Validation and Application of Biocrates AbsoluteIDQ® p180 Targeted Metabolomics Kit Using Human Milk

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. AbsoluteIDQ® p180 Assay and Sample Preparation
2.3. LC-MS
2.4. Method Validation
2.5. Human Milk Samples
2.6. Statistical Analysis
3. Results
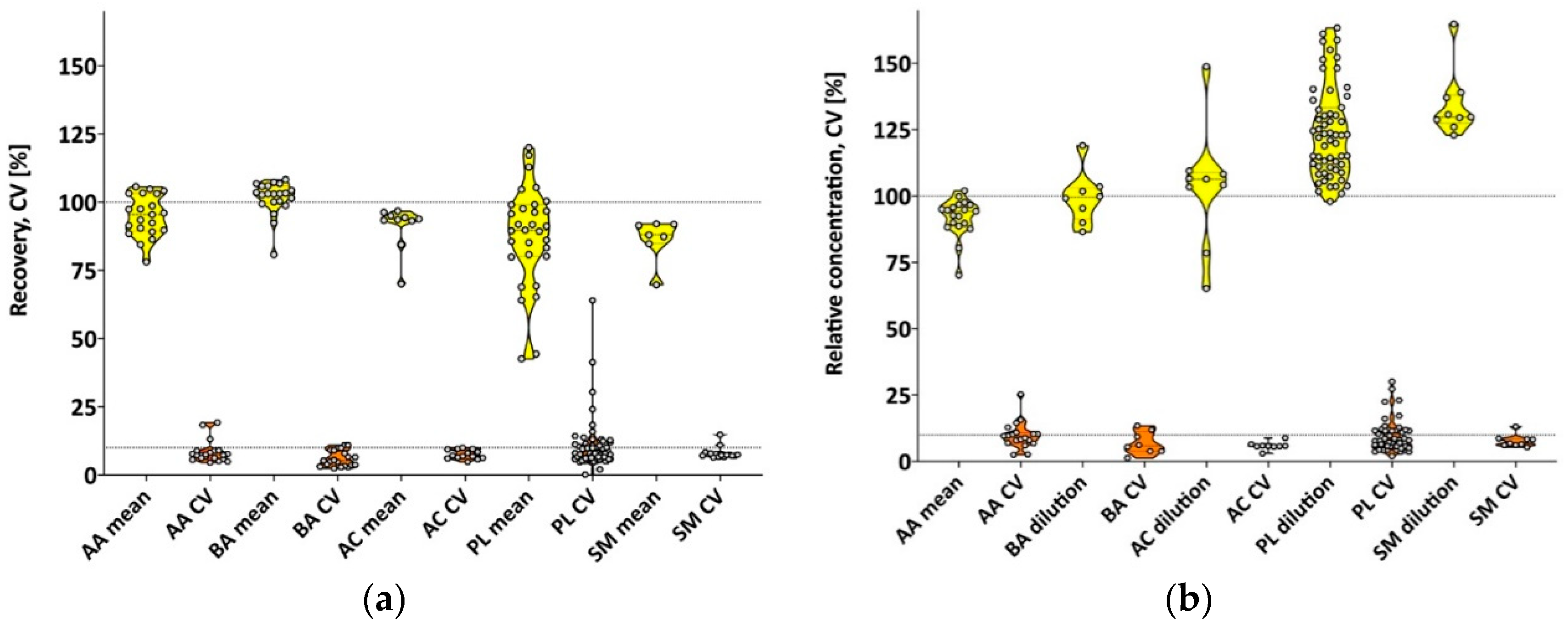
3.1. Validation Using Pooled Human Milk
3.1.1. LC-MS (Amino Acids and Biogenic Amines)
3.1.2. FIA (Acylcarnitines, Phospholipids, Sphingomyelins, Hexoses)
3.2. Feasibility Plate
4. Discussion
4.1. Validation Using Pooled Human Milk
4.2. Feasibility Plate
Differences by Group Assignment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slupsky, C. Metabolomics in Human Milk Research. In Human Milk: Composition, Clinical Benefits and Future Opportunities; Donovan, S.M., German, B.J., Lonnerdal, B., Lucas, A., Eds.; Karger AG: Basel, Switzerland, 2019; Volume 90, pp. 179–190. [Google Scholar]
- World Health Organization. The Optimal Duration of Exclusive Breastfeeding; Report of an Expert Consultation; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- Hampel, D.; York, E.R.; Allen, L.H. Ultra-performance liquid chromatography tandem mass-spectrometry (UPLC-MS/MS) for the rapid, simultaneous analysis of thiamin, riboflavin, flavin adenine dinucleotide, nicotinamide and pyridoxal in human milk. J. Chromatrogr. B Anal. Technol. Biomed. Life Sci. 2012, 903, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Shahab-Ferdows, S.; Domek, J.M.; Siddiqua, T.; Raqib, R.; Allen, L.H. Competitive chemiluminescent enzyme immunoassay for vitamin B12 analysis in human milk. Food Chem. 2014, 153, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Shahab-Ferdows, S.; Adair, L.S.; Bentley, M.E.; Flax, V.L.; Jamieson, D.J.; Ellington, S.R.; Tegha, G.; Chasela, C.S.; Kamwendo, D.; et al. Thiamin and Riboflavin in Human Milk: Effects of Lipid-Based Nutrient Supplementation and Stage of Lactation on Vitamer Secretion and Contributions to Total Vitamin Content. PLoS ONE 2016, 11, e0149479. [Google Scholar] [CrossRef] [PubMed]
- Redeuil, K.; Bénet, S.; Affolter, M.; Thakkar, S.; Campos-Giménez, E. A Novel Methodology for the Quantification of B-Vitamers in Breast Milk. J. Anal. Bioanal. Tech. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.N.; Yin, S.A.; Yang, Z.Y.; Yang, X.G.; Bing, S.; Ren, Y.P.; Zhang, J. Application of UPLC-MS/MS Method for Analyzing B-vitamins in Human Milk. Biomed. Environ. Sci. 2015, 28, 738–750. [Google Scholar] [PubMed]
- Lildballe, D.L.; Hardlei, T.F.; Allen, L.H.; Nexo, E. High concentrations of haptocorrin interfere with routine measurement of cobalamins in human serum and milk. A problem and its solution. Clin. Chem. Lab. Med. 2009, 47, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Shahab-Ferdows, S.; Gertz, E.; Flax, V.L.; Adair, L.S.; Bentley, M.E.; Jamieson, D.J.; Tegha, G.; Chasela, C.S.; Kamwendo, D.; et al. The effects of a lipid-based nutrient supplement and antiretroviral therapy in a randomized controlled trial on iron, copper, and zinc in milk from HIV-infected Malawian mothers and associations with maternal and infant biomarkers. Matern. Child. Nutr. 2017, 14. [Google Scholar] [CrossRef]
- Redeuil, K.; Vulcano, J.; Prencipe, F.P.; Bénet, S.; Campos-Giménez, E.; Meschiari, M. First quantification of nicotinamide riboside with B3 vitamers and coenzymes secreted in human milk by liquid chromatography-tandem-mass spectrometry. J. Chrom. B Analyt. Technol. Biomed. Life Sci. 2019, 1110–1111, 74–80. [Google Scholar] [CrossRef]
- Huang, J.; Kailemia, M.J.; Goonatilleke, E.; Parker, E.A.; Hong, Q.; Sabia, R.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Quantitation of human milk proteins and their glycoforms using multiple reaction monitoring (MRM). Anal Bioanal. Chem. 2017, 409, 589–606. [Google Scholar] [CrossRef]
- Wu, S.; Tao, N.; German, J.B.; Grimm, R.; Lebrilla, C.B. Development of an annotated library of neutral human milk oligosaccharides. J. Proteome Res. 2010, 9, 4138–4151. [Google Scholar] [CrossRef]
- Fernández, L.; Langa, S.; Martín, V.; Maldonado, A.; Jiménez, E.; Martín, R.; Rodríguez, J.M. The human milk microbiota: Origin and potential roles in health and disease. Pharmacol. Res. 2013, 69, 1–10. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.K.; McGuire, M.A. Got bacteria? The astounding, yet not-so-surprising, microbiome of human milk. Cur. Opin. Biotechnol. 2017, 44, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, E.; Ulven, T.; Færgeman, N.J.; Ejsing, C.S. Comprehensive and quantitative profiling of lipid species in human milk, cow milk and a phospholipid-enriched milk formula by GC and MS/MSALL. Eur. J. Lipid Sci. Technol. 2015, 117, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Sinanoglou, V.J.; Cavouras, D.; Boutsikou, T.; Briana, D.D.; Lantzouraki, D.Z.; Paliatsiou, S.; Volaki, P.; Bratakos, S.; Malamitsi-Puchner, A.; Zoumpoulakis, P. Factors affecting human colostrum fatty acid profile: A case study. PLoS ONE 2017, 12, e0175817. [Google Scholar] [CrossRef] [PubMed]
- Marincola, F.C.; Dessì, A.; Corbu, S.; Reali, A.; Fanos, V. Clinical impact of human breast milk metabolomics. Clin. Chim. Acta 2015, 451, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Gay, M.; Koleva, P.; Slupsky, C.; Toit, E.; Eggesbo, M.; Johnson, C.; Wegienka, G.; Shimojo, N.; Campbell, D.; Prescott, S.; et al. Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle? Nutrients 2018, 10, 1151. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B. Milk—A nutrient system of mammalian evolution promoting mTORC1-dependent translation. Int. J. Mol. Sci. 2015, 16, 17048–17087. [Google Scholar] [CrossRef] [PubMed]
- Russell, H.; Taylor, P.M.; Hundal, H.S. Amino acid transporters: Roles in amino acid sensing and signalling in animal cells. Biochem. J. 2003, 373, 1–18. [Google Scholar]
- Moran-Ramos, S.; Ocampo-Medina, E.; Gutierrez-Aguilar, R.; Macías-Kauffer, L.; Villamil-Ramírez, H.; López-Contreras, B.E.; León-Mimila, P.; Vega-Badillo, J.; Gutierrez-Vidal, R.; Villarruel-Vazquez, R.; et al. An amino acid signature associated with obesity predicts 2-year risk of hypertriglyceridemia in school-age children. Sci. Rep. 2017, 7, 5607. [Google Scholar] [CrossRef]
- Isganaitis, E.; Venditti, S.; Matthews, T.J.; Lerin, C.; Demerath, E.W.; Fields, D.A. Maternal obesity and the human milk metabolome: Associations with infant body composition and postnatal weight gain. Am. J. Clin. Nutr. 2019, 110, 111–120. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lönnerdal, B.; Slupsky, C.M. The human milk metabolome reveals diverse oligosaccharide profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.M.; Rudolph, M.C.; McManaman, J.L.; Neville, M.C. Key stages in mammary gland development. Secretory activation in the mammary gland: It’s not just about milk protein synthesis! Breast Cancer Res. 2007, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, S.L.; Stoeggl, W.M.; Weinberger, K.M.; Graber, A.; Guggenbichler, W. Apparatus and method for analyzing a metabolite profile. Google Patents US8265877 B2, 11 September 2012. [Google Scholar]
- Ramsay, S.L.; Guggenbichler, W.; Weinberger, K.M.; Graber, A.; Stoeggl, W.M. Device for Quantitative Analysis of a Drug or Metabolite Profile. Google Patents US8116983 B2, 14 February 2012. [Google Scholar]
- Rist, M.J.; Roth, A.; Frommherz, L.; Weinert, C.H.; Krüger, R.; Merz, B.; Bunzel, D.; Mack, C.; Egert, B.; Bub, A.; et al. Metabolite patterns predicting sex and age in participants of the Karlsruhe Metabolomics and Nutrition (KarMeN) study. PLoS ONE 2017, 12, e0183228. [Google Scholar] [CrossRef] [PubMed]
- Klavins, K.; Koal, T.; Dallmann, G.; Marksteiner, J.; Kemmler, G.; Humpel, C. The ratio of phosphatidylcholines to lysophosphatidylcholines in plasma differentiates healthy controls from patients with Alzheimer’s disease and mild cognitive impairment. Alzheimers Dement 2015, 1, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Stefan, N.; Yu, Z.; Mühlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Häring, H.U.; de Angelis, M.H.; Peters, A.; et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Shardell, M.; Ashour, F.A.S.; Moaddel, R.; Trehan, I.; Maleta, K.M.; Ordiz, M.I.; Kraemer, K.; Khadeer, M.A.; Ferrucci, L.; et al. Child stunting is associated with low circulating essential amino acids. EBioMedicine 2016, 6, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Dewey, K.G.; Begum, K. Long-term consequences of stunting in early life. Matern. Child. Nutr. 2011, 7, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Instrument | Parameter | LC-MS | FIA (MS Only) |
|---|---|---|---|
| 5500 QTRAP MS | CUR | 45 | 20 |
| IS | 5500 | 5500 | |
| TEM | 500 | 200 | |
| GS1 | 60 | 40 | |
| GS2 | 70 | 50 | |
| CAD | 9 | 9 | |
| EP | 10 | 10 | |
| CXP | 15 | 15 | |
| ACQUITY UPLC | Time [min] | Flow rate [mL/min] | Solvent A [%] |
| LC-MS | 0 | 0.8 | 100 |
| 0.45 | 0.8 | 100 | |
| 3.3 | 0.8 | 85 | |
| 5.9 | 0.8 | 30 | |
| 6.05 | 0.8 | 0 | |
| 6.2 | 0.9 | 0 | |
| 6.42 | 0.9 | 0 | |
| 6.52 | 0.8 | 0 | |
| 6.7 | 0.8 | 100 | |
| 7.3 | 0.8 | 100 | |
| FIA | 0 | 0.03 | Biocrates Solvent I MS running buffer in isocratic mode |
| 1.6 | 0.03 | ||
| 2.4 | 0.2 | ||
| 2.8 | 0.2 | ||
| 3.0 | 0.03 |
| Metabolite 1 | BMI > 18.5 Mothers | Stunted Infants | p-Value 2 |
|---|---|---|---|
| [µmol/L] | |||
| C 5 | 0.13 (0.12, 0.14) | 0.19 (0.17, 0.37) | 0.011 |
| PC aa C36:6 | 0.028 (0.025, 0.028) | 0.031 (0.028, 0.035) | 0.044 |
| PC ae C30:2 | 0.011 (0.01, 0.011) | 0.012 (0.012, 0.013) | 0.047 |
| SM C22:3 | 0.017 (0.014, 0.019) | 0.022 (0.018, 0.037) | 0.014 |
| Citrulline | 18.5 (12.2, 22.3) | 10.6 (8.7, 16.3) | 0.021 |
| Glutamate | 1825 (1477, 1977) | 1181 (976, 1472) | 0.002 |
| Glycine | 116 (103, 147) | 96.0 (79.5, 104) | 0.017 |
| Phenylalanine | 12.5 (11.7, 16.1) | 11.2 (10.3, 12.0) | 0.012 |
| Serine | 142 (94.2, 184) | 85.0 (79.4, 108) | 0.026 |
| Sarcosine | 0.61 (0.52, 0.72) | 0.46 (0.38, 0.51) | 0.044 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hampel, D.; Shahab-Ferdows, S.; Hossain, M.; Islam, M.M.; Ahmed, T.; Allen, L.H. Validation and Application of Biocrates AbsoluteIDQ® p180 Targeted Metabolomics Kit Using Human Milk. Nutrients 2019, 11, 1733. https://doi.org/10.3390/nu11081733
Hampel D, Shahab-Ferdows S, Hossain M, Islam MM, Ahmed T, Allen LH. Validation and Application of Biocrates AbsoluteIDQ® p180 Targeted Metabolomics Kit Using Human Milk. Nutrients. 2019; 11(8):1733. https://doi.org/10.3390/nu11081733
Chicago/Turabian StyleHampel, Daniela, Setareh Shahab-Ferdows, Muttaquina Hossain, M. Munirul Islam, Tahmeed Ahmed, and Lindsay H. Allen. 2019. "Validation and Application of Biocrates AbsoluteIDQ® p180 Targeted Metabolomics Kit Using Human Milk" Nutrients 11, no. 8: 1733. https://doi.org/10.3390/nu11081733
APA StyleHampel, D., Shahab-Ferdows, S., Hossain, M., Islam, M. M., Ahmed, T., & Allen, L. H. (2019). Validation and Application of Biocrates AbsoluteIDQ® p180 Targeted Metabolomics Kit Using Human Milk. Nutrients, 11(8), 1733. https://doi.org/10.3390/nu11081733