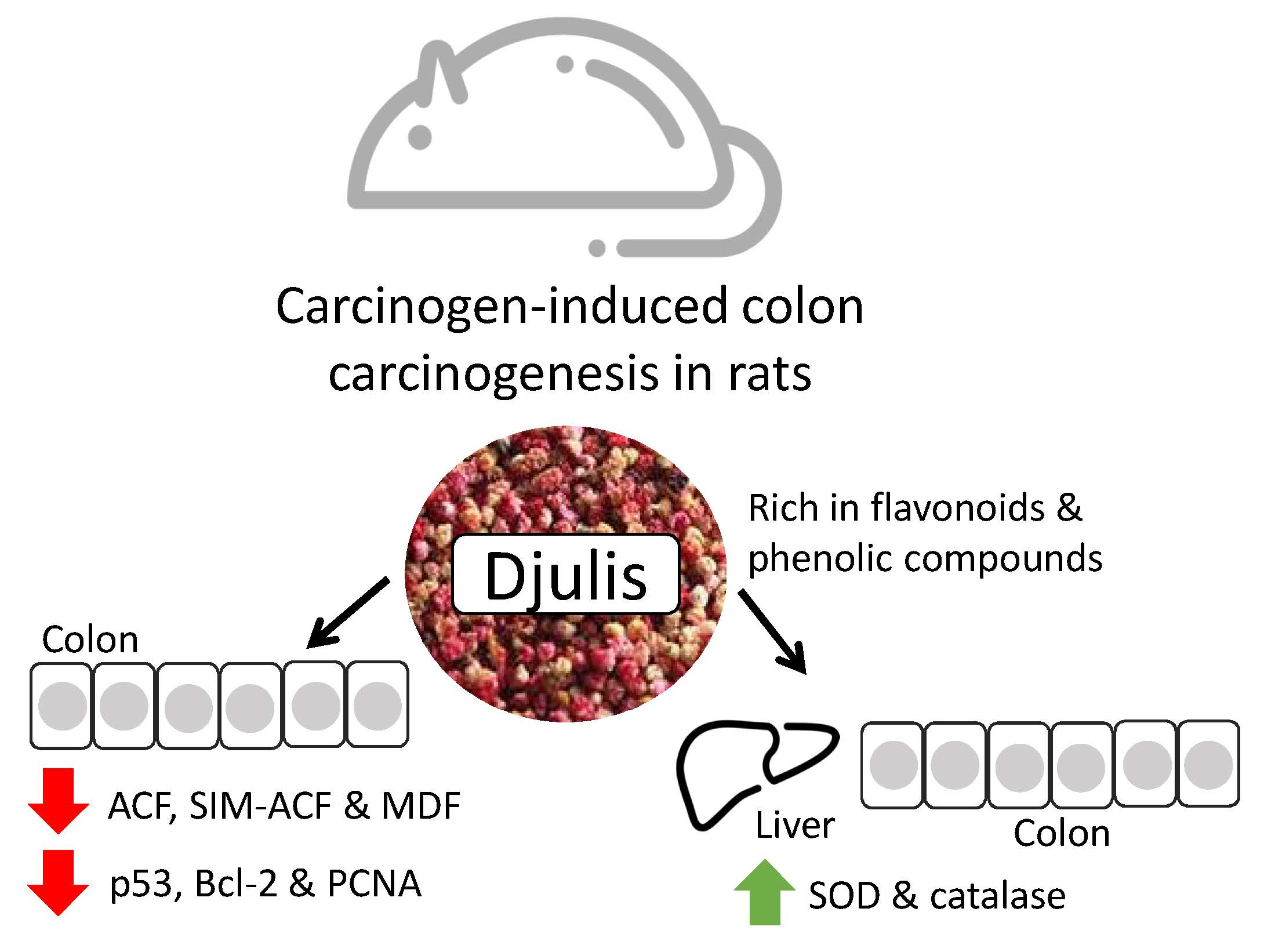
Djulis (Chenopodium Formosanum) Prevents Colon Carcinogenesis via Regulating Antioxidative and Apoptotic Pathways in Rats



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design
2.3. Aberrant Crypt Foci (ACF) Counts in the Colon
2.4. Identification of Mucin-Producing Aberrant Crypt Foci (ACF) and Mucin-Depleted Foci (MDF)
2.5. Measuring Activities of Antioxidant Enzymes
2.6. Western Blot Analysis
2.7. Djulis Extraction
2.8. Determination of Total Phenolics Content in Djulis Samples
2.9. Determination of Total Flavonoids Content in Djulis Samples
2.10. Statistical Analysis
3. Results
3.1. Numbers of Colonic Aberrant Crypt Foci (ACF) and ACs
3.2. Mucin Production by Aberrant Crypt Foci (ACF) and Mucin-Depleted Foci (MDF)
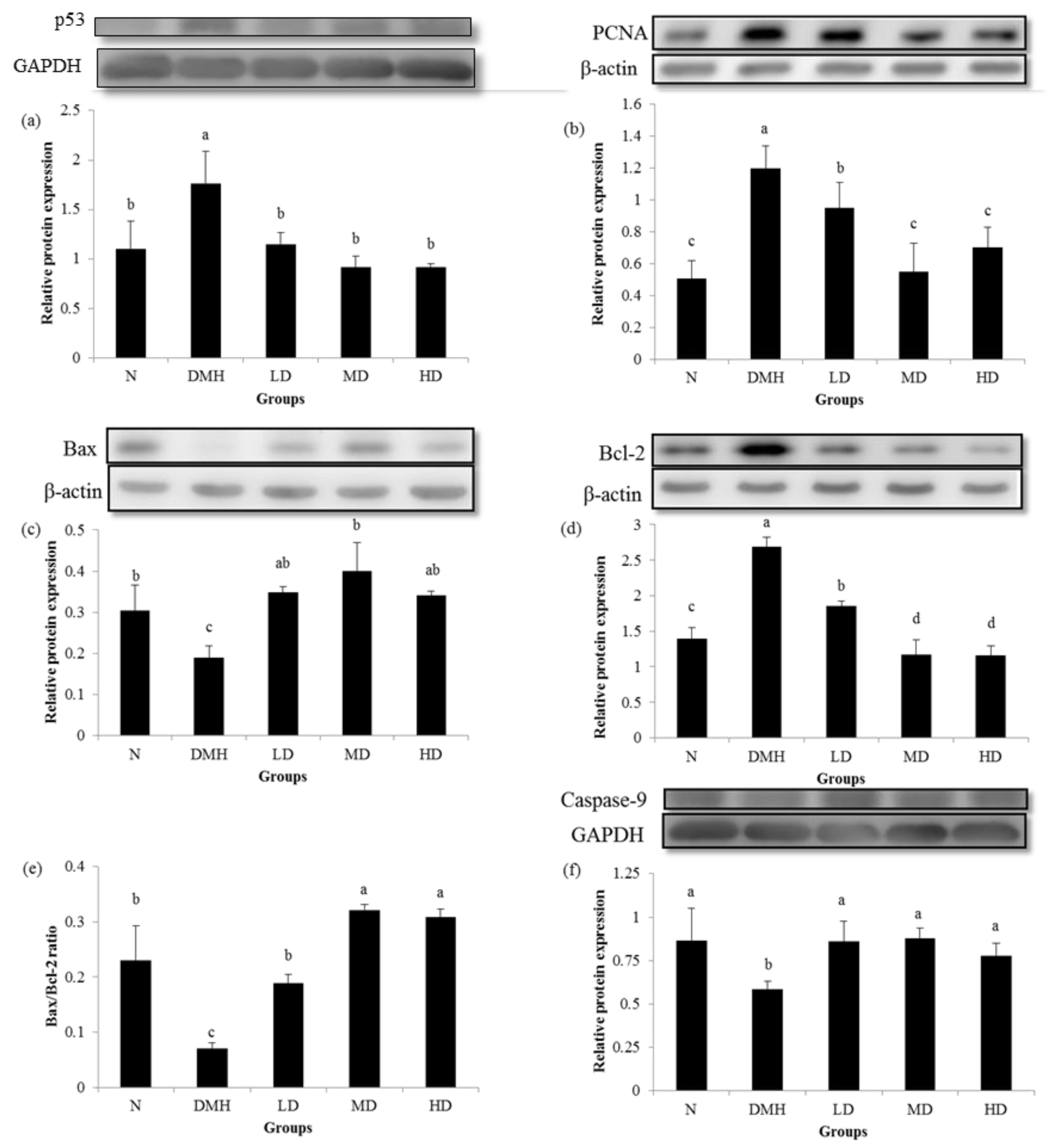
3.4. Expression of Proliferation- and Apoptosis-Related Proteins
3.5. Total Phenolics Content and Total Flavonoids Content in Djulis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Vargas, A.J.; Thompson, P.A. Diet and nutrient factors in colorectal cancer risk. Nutr. Clin. Pract. 2012, 27, 613–623. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Schwedhelm, C.; Hoffmann, G.; Knuppel, S.; Laure Preterre, A.; Iqbal, K.; Bechthold, A.; De Henauw, S.; Michels, N.; Devleesschauwer, B.; et al. Food groups and risk of colorectal cancer. Int. J. Cancer 2018, 142, 1748–1758. [Google Scholar] [CrossRef]
- Batra, P.; Sharma, A.K. Anti-cancer potential of flavonoids: Recent trends and future perspectives. 3 Biotech 2013, 3, 439–459. [Google Scholar] [CrossRef]
- Rosa, L.; Silva, N.; Soares, N.; Monteiro, M.; Teodoro, A.J. Anticancer properties of phenolic acids in colon cancer–a review. J. Nutr. Food Sci. 2016, 6, 10–4172. [Google Scholar]
- Guruswamy, S.; Swamy, M.V.; Choi, C.I.; Steele, V.E.; Rao, C.V. S-Adenosyl l-methionine inhibits azoxymethane‐induced colonic aberrant crypt foci in F344 rats and suppresses human colon cancer Caco‐2 cell growth in 3D culture. Int. J. Cancer 2008, 122, 25–30. [Google Scholar] [CrossRef]
- Perše, M.; Cerar, A. Morphological and molecular alterations in 1, 2 dimethylhydrazine and azoxymethane induced colon carcinogenesis in rats. J. Biomed. Biotechnol. 2010, 2011, 473964. [Google Scholar] [CrossRef]
- Kim, S.; Guo, J.; O'Sullivan, M.G.; Gallaher, D.D.; Turesky, R.J. Comparative DNA adduct formation and induction of colonic aberrant crypt foci in mice exposed to 2-amino-9H-pyrido [2, 3-b] indole, 2-amino-3, 4-dimethylimidazo [4, 5-f] quinoline, and azoxymethane. Env. Mol. Mutagen. 2016, 57, 125–136. [Google Scholar] [CrossRef]
- Jenab, M.; Chen, J.M.; Thompson, L.U. Sialomucin production in aberrant crypt foci relates to degree of dysplasia and rate of cell proliferation. Cancer Lett. 2001, 165, 19–25. [Google Scholar] [CrossRef]
- Femia, A.P.; Bendinelli, B.; Giannini, A.; Salvadori, M.; Pinzani, P.; Dolara, P.; Caderni, G. Mucin-depleted foci have β-catenin gene mutations, altered expression of its protein, and are dose-and time-dependent in the colon of 1, 2-dimethylhydrazine-treated rats. Int. J. Cancer 2005, 116, 9–15. [Google Scholar] [CrossRef]
- Pan, M.H.; Lai, C.S.; Wu, J.C.; Ho, C.T. Molecular mechanisms for chemoprevention of colorectal cancer by natural dietary compounds. Mol. Nutr. Food Res. 2011, 55, 32–45. [Google Scholar] [CrossRef]
- Perše, M. Oxidative stress in the pathogenesis of colorectal cancer: Cause or consequence? Biomed Res. Int. 2013, 2013, 725710. [Google Scholar] [CrossRef]
- Hamiza, O.O.; Rehman, M.U.; Tahir, M.; Khan, R.; Khan, A.Q.; Lateef, A.; Ali, F.; Sultana, S. Amelioration of 1, 2 dimethylhydrazine (DMH) induced colon oxidative stress, inflammation and tumor promotion response by tannic acid in Wistar rats. Asian Pac. J. Cancer Prev. 2012, 13, 4393–4402. [Google Scholar] [CrossRef]
- Chyau, C.-C.; Chu, C.-C.; Chen, S.-Y.; Duh, P.-D. The inhibitory effects of djulis (Chenopodium formosanum) and its bioactive compounds on adipogenesis in 3T3-L1 adipocytes. Molecules 2018, 23, 1780. [Google Scholar] [CrossRef]
- Chu, C.-C.; Chen, S.-Y.; Chyau, C.-C.; Fu, Z.-H.; Liu, C.-C.; Duh, P.-D. Protective effect of djulis (Chenopodium formosanum) and its bioactive compounds against carbon tetrachloride-induced liver injury, in vivo. J. Funct. Foods 2016, 26, 585–597. [Google Scholar] [CrossRef]
- Lin, T.A.; Ke, B.J.; Cheng, C.S.; Wang, J.J.; Wei, B.L.; Lee, C.L. Red quinoa bran extracts protects against carbon tetrachloride-induced liver injury and fibrosis in mice via activation of antioxidative enzyme systems and blocking FGF-beta1 pathway. Nutrients 2019, 11, 395. [Google Scholar] [CrossRef]
- Tsai, P.-J.; Sheu, C.-H.; Wu, P.-H.; Sun, Y.-F. Thermal and pH stability of betacyanin pigment of djulis (Chenopodium formosanum) in Taiwan and their relation to antioxidant activity. J. Agric. Food Chem. 2009, 58, 1020–1025. [Google Scholar] [CrossRef]
- Tsai, P.-J.; Chen, Y.-S.; Sheu, C.-H.; Chen, C.-Y. Effect of nanogrinding on the pigment and bioactivity of djulis (Chenopodium formosanum koidz.). J. Agric. Food Chem. 2011, 59, 1814–1820. [Google Scholar] [CrossRef]
- Alonso-Castro, A.J.; Domínguez, F.; García-Carrancá, A. Rutin exerts antitumor effects on nude mice bearing SW480 tumor. Arch. Med. Res. 2013, 44, 346–351. [Google Scholar] [CrossRef]
- Matsunaga, K.; Katayama, M.; Sakata, K.; Kuno, T.; Yoshida, K.; Yamada, Y.; Hirose, Y.; Yoshimi, N.; Mori, H. Inhibitory effects of chlorogenic acid on azoxymethane-induced colon carcinogenesis in male F344 rats. Asian Pac. J. Cancer Prev. 2002, 3, 163–166. [Google Scholar]
- Li, S.C.; Chen, C.M.; Lin, S.H.; Chiang, W.; Shih, C.K. Effects of adlay bran and its ethanolic extract and residue on preneoplastic lesions of the colon in rats. J. Sci. Food Agric. 2011, 91, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-L.; Lu, C.-K.; Huang, Y.-J.; Chen, H.-J. Antioxidative caffeoylquinic acids and flavonoids from Hemerocallis fulva flowers. J. Agric. Food Chem. 2011, 59, 8789–8795. [Google Scholar] [CrossRef] [PubMed]
- Lau, D.S.; Archer, M.C. Fatty acid synthase is over-expressed in large aberrant crypt foci in rats treated with azoxymethane. Int. J. Cancer 2009, 124, 2750–2753. [Google Scholar] [CrossRef] [PubMed]
- Femia, A.P.; Dolara, P.; Caderni, G. Mucin-depleted foci (MDF) in the colon of rats treated with azoxymethane (AOM) are useful biomarkers for colon carcinogenesis. Carcinogenesis 2004, 25, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Tang, B.; Zhou, L.; Jikihara, H.; Kiwata, A.; Sakamoto, Y.; Tang, F.; Xiao, S.; Wang, Z.; Wu, Q. Effects of high-fat diet on 1, 2-dimethylhydrazine-induced aberrant crypt foci and colorectal tumours in rats. Biomed. Rep. 2015, 3, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Kozaki, K.i.; Nishikawa, Y.; Yamamoto, M.; Fukami, H.; Inoue, M.; Wakabayashi, K.; Tatematsu, M. Development and distribution of 2-amino-1-methyl-6-phenylimidazo [4, 5-b]-pyridine (PHIP)-induced aberrant crypt foci in the rat large intestine. Jpn. J. Cancer Res. 1999, 90, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Kasdagly, M.; Radhakrishnan, S.; Reddivari, L.; Veeramachaneni, D.R.; Vanamala, J. Colon carcinogenesis: Influence of western diet-induced obesity and targeting stem cells using dietary bioactive compounds. Nutrition 2014, 30, 1242–1256. [Google Scholar] [CrossRef] [PubMed]
- Jass, J.; Walsh, M. Altered mucin expression in the gastrointestinal tract: A review. J. Cell Mol. Med. 2001, 5, 327–351. [Google Scholar] [CrossRef] [PubMed]
- Milosevic, V.; Vukmirovic, F.; Zindovic, M.; Krstic, M.; Milenkovic, S.; Jancic, S. Interplay between expression of leptin receptors and mucin histochemical aberrations in colorectal adenocarcinoma. Rom. J. Morphol. Embryol. 2015, 56, 709–716. [Google Scholar] [PubMed]
- Uchida, K.; Kado, S.; Ando, M.; Nagata, Y.; Takagi, A.; Onoue, M. A mucinous histochemical study on malignancy of aberrant crypt foci (ACF) in rat colon. J. Vet. Med. Sci. 2001, 63, 145–149. [Google Scholar] [CrossRef]
- Cui, C.; Takamatsu, R.; Doguchi, H.; Matsuzaki, A.; Saio, M.; Yoshimi, N. Pre-neoplastic lesion, mucin-depleted foci, reveals de novo high-grade dysplasia in rat colon carcinogenesis. Oncol. Rep. 2012, 27, 1365–1370. [Google Scholar] [PubMed]
- Chang, D.; Wang, F.; Zhao, Y.-S.; Pan, H.-Z. Evaluation of oxidative stress in colorectal cancer patients. Biomed. Env. Sci. 2008, 21, 286–289. [Google Scholar] [CrossRef]
- Hamiza, O.; Rehman, M.; Khan, R.; Tahir, M.; Khan, A.; Lateef, A.; Sultana, S. Chemopreventive effects of aloin against 1, 2-dimethylhydrazine-induced preneoplastic lesions in the colon of wistar rats. Hum. Exp. Toxicol. 2014, 33, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Bahnassy, A.A.; Zekri, A.; Salem, S.E.; Abou-Bakr, A.A.; Sakr, M.A.; Abdel-Samiaa, A.G.; Al-Bradei, M. Differential expression of p53 family proteins in colorectal adenomas and carcinomas: Prognostic and predictive values. Histol. Histopathol. 2014, 29, 207–216. [Google Scholar] [PubMed]
- Li, X.-L.; Zhou, J.; Chen, Z.-R.; Chng, W.-J. P53 mutations in colorectal cancer-molecular pathogenesis and pharmacological reactivation. World J. Gastroenterol. 2015, 21, 84. [Google Scholar] [CrossRef]
- Sarotra, P.; Kansal, S.; Sandhir, R.; Agnihotri, N. Chemopreventive effect of different ratios of fish oil and corn oil on prognostic markers, DNA damage and cell cycle in colon carcinogenesis. Eur. J. Cancer Prev. 2012, 21, 147–154. [Google Scholar] [CrossRef]
- Vikhanskaya, F.; Lee, M.K.; Mazzoletti, M.; Broggini, M.; Sabapathy, K. Cancer-derived p53 mutants suppress p53-target gene expression—Potential mechanism for gain of function of mutant p53. Nucleic Acids Res. 2007, 35, 2093–2104. [Google Scholar] [CrossRef]
- Saini, M.K.; Sanyal, S.N.; Vaiphei, K. Piroxicam and C-phycocyanin mediated apoptosis in 1, 2-dimethylhydrazine dihydrochloride induced colon carcinogenesis: Exploring the mitochondrial pathway. Nutr. Cancer 2012, 64, 409–418. [Google Scholar] [CrossRef]
- Kim, W.K.; Bang, M.H.; Kim, E.S.; Kang, N.E.; Jung, K.C.; Cho, H.J.; Park, J.H. Quercetin decreases the expression of ErbB2 and ErbB3 proteins in HT-29 human colon cancer cells. J. Nutr. Biochem. 2005, 16, 155–162. [Google Scholar] [CrossRef]
- Shen, X.G.; Wang, C.; Li, Y.; Wang, L.; Zhou, B.; Xu, B.; Jiang, X.; Zhou, Z.G.; Sun, X.F. Downregulation of caspase-9 is a frequent event in patients with stage II colorectal cancer and correlates with poor clinical outcome. Colorectal Dis. 2010, 12, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Hajrezaie, M.; Shams, K.; Moghadamtousi, S.Z.; Karimian, H.; Hassandarvish, P.; Emtyazjoo, M.; Zahedifard, M.; Majid, N.A.; Ali, H.M.; Abdulla, M.A. Chemoprevention of colonic aberrant crypt foci by novel schiff based dichlorido (4-methoxy-2-{[2-(piperazin-4-ium-1-yl) ethyl] iminomethyl} phenolate) cd complex in azoxymethane-induced colorectal cancer in rats. Sci. Rep. 2015, 5, 12379. [Google Scholar] [CrossRef] [PubMed]
- Shwter, A.N.; Abdullah, N.A.; Alshawsh, M.A.; Alsalahi, A.; Hajrezaei, M.; Almaqrami, A.A.; Salem, S.D.; Abdulla, M.A. Chemoprevention of colonic aberrant crypt foci by Gynura procumbens in rats. J. Ethnopharmacol. 2014, 151, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- Almagrami, A.A.; Alshawsh, M.A.; Saif-Ali, R.; Shwter, A.; Salem, S.D.; Abdulla, M.A. Evaluation of chemopreventive effects of Acanthus ilicifolius against azoxymethane-induced aberrant crypt foci in the rat colon. PLoS ONE 2014, 9, e96004. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xiong, L.-j.; Xu, F.; Zhao, X.; Liu, B.; Cai, K.-L.; Wang, G.-b. Estrogen inhibits colon polyp formation by reducing angiogenesis in a carcinogen-induced rat model. Int. J. Endocrinol. 2013, 2013, 453898. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Group 2 | ACF Incidence (% of Animals with ACF) | Number of ACF | Number of ACs | Crypt Multiplicity (Number of ACs/Focus) |
|---|---|---|---|---|
| DMH | 100% | 7.8 ± 1.3 a | 19.2 ± 3.4 a | 2.5 ± 0.3 |
| LD | 100% | 4.5 ± 1.1 b | 10.7 ± 2.4 b | 2.4 ± 0.2 |
| MD | 100% | 4.3 ± 0.7 bc | 11.2 ± 2.5 b | 2.6 ± 0.2 |
| HD | 100% | 3.4± 0.8 c | 8.7± 2.6 b | 2.5 ± 0.2 |
| Group 2 | Number of Focus Containing | Small ACF | Large ACF | ||
|---|---|---|---|---|---|
| 1 crypt | 2 crypts | 3 crypts | (≤3 crypts) | (≥4 crypts) | |
| DMH | 1.3 ± 0.6 a | 2.9 ± 0.9 a | 2.6 ± 0.7 a | 6.7 ± 1.1 a | 1.0 ± 0.6 |
| LD | 0.9 ± 0.4 b | 1.9 ± 0.7 b | 1.0 ± 0.3 bc | 3.8 ± 1.1 b | 0.7 ± 0.3 |
| MD | 0.6 ± 0.2 bc | 1.5 ± 0.4 b | 1.3 ± 0.4 b | 3.4 ± 0.5 bc | 0.9 ± 0.4 |
| HD | 0.4 ± 0.2 c | 1.5 ± 0.4 b | 0.8 ± 0.3 c | 2.7 ± 0.5 c | 0.7 ± 0.4 |
| Group 2 | Number of ACF Producing 3 | MDF | ||
|---|---|---|---|---|
| SUM | MIX | SIM | ||
| DMH | 8.6 ± 1.9 a | 1.4 ± 0.9 a | 1.8 ± 0.9 a | 0.7 ± 0.3 a |
| LD | 6.6 ± 2.9 ab | 0.5 ± 0.2 b | 0.4 ± 0.3 b | 0.1 ± 0.1 b |
| MD | 4.8 ± 1.9 b | 0.8 ± 0.3 b | 0.5 ± 0.4 b | 0.1 ± 0.1 b |
| HD | 5.1 ± 2.3 b | 0.7 ± 0.4 b | 0.4 ± 0.4 b | 0.1 ± 0.1 b |
| Group 2 | Colon | Liver | ||
|---|---|---|---|---|
| CAT 3 (U/mg Protein) | SOD 4 (U/mg Protein) | CAT (U/mg Protein) | SOD (U/mg Protein) | |
| N | 29.6 ± 7.7 c | 4.1 ± 0.7 b | 2409 ± 281 b | 30.6 ± 2.4 a |
| DMH | 20.8 ± 6.7 d | 3.0 ± 0.6 c | 1852 ± 251 c | 21.1 ± 3.7 b |
| LD | 29.9 ± 4.0 c | 3.4 ± 0.3 bc | 1649 ± 130 c | 20.1 ± 2.3 b |
| MD | 47.5 ± 2.9 b | 5.3 ± 0.3 a | 2328 ± 366 b | 27.9 ± 3.3 a |
| HD | 58.1 ± 6.2 a | 5.8 ± 0.9 a | 3282 ± 312 a | 28.5 ± 4.4 a |
| Sample | Free Form | Conjugated Form | Bound Form | Total Phenolics | ||
|---|---|---|---|---|---|---|
| Base-Hydrolyzable | Acid-Hydrolyzable | Base-Hydrolyzable | Acid-Hydrolyzable | |||
| Whole djulis | 666.92 ± 17.55 | 850.39 ± 51.28 | 1949.53 ± 54.31 | 1311.87 ± 11.00 | 438.38 ± 35.03 | 5217.10 |
| Djulis husk | 1823.30 ±25.08 | 2414.63 ± 62.61 | 4570.61 ± 77.72 | 1820.47 ± 20.55 | 96.40 ± 13.54 | 10725.40 |
| Djulis without husk | 308.61 ± 5.59 | 353.47 ± 6.25 | 1159.80 ± 11.90 | 421.87 ± 31.94 | 325.84 ± 11.14 | 2569.59 |
| Sample | Free Form | Conjugated Form | Bound Form | Total Flavonoids | ||
|---|---|---|---|---|---|---|
| Base-Hydrolyzable | Acid-Hydrolyzable | Base-Hydrolyzable | Acid-Hydrolyzable | |||
| Whole djulis | 582.25 ± 42.96 | 670.96 ± 31.78 | 750.75 ± 46.16 | 720.66 ± 27.11 | 373.88 ± 490.41 | 3098.80 |
| Djulis husk | 1904.03 ± 32.75 | 1790.28 ± 20.15 | 2282.50 ± 53.77 | 996.93 ± 21.82 | 509.38 ± 689.50 | 7483.10 |
| Djulis without husk | 296.26 ± 5.32 | 137.23 ± 12.60 | 363.41 ± 6.66 | 198.90 ± 14.00 | 106.45 ± 130.74 | 1102.24 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.-W.; Chen, H.-J.; Xie, G.-R.; Shih, C.-K. Djulis (Chenopodium Formosanum) Prevents Colon Carcinogenesis via Regulating Antioxidative and Apoptotic Pathways in Rats. Nutrients 2019, 11, 2168. https://doi.org/10.3390/nu11092168
Lee C-W, Chen H-J, Xie G-R, Shih C-K. Djulis (Chenopodium Formosanum) Prevents Colon Carcinogenesis via Regulating Antioxidative and Apoptotic Pathways in Rats. Nutrients. 2019; 11(9):2168. https://doi.org/10.3390/nu11092168
Chicago/Turabian StyleLee, Chih-Wei, Hong-Jhang Chen, Gui-Ru Xie, and Chun-Kuang Shih. 2019. "Djulis (Chenopodium Formosanum) Prevents Colon Carcinogenesis via Regulating Antioxidative and Apoptotic Pathways in Rats" Nutrients 11, no. 9: 2168. https://doi.org/10.3390/nu11092168