.J.; Uchakina, O.N.; Ban, H.; McKallip, R.J. Treatment of Hematological Malignancies with Glycyrrhizic Acid. Anticancer Res. 2017, 37, 997–1004. [Google Scholar] [Green Version]
- Kuttan, G.; Pratheeshkumar, P.; Manu, K.A.; Kuttan, R. Inhibition of tumor progression by naturally occurring terpenoids. Pharm. Biol. 2011, 49, 995–1007. [Google Scholar] [CrossRef] [Green Version]
- Koh, D.W.; Dawson, T.M.; Dawson, V.L. Mediation of cell death by poly(ADP-ribose) polymerase-1. Pharmacol. Res. 2005, 52, 5–14. [Google Scholar] [CrossRef]
- Konopleva, M.; Zhao, S.; Xie, Z.; Segall, H.; Younes, A.; Claxton, D.F.; Estrov, Z.; Kornblau, S.M.; Andreeff, M. Apoptosis. Molecules and mechanisms. Adv. Exp. Med. Biol. 1999, 457, 217–236. [Google Scholar]
- Soldani, C.; Scovassi, A.I. Poly(ADP-ribose) polymerase-1 cleavage during apoptosis: An update. Apoptosis 2002, 7, 321–328. [Google Scholar] [CrossRef]
- Duriez, P.J.; Shah, G.M. Cleavage of poly(ADP-ribose) polymerase: A sensitive parameter to study cell death. Biochem. Cell Biol. 1997, 75, 337–349. [Google Scholar] [CrossRef]
- Thurlings, I.; de Bruin, A. E2F Transcription Factors Control the Roller Coaster Ride of Cell Cycle Gene Expression. Methods Mol. Biol. 2016, 1342, 71–88. [Google Scholar]
- Chen, H.Z.; Tsai, S.Y.; Leone, G. Emerging roles of E2Fs in cancer: An exit from cell cycle control. Nat. Rev. Cancer 2009, 9, 785–797. [Google Scholar] [CrossRef]
- Classon, M.; Harlow, E. The retinoblastoma tumour suppressor in development and cancer. Nat. Rev. Cancer 2002, 2, 910–917. [Google Scholar] [CrossRef]
- Knudsen, E.S.; Knudsen, K.E. Tailoring to RB: Tumour suppressor status and therapeutic response. Nat. Rev. Cancer 2008, 8, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Henry, D.; Brumaire, S.; Hu, X. Involvement of pRb-E2F pathway in green tea extract-induced growth inhibition of human myeloid leukemia cells. Leuk. Res. 2019, 77, 34–41. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, M.; Chen, N.; Ma, A.; Zhu, C.; Zhao, R.; Jiang, M.; Zhou, J.; Ye, L.; Fu, H.; et al. Glycyrrhetinic acid induces G1phase cell cycle arrest in human nonsmall cell lung cancer cells through endoplasmic reticulum stress pathway. Int. J. Oncol. 2015, 46, 981–988. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Peña, J.; Díaz-Rodríguez, E.; Sanz, E.; Pandiella, A. Central Role of Cell Cycle Regulation in the Antitumoral Action of Ocoxin. Nutrients 2019, 11, 1068. https://doi.org/10.3390/nu11051068
Pérez-Peña J, Díaz-Rodríguez E, Sanz E, Pandiella A. Central Role of Cell Cycle Regulation in the Antitumoral Action of Ocoxin. Nutrients. 2019; 11(5):1068. https://doi.org/10.3390/nu11051068
Chicago/Turabian StylePérez-Peña, Javier, Elena Díaz-Rodríguez, Eduardo Sanz, and Atanasio Pandiella. 2019. "Central Role of Cell Cycle Regulation in the Antitumoral Action of Ocoxin" Nutrients 11, no. 5: 1068. https://doi.org/10.3390/nu11051068
APA StylePérez-Peña, J., Díaz-Rodríguez, E., Sanz, E., & Pandiella, A. (2019). Central Role of Cell Cycle Regulation in the Antitumoral Action of Ocoxin. Nutrients, 11(5), 1068. https://doi.org/10.3390/nu11051068