1. Introduction
Skeletal muscle is a dynamic tissue that adapts to external conditions and reacts to acute and long-term physical activity and feeding conditions [
1]. Maintenance of muscle mass is not only important for athletes, but also for health and aging [
2]. In particular, resistance training using a repeated high-weight stimulus optimizes the preservation of muscle mass that is correlated with body strength [
3]. The underlying mechanism of adaptation is an increase in muscle protein synthesis (MPS) stimulated through mechanical load, which can be further augmented by additive protein supplementation [
4]. Amino acids are required for protein synthesis. Leucine, a branched-chain amino acid (BCAA), has been shown to be a prerequisite stimulator of the mammalian target of rapamycin (mTOR) signaling pathway, which plays a critical role in MPS [
5,
6].
Besides the contractile muscle components, passive tissues like bone, cartilage, ligaments, and tendons also adapt to resistance training [
7]. In this context, Mackey et al. [
8] showed that the synthesis of collagen in fibroblasts behaves similarly to MPS after a single bout of resistance training. Collagen is the most represented protein in the human body (30% of the protein concentration). It is an important element of the extracellular matrix (ECM) of skeletal muscle and tendons and is mainly responsible for their functionality in terms of force transmission, flexibility, and adaptation [
9]. Thus, to ensure muscle fiber strength transmission, the ECM structures need to adapt in response to resistance training.
The benefit of collagen supplementation on collagen synthesis regarding tendon architecture has been demonstrated by various studies through the analysis of injured participants [
10,
11]. Studies have reported less joint pain perception after collagen peptide supplementation in healthy active participants and in patients with osteoarthritis [
12,
13,
14,
15,
16,
17], which could be of interest for those involved in elite sport. Clifford et al. [
18] reported a faster recovery from jumping exercise and a tendency to reduce muscle soreness after collagen supplementation.
The beneficial effects of collagen supplementation on passive structures are associated with higher muscle strength following an improvement in force transmission. A study by Zdzieblik et al. [
19] was, to the best of our knowledge, the first study to find a positive effect of collagen supplementation on strength and body composition. In a double-blind controlled trial, 53 elderly men (age: 72.2 ± 4.7 years) completed a 12-week resistance training intervention in combination with supplementation with 15 g of collagen or a placebo, resulting in significantly higher gains in fat-free mass (FFM) and muscle strength in the collagen peptide group. However, the underlying mechanism by which collagen peptides act on muscle protein metabolism remains unclear. Collagen is rich in non-essential amino acids, like proline and glycine, and relatively poor in essential amino acids like leucine, the main trigger for MPS.
Proteomic analysis methods have been increasingly developed and used to provide insight into molecular pathways and analyze responses to physical activity or nutrients [
20,
21,
22,
23]. The advantage of proteomic analysis based on mass spectrometry is the large number of proteins identified, which provides information about all processes with no need to select specific pathways.
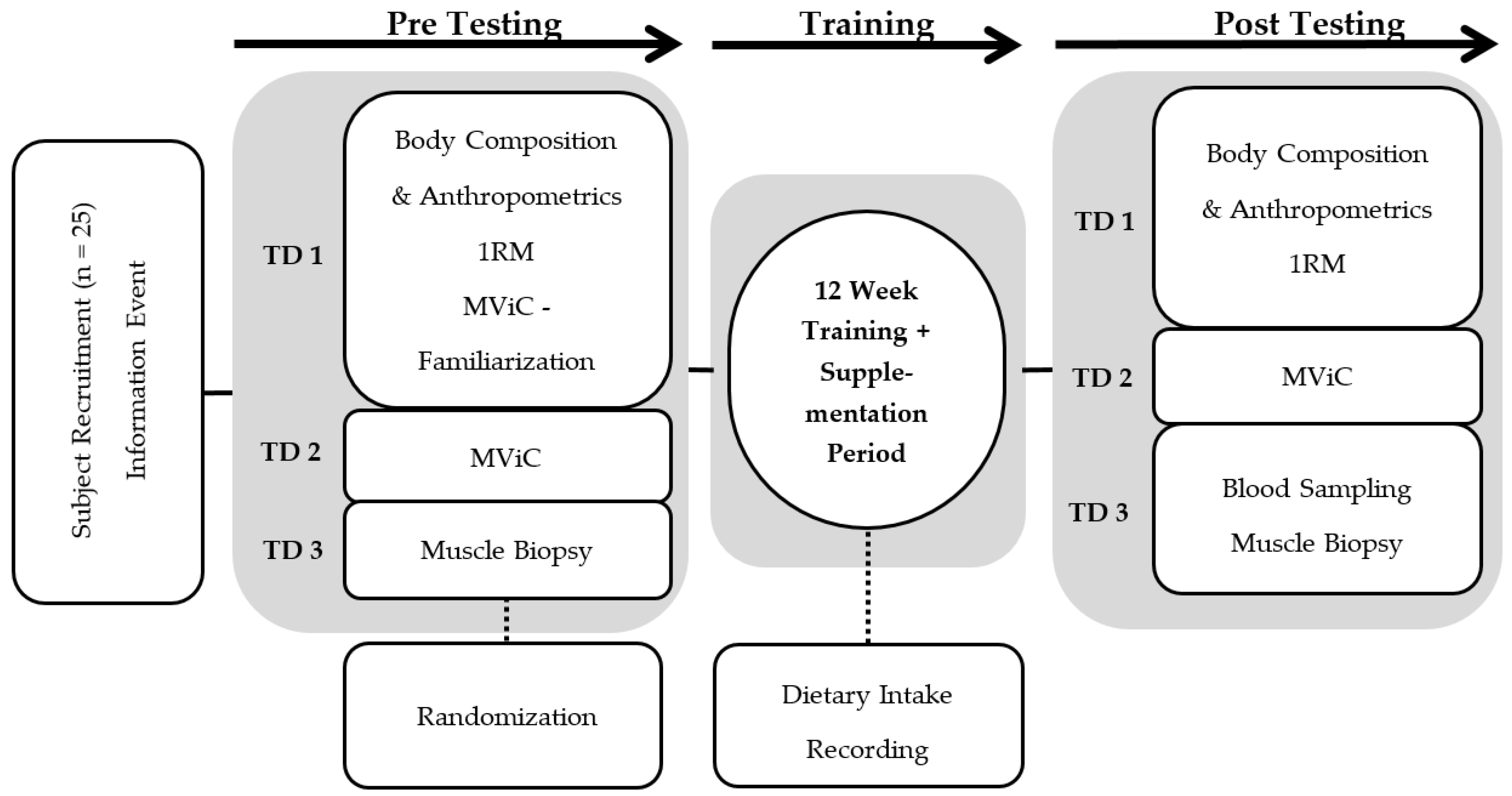
To obtain novel information about long-term adaptations and changes in protein content in response to collagen hydrolysate supplementation in combination with hypertrophy resistance training, we designed a double-blind, randomized, placebo-controlled trial and analyzed muscle proteome before and after a 12-week intervention.
3. Results and Discussion
The analysis of the blood samples revealed significant increases in hydroxyproline for each participant in COL two hours after ingesting the collagen peptide (pre: 33.3 ± 19.7 µmol/L, post: 95.8 ± 27.1 µmol/L;
p ≤ 0.0125) compared to participants in the PLA group (pre: 14.4 ± 6.4 µmol/L, post: 14.3 ± 11.3 µmol/L;
p = 0.956), indicating that all subjects were able to absorb the hydrolyzed collagen [
10].
The dietary assessment showed no differences in caloric intake or macronutrients between the groups. The average protein intake over three days including the supplement was 135.1 ± 29.0 g/day in COL and 145.8 ± 52.7 g/day in PLA (p = 0.557), representing an adequate and equal protein intake of 1.66 g/kg/day (COL) and 1.86 g/kg/day (PLA).
At the beginning of the 12-week intervention period, no statistical differences existed between COL and PLA for any anthropometric or strength parameter (
Table 1). From pre to post (main effect
p ≤ 0.05) the whole cohort increased significantly in all parameters. ANOVA with repeated measurements showed significant interaction effects (
p ≤ 0.05) for BM, FFM, and R, and trends for an interaction effect for SQ (
p = 0.073), BP (
p = 0.099), and MViC (
p = 0.066), which increased slightly more in COL than in PLA, respectively. FM and DL showed no interaction effect between groups. Post-hoc
t-tests with the Bonferroni correction (to
p ≤ 0.0125) revealed no significant differences between groups in BM, FFM, and R after 12 weeks. FFM only increased in COL, whereas no difference was found in PLA.
Considering the differences between groups, delta values from pre to post are given in
Table 2. Individuals in COL gained significantly more BM and FFM in contrast with those in PLA. Individuals in COL had a significantly higher strength increase in R compared to those in PLA (
Table 2).
The study by Zdzieblik et al. [
19] showed a clear improvement in isokinetic quadriceps strength, a greater loss in FM and higher gains in FFM in 26 sarcopenic older men supplementing 15 g collagen peptides per day during a 12-week training intervention compared with a placebo group (
n = 27). In our study, FFM also increased to a higher extend in COL compared with PLA, while strength only slightly increased further in COL compared with PLA, suggesting that mechanisms are more pronounced in the elderly populations than in younger healthy men. Because collagen has a high impact on tendon tissues [
10,
11], it is possible that exercises that stimulate reactive power of the tendons would have shown clearer effects of collagen peptide supplementation [
18]. The strength tests used in this study only slightly depend on reactive forces and might therefore have underestimated possible effects of the supplement on tendon tissues. Another part of our study analyzed the muscle fiber cross-sectional area and showed no differences between groups [
24], underpinning the hypothesis that adaptations induced by collagen supplements do not affect the muscle cell per se but structural components of muscle tissue surrounding the cells.
Overall comparisons of 1377 proteins or protein groups (proteins and protein groups are referred to as “proteins” in the following text for better readability) were quantified in our analysis using Progenesis QI for proteomics.
The baseline data of the two groups were compared against each other to identify any differences on the proteomic level due to the cohort composition itself, to prevent possible data misinterpretation due to cohort variability. In total, three proteins were significantly increased in PLA compared with COL prior the intervention due to biological variability (
Table 3). In COL, no higher abundant proteins could be identified compared with PLA. In order to avoid possible influences of the different baseline values, these proteins were excluded from further data analysis.
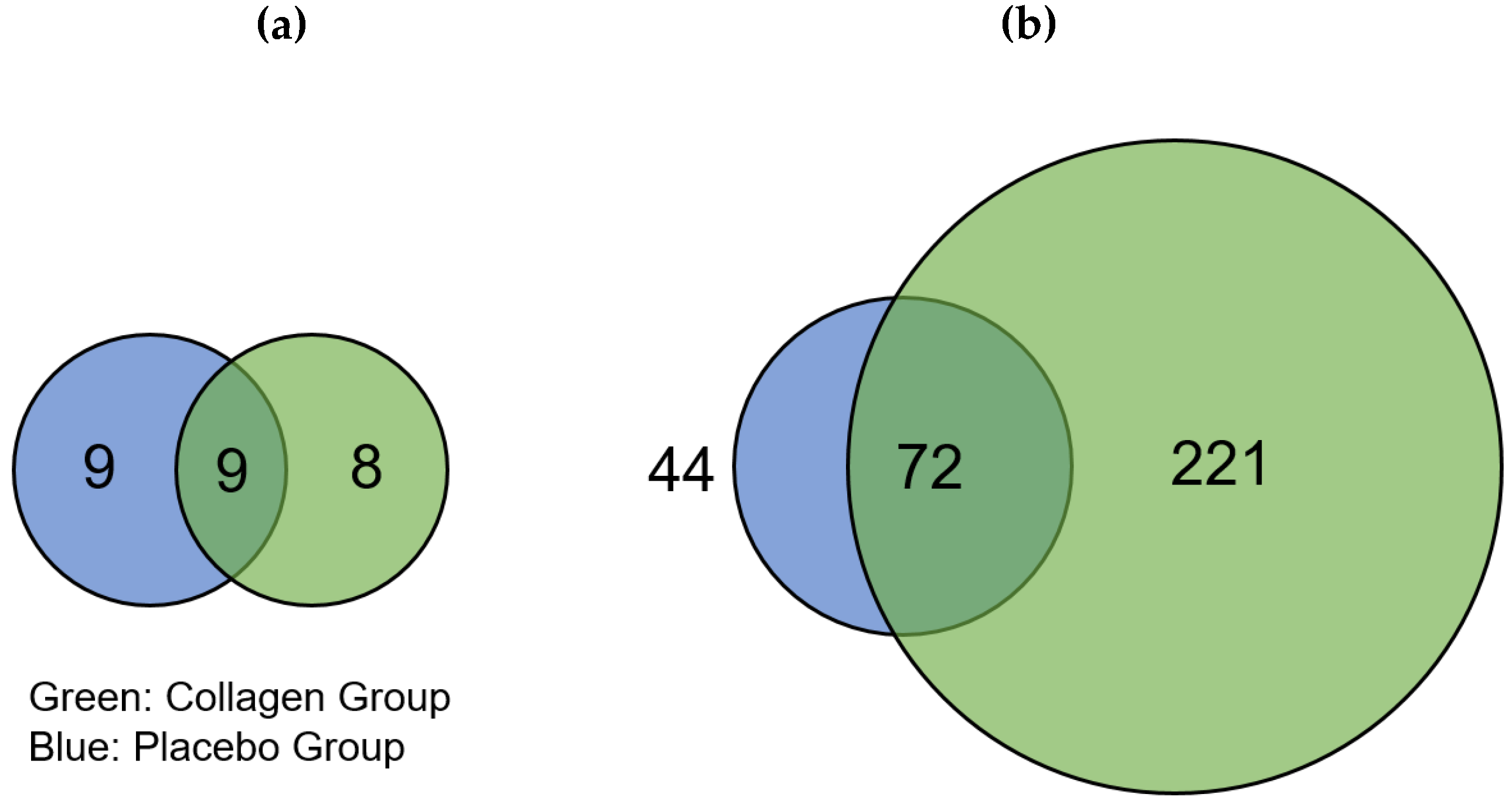
The distribution of regulated proteins before and after 12 weeks of resistance training and supplementation differed; there were 17 downregulated proteins in COL compared to 18 downregulated proteins in PLA, of which 9 proteins appeared in both groups (
Figure 2a). We found that 293 proteins were highly regulated in COL after the 12-week intervention, of which 72 were also of higher abundance in PLA. These changes thus resulted from the hypertrophy training intervention. Hence, 221 proteins were only upregulated in COL compared to 44 proteins in PLA, showing a pronounced difference in the number of upregulated proteins between the two groups (
Figure 2). The direct comparison between COL and PLA after 12 weeks of supplementation and resistance exercise training revealed no significant different proteins.
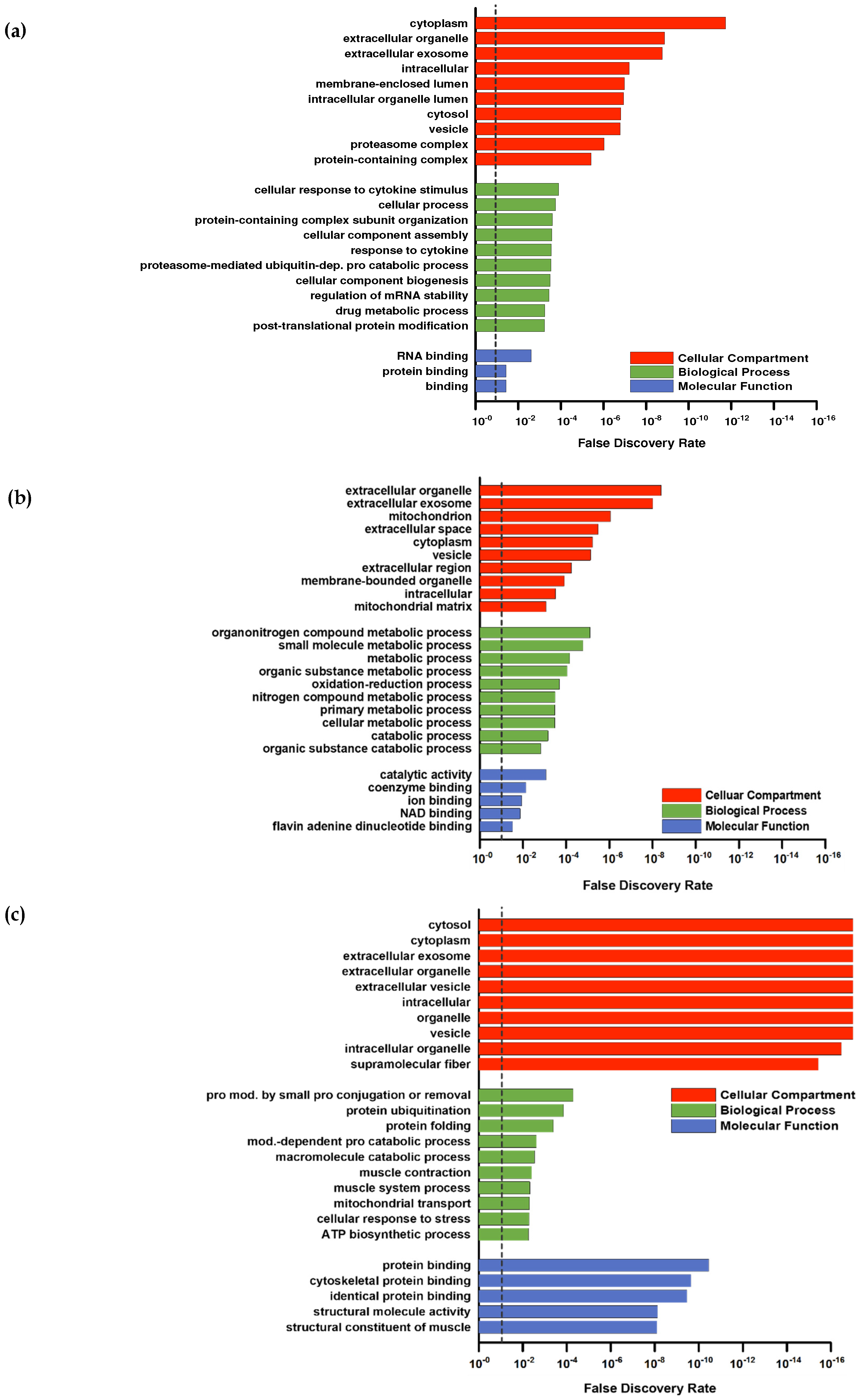
For a deeper proteomic characterization of our data set Gene Ontology (GO) categorization was performed using PANTHER (default settings using Fisher’s exact and FDR-correction) in respect to three different aspects: Cellular components, biological processes, and molecular functions [
37]. To provide a better overall view of the large number of hits, the REVIGO tool [
38] was used to sum classes ranked by FDR (
Figure 3a–c). The downregulated proteins (
Figure 2a) were not considered because the low number (PLA = 9; COL = 8) of hits was not appropriate for group allocations.
Higher abundant proteins in the intersection (
Figure 2b) were assigned to GO categories often found in response to resistance training. For example, upregulated proteins in the GO category “biological processes” (
Figure 3a) included the response to cytokines or protein modifications, which are in line with other study outcomes [
39,
40]. No subclass of the aspects “molecular functions” and “cellular components” showed specific categories associated with skeletal muscle. Upregulated proteins in PLA showed no direct association with the skeletal muscle system, although the upregulated proteins in PLA in the biological processes category that could be classified into organonitrogen compound metabolic processes and nitrogen compound metabolic processes indicated some responses to an amino acid stimulus (
Figure 3c).
In contrast, annotation of upregulated proteins of COL included those associated with GO biological processes related to the skeletal muscle (muscle contraction and muscle system process) and protein modifications (protein ubiquitination, protein modification by small protein conjugation or removal) (
Figure 3b).
The annotated GOs for cellular components were similar in COL and PLA: extracellular parts, cytoplasm, intracellular, and vesicles. However, PLA contained a higher number of upregulated proteins associated with the terms mitochondria and mitochondrial matrix (
Figure 3c), and COL had a few upregulated proteins related to the GO category supramolecular fibers, which are superior to contractile fibers (
Figure 3b).
The GOs belonging to molecular function in COL also included categories directly related to the skeletal muscle fiber structure, like cytoskeletal protein binding and the structural constitution of muscle. Upregulated proteins in PLA were assigned with general categories, such as catalytic activity and coenzyme binding, with respect to their molecular function.
Overall, COL showed a strong association with annotations spanning contractile muscle fiber (sarcomere or cytoskeleton) in all three GO categories, whereas PLA showed none of them or not enough regulated proteins to form a category. To summarize, we observed considerable differences in the protein response due to collagen supplementation, as both groups performed the same training protocol.
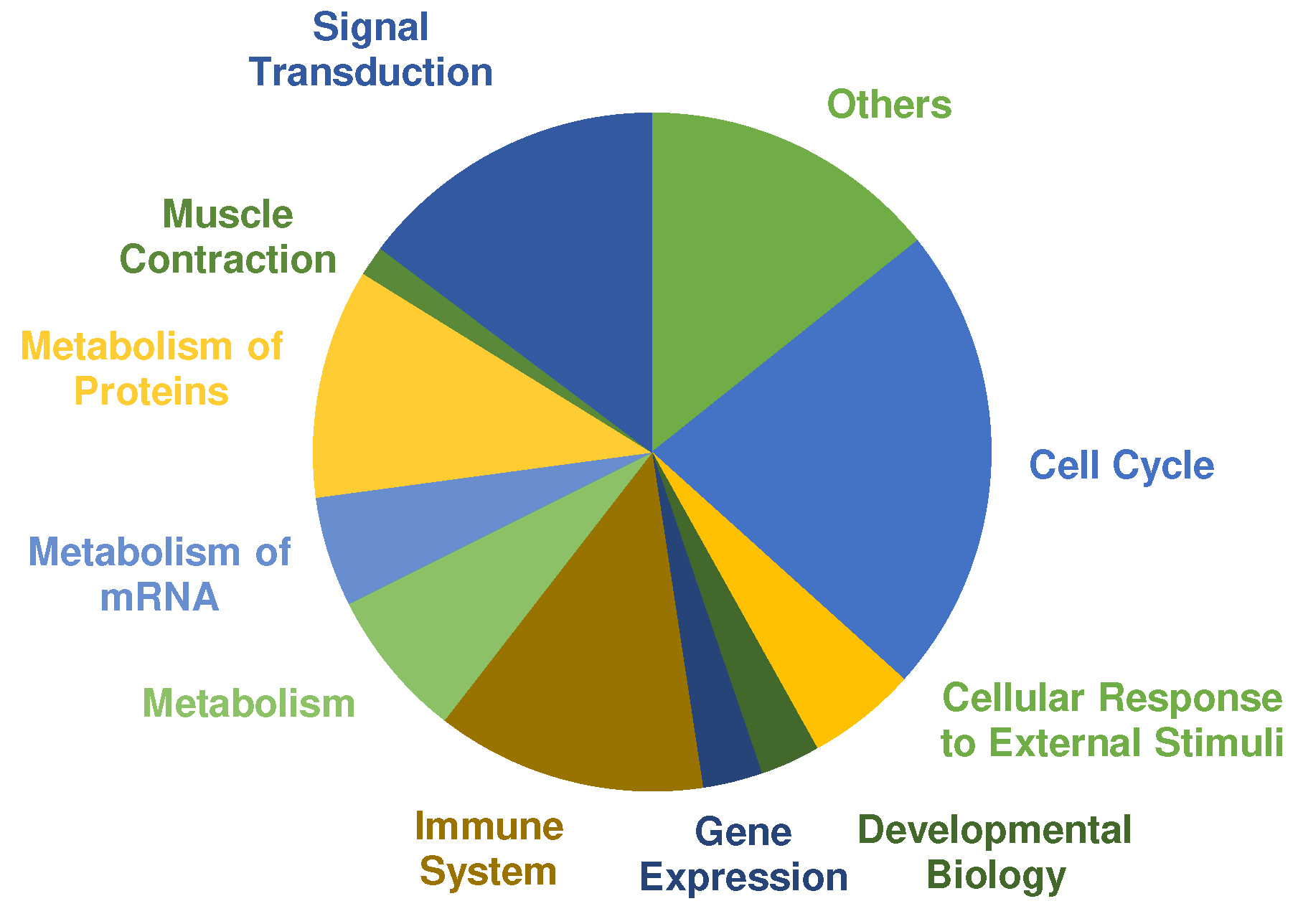
A pathway analysis was performed using PANTHERs overrepresentation test with default settings and the “Reactome pathways” annotated data set. The main categories of significant regulated pathways (Cut off values: FDR ≤ 0.01, fold enrichment score ≥ 2.00 and at least three or more proteins assigned) are presented in
Figure 4. Most pathways were annotated to cell cycle, signal transduction, metabolism of proteins and immune system. Selected pathways that are relevant in the context of exercise training and adaptation according to the literature are listed in
Table 3 (complete list in
supplemental Table S1). COL showed 192 pathways that are significantly overrepresented. In comparison, only one pathway was assigned to PLA according to the cut-off criteria, which was “metabolism” (fold enrichment = 3.70; FDR = 4.75 × 10
−3).
Overrepresented pathways in COL showed many responses characteristic to exercise training and adaptation (
Table 4). Of 192 significant pathways, 31 pathways were assigned to signal transduction. Several mitogen-activated protein kinases (MAPK) pathways were overrepresented that have been reported before in human skeletal muscle, after cycling or running exercise [
41,
42]. MAPK cascades are activated through cellular stress such as muscle contraction and exercise. In particular, MAPK1 and MAPK3, also known as extracellular-signal regulated kinase (ERK) 2 and 1, are the key components of the MAPK/ERK cascade, which regulates cell growth and differentiation through influencing transcription, translation, and cytoskeleton structure [
43]. Furthermore, signal transduction through protein kinase B (Akt) is upregulated in COL. The Akt/mTOR pathway is one of the most researched pathways on cell growth and is regarded as the main regulator of hypertrophy [
44]. As expected after exercise, the immune system response increased cytokine signaling through interleukins to improve the regeneration process [
45]. The metabolism of proteins belongs mainly to translational processes and protein folding, especially for cytoskeletal proteins. Overrepresented metabolic pathways are mainly glycolytic energy metabolism associated pathways as expected according to the chosen training stimulus [
46].
Cellular response to external stimuli pathway, showed downstream cascades responding to stress, hypoxia and heat stress. Hypoxia inducible factor alpha 1 (HIF-1) is regulated through muscle oxygen concentration that decreased during exercise or injury. HIF-1 stimulates via erythropoietin (EPO) myoblast proliferation and muscle recovery after injury and is important for muscle repair and regeneration [
47]. Furthermore, heat shock transcription factor 1 (HSF1) is present in multiple pathways. HSF1 acts as the major transcription factor in human muscle cells in response to stress. Striated and smooth muscle contraction pathways are also significantly overrepresented after 12 weeks of strength training and collagen supplementation.
To create an initial understanding of the related mechanisms, proteins belonging to the term contractile fibers (assigned according to PANTHER) were further analyzed, since proteins of this category were detected in both groups. Only two proteins from 44 differentially upregulated proteins of PLA were assigned to this type, in COL 26 out of 221 higher abundant proteins were annotated to the term contractile fibers (
Table 5 and
Table S2). One of the upregulated proteins in PLA was alpha-crystallin B chain, which often increases to protect the cytoskeleton and myofibril structure after muscle damage caused by eccentric exercise [
48]. The other protein belonging to this term detected in PLA was leiomodin-2, which promotes the polymerization of actin filaments in skeletal muscle cells [
49].
In COL several proteins with different myofibrillar function were identified as being in higher abundance compared to PLA, e.g., myosin proteins (myosin regulatory light chain 2, myosin-8, myosin light chain 1/3) and actin-binding proteins (alpha-actinin-2, synaptopodin, tropomodulin-4), including tropomyosins (tropomyosin beta chain, tropomyosin alpha-3 chain). Many have already been observed before in proteome analyses after resistance exercise [
50]. Troponin C and I were also found to be of higher abundance in COL. Troponin I is already known to be increased by chronic exercise and regulates the interaction between actin and myosin filaments [
51]. One of the upregulated proteins belonging to the Z-disk (five proteins total) is myotilin, an important marker for remodeling the myofibril structures after exercise. Myotilin binds alpha-actinin-2 and PDZ and LIM domain protein 3 to control Z-disk organization [
52]. Five sarcolemmal proteins showed an increased fold enrichment, including desmin, which has already been reported in other studies to be a responder to resistance exercise, which is relevant for force transmission through the cytoskeleton [
28]. The cytoskeleton is not just a structural component but is also responsible for the communication between the muscle cells and the ECM [
9]. Small muscular protein and smoothelin-like protein 1 were also upregulated in COL. These two proteins are associated with the costamere, being responsible for the force transmission, which is generated in the myofibrils and transferred to the ECM [
53].
COL induced a higher abundance in proteins being associated with resistance training adaptations. These effects might be caused by the high hydroxyproline-peptide content of the collagen peptide supplementation, as Kitakaze et al. [
54] were able to show an increase myoblast differentiation and myotube hypertrophy in murine skeletal muscle C2C12 cells by hydroxyproline-glycine-peptides.
The number of proteins belonging to relevant categories of the skeletal muscle (manually annotated) was significantly different between both groups, resulting in more relevant proteins for adaptation processes observed in COL compared with PLA.
Because participants in COL ingested 15 g collagen hydrolysate every day, collagen proteins and collagen-associated proteins that were differently expressed after 12 weeks are shown in
Table 6. Collagen 6alpha2, fibronectin, and laminin subunit gamma 1 showed increased FC in both groups after 12 weeks as a result of resistance training. Laminins, collagens, as well as fibronectins are the main fibrous parts of the ECM. Fibronectin has many different functions, for example, organization of ECM structures, cell migration, and adhesion and tissue repair through fibroblast migration [
55]. The increase in fibronectin after resistance training is consistent with other studies that reported increased values 72 h after exercise [
56]. An increase in laminin was associated with myogenic differentiation in mice [
57]. Consequently, both groups adapted their ECM protein production independently from collagen supplementation.
In contrast, collagen 5alpha1, collagen 15alpha1 and collagen 18alpha1 were only upregulated in COL, whereas PLA showed a fold enrichment of the protein lumican. Collagen 5alpha1 is a fibrillar collagen normally contained in collagen 1-rich regions, playing a critical role in fibrillogenesis and fibril organization [
58]. It is mostly associated with Ehlers-Danlos syndrome, which is caused by a congenital mutation of the
col5a1 gene and is characterized by hyperextensibility of skin and joints [
59]. Baghdadi et al. [
60] identified collagen 5alpha1 in muscle stem cells as a regulator of the notch signaling pathway that is concerned with the prevention of satellite cell storage by repressing differentiation process initiated by myoD (myogenic differentiation 1) to maintain quiescence, in addition to the main function of organizing the structure of the ECM through collagen 1 formation. The FC of collagen 5alpha1, which is extremely high compared with all other proteins, should be considered with caution because data validation (using immunoblotting) is still missing, and proteomic analysis itself struggles with a high variability between individuals [
61,
62].
The other protein belonging to the collagen family that was upregulated in COL is collagen 18alpha1. Together with collagen 15, it forms a collagen category named multiplexins, which is characterized by compounds with multiple triple-helix domains in containing non-collagenous parts. Collagen 18alpha1 is located next to collagen 4 in the basement membrane, occurs nearly ubiquitously in the human body, and seems to differ in function depending on the location [
63]. It contains a non-collagen domain called endostatin, which is known to be a suppressor of angiogenesis and tumor growth [
64]. Because of its inhibitory effect on tumor growth, endostatin has been well investigated in contrast to the function of collagen 18alpha1 in different tissues. Its role in skeletal muscle has not been directly investigated, but it is transferred from other tissues where it is the structural component of the basement membrane, binding ECM proteins like laminin.
Lumican, which was upregulated in PLA, is also a component of the ECM that binds to collagen 1 and affects collagen fibrillogenesis and therefore, the matrix assembly [
65].
Connective tissue remodeling has been studied in previous years, as
collagen 1, 3, and
4 genes are elevated after a single bout of exercise, but are unchanged after a second bout [
66]. After 36 training sessions, we would have expected a higher abundance in these proteins caused by adaptation, but they were unaffected in both groups, and instead, other collagenous proteins were upregulated in both groups. This shows their importance not only for contractile fibers but also for passive structures, like the ECM around muscle cells, to adapt to the mechanical load [
9]. The adaptation of the ECM seems to occur even without collagen supplementation, although more collagenous proteins were upregulated following COL supplementation.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}