Cellular Oxidative Stress in Pediatric Leukemia and Lymphoma Patients Undergoing Treatment Is Associated with Protein Consumption

 ,
,
Abstract
1. Introduction
2. Materials and Methods
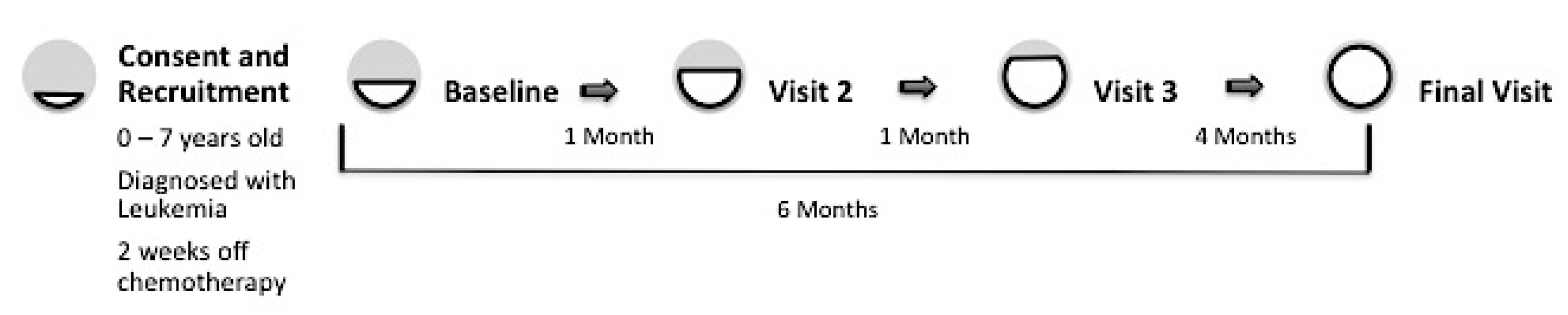
2.1. Participants and Procedure
2.2. Cells
2.3. Quantification of ROS
2.4. Quantification of Glutathione
2.5. Nutrition Measurements
2.6. Demographic and Anthropometric Measurements, and Assessment of Blood Counts
2.7. Response to Therapy
2.8. Statistical Analysis
3. Results
3.1. Participants
3.2. Changes in Oxidative Stress and Diet
3.3. Oxidative Stress Is Associated with Certain Dietary Components
3.4. Oxidative Stress and Clinical Characteristics
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.; Jürgens, H.; Frühwald, M.C. Important aspects of nutrition in children with cancer. Adv. Nutr. 2011, 2, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Lobato-Mendizábal, E.; López-Martínez, B.; Ruiz-Argüelles, G. A critical review of the prognostic value of the nutritional status at diagnosis in the outcome of therapy of children with acute lymphoblastic leukemia. Rev. Investig. Clin. Organo Hosp. Enferm. Nutr. 2003, 55, 31–35. [Google Scholar]
- Sala, A.; Rossi, E.; Antillon, F.; Molina, A.L.; de Maselli, T.; Bonilla, M.; Hernandez, A.; Ortiz, R.; Pacheco, C.; Nieves, R. Nutritional status at diagnosis is related to clinical outcomes in children and adolescents with cancer: A perspective from Central America. Eur. J. Cancer 2012, 48, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Israëls, T.; van de Wetering, M.D.; Hesseling, P.; van Geloven, N.; Caron, H.N.; Molyneux, E.M. Malnutrition and neutropenia in children treated for Burkitt lymphoma in Malawi. Pediatr. Blood Cancer 2009, 53, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Gelelete, C.B.; Pereira, S.H.; Azevedo, A.M.B.; Thiago, L.S.; Mundim, M.; Land, M.G.; Costa, E.S. Overweight as a prognostic factor in children with acute lymphoblastic leukemia. Obesity 2011, 19, 1908–1911. [Google Scholar] [CrossRef] [PubMed]
- Dickerman, J.D. The late effects of childhood cancer therapy. Pediatrics 2007, 119, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M.; Das, S.; Band, M.; Epstein, S.; Roberts, S. The effect of caloric restriction and glycemic load on measures of oxidative stress and antioxidants in humans: Results from the CALERIE Trial of Human Caloric Restriction. J. Nutr. Health Aging 2011, 15, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, M.; Stiakaki, E.; Dimitriou, H.; Malliaraki, N.; Notas, G.; Castanas, E.; Kalmanti, M. Cancer chemotherapy reduces plasma total antioxidant capacity in children with malignancies. Leuk. Res. 2005, 29, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Conklin, K.A. Chemotherapy-associated oxidative stress: Impact on chemotherapeutic effectiveness. Integr. Cancer Ther. 2004, 3, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Bindokas, V.P.; Jordan, J.; Lee, C.C.; Miller, R.J. Superoxide production in rat hippocampal neurons: Selective imaging with hydroethidine. J. Neurosci. 1996, 16, 1324–1336. [Google Scholar] [CrossRef] [PubMed]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2′,7′-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Kamencic, H.; Lyon, A.; Paterson, P.G.; Juurlink, B.H. Monochlorobimane fluorometric method to measure tissue glutathione. Anal. Biochem. 2000, 286, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Seki, T.; Maeda, H. Therapeutic strategies by modulating oxygen stress in cancer and inflammation. Adv. Drug Deliv. Rev. 2009, 61, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Beel, J.; Lillehei, K. A threshold concept for cancer therapy. Med. Hypotheses 2000, 55, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.E.; Rivera-Del Valle, N.; Chandra, J. Redox control of leukemia: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2013, 18, 1349–1383. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Shi, Y.; Le, G. Effect of dietary protein level and origin on the redox status in the digestive tract of mice. Int. J. Mol. Sci. 2008, 9, 464–475. [Google Scholar] [CrossRef]
- Rubio-Patiño, C.; Bossowski, J.P.; De Donatis, G.M.; Mondragón, L.; Villa, E.; Aira, L.E.; Chiche, J.; Mhaidly, R.; Lebeaupin, C.; Marchetti, S. Low-protein diet induces IRE1α-dependent anticancer immunosurveillance. Cell Metab. 2018, 27, 828–842. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Total n (%) | Age | p-Value | |||
|---|---|---|---|---|---|
| ≤48 Months | >48 Months | ||||
| Sex | Female | 13 (40.6%) | 8 (42.1%) | 5 (38.5%) | 1.0000 |
| Male | 19 (59.4%) | 11 (57.9%) | 8 (61.5%) | ||
| Diagnosis | Acute Myeloid Leukemia (AML) | 2 (6.3%) | 2 (10.5%) | 0 (0%) | 0.1238 |
| Anaplastic Large cell Lymphoma | 1 (3.1%) | 1 (5.3%) | 0 (0%) | ||
| High Risk pre-B Acute Lymphocytic (or lymphoblastic) Leukemia (ALL) | 8 (25%) | 5 (26.3%) | 3 (23.1%) | ||
| Relapse ALL | 3 (9.4%) | 0 (0%) | 3 (23.1%) | ||
| Standard Risk pre-B ALL | 14 (43.8%) | 8 (42.1%) | 6 (46.2%) | ||
| T-cell ALL | 3 (9.4%) | 3 (15.8%) | 0 (0%) | ||
| T-cell Lymphoma | 1 (3.1%) | 0 (0%) | 1 (7.7%) | ||
| Log (Ox Stress Measure) | Age | Visit | N | Mean | S.D. | Mixed Model Effect (p-Value) | ||
|---|---|---|---|---|---|---|---|---|
| Age | Visit | Age × Visit | ||||||
| Hydrogen Peroxide | ≤48 M | 1 | 19 | 6.33 | 1.60 | 0.007 | 0.464 | 0.798 |
| 2 | 19 | 6.19 | 1.56 | |||||
| 3 | 19 | 6.58 | 1.08 | |||||
| 4 | 16 | 6.59 | 1.23 | |||||
| >48 M | 1 | 12 | 7.08 | 0.98 | ||||
| 2 | 13 | 7.02 | 1.43 | |||||
| 3 | 13 | 7.33 | 1.35 | |||||
| 4 | 12 | 7.85 | 1.13 | |||||
| Superoxide | ≤48 M | 1 | 18 | 5.04 | 1.42 | 0.041 | 0.043 | 0.285 |
| 2 | 19 | 5.23 | 1.56 | |||||
| 3 | 19 | 5.94 | 1.33 | |||||
| 4 | 16 | 5.39 | 1.42 | |||||
| >48 M | 1 | 12 | 6.19 | 1.35 | ||||
| 2 | 13 | 5.49 | 1.87 | |||||
| 3 | 13 | 6.31 | 1.67 | |||||
| 4 | 13 | 6.61 | 0.79 | |||||
| Glutathione | ≤48 M | 1 | 18 | 3.36 | 1.07 | 0.016 | 0.333 | 0.355 |
| 2 | 19 | 3.36 | 1.19 | |||||
| 3 | 18 | 3.47 | 1.10 | |||||
| 4 | 16 | 3.29 | 1.22 | |||||
| >48 M | 1 | 13 | 3.83 | 0.74 | ||||
| 2 | 13 | 4.24 | 1.07 | |||||
| 3 | 13 | 4.26 | 1.22 | |||||
| 4 | 12 | 4.51 | 0.71 | |||||
| Visit 1 Mean (Range) | Visit 4 Mean (Range) | |||||
|---|---|---|---|---|---|---|
| ≤48 | >48 | All | ≤48 | >48 | All | |
| Protein (g) | 52 | 68 | 58 | 44 | 60 | 51 |
| (15–166) | (22–129) | (15–166) | (19–70) | (31–104) | (19–104) | |
| Fat (g) | 59 | 61 | 59 | 48 | 58 | 53 |
| (20–197) | (14–118) | (13–197) | (11–86) | (22–97) | (11–97) | |
| Carb (g) | 203 | 225 | 212 | 189 | 216 | 201 |
| (85–488) | (80–605) | (80–605) | (67–453) | (96–300) | (67–453) | |
| Kilocalories | 1523 | 1687 | 1590 | 1366 | 1560 | 1453 |
| (720–3182) | (672–3934) | (672–3934) | (444–2284) | (707–2326) | (444–2326) | |
| Association (p Value) | |||
|---|---|---|---|
| Dietary Components | Hydrogen Peroxide (DCF) | Superoxide (HE) | Glutathione (mBCI) |
| Kilocalories | 0.1904 | 0.4873 | 0.2428 |
| Riboflavin (mg) | 0.0314 * | 0.6332 | 0.4172 |
| Combined Carotene | 0.0845 | 0.1455 | 0.8918 |
| Iron (mg) | 0.2496 | 0.753 | 0.9812 |
| Vitamin A (IU) | 0.9154 | 0.2002 | 0.2876 |
| Vitamin B3 (mg) | 0.8738 | 0.2538 | 0.0401 * |
| Vitamin C (mg) | 0.3989 | 0.9296 | 0.4837 |
| Vitamin E (IU) | 0.1719 | 0.8186 | 0.7604 |
| Glutamic Acid (g) | 0.0444 * | 0.2042 | 0.0646 |
| Selenium (mcg) | 0.4216 | 0.076 | 0.5178 |
| Vitamin D (µg) | 0.841 | 0.7973 | 0.6851 |
| Natural Vitamin E (mg) | 0.4118 | 0.3922 | 0.5751 |
| Synthetic Vitamin E (mg) | 0.6466 | 0.7128 | 0.2702 |
| Total Protein (g) | 0.0534 * | 0.0226 * | 0.719 |
| Animal Protein (g) | 0.0001 ** | 0.0192 * | 0.6254 |
| Vegetable Protein (g) | 0.0001 ** | 0.0349 * | 0.0286 * |
| Association (p Value) | |||
|---|---|---|---|
| Dietary components | Hydrogen Peroxide (DCF) | Superoxide (HE) | Glutathione (mBCI) |
| Riboflavin (mg) | 0.0444 * | NS | NS |
| Combined Carotene (mcg) | 0.0820 | NS | NS |
| Total Protein (g) | 0.0002 ** | NS | NS |
| Selenium (mcg) | NS | 0.0760 | NS |
| Vegetable Protein (g) | NS | NS | 0.0286 * |
| Association (p Value) | |||
|---|---|---|---|
| Clinical Characteristics | Hydrogen Peroxide (DCF) | Superoxide (HE) | Glutathione (mBCI) |
| Height (cm) | 0.1084 | 0.0436 * | 0.9852 |
| Weight (kg) | 0.0497 * | 0.1119 | 0.9119 |
| BMI (kg/m2) | 0.6846 | 0.4628 | 0.2736 |
| WBC Count (K/µL) | 0.0765 | 0.5209 | 0.9234 |
| Hb (G/dL) | 0.4157 | 0.1066 | 0.9386 |
| Platelets (K/µL) | 0.004 ** | 0.1327 | 0.3905 |
| Any Infections (Y/N) | 0.1465 | 0.9638 | 0.7497 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raber, M.; Wu, J.; Donnella, H.; Knouse, P.; Pise, M.; Munsell, M.; Liu, D.; Chandra, J. Cellular Oxidative Stress in Pediatric Leukemia and Lymphoma Patients Undergoing Treatment Is Associated with Protein Consumption. Nutrients 2020, 12, 75. https://doi.org/10.3390/nu12010075
Raber M, Wu J, Donnella H, Knouse P, Pise M, Munsell M, Liu D, Chandra J. Cellular Oxidative Stress in Pediatric Leukemia and Lymphoma Patients Undergoing Treatment Is Associated with Protein Consumption. Nutrients. 2020; 12(1):75. https://doi.org/10.3390/nu12010075
Chicago/Turabian StyleRaber, Margaret, Jimin Wu, Hayley Donnella, Phillip Knouse, Mayurika Pise, Mark Munsell, Diane Liu, and Joya Chandra. 2020. "Cellular Oxidative Stress in Pediatric Leukemia and Lymphoma Patients Undergoing Treatment Is Associated with Protein Consumption" Nutrients 12, no. 1: 75. https://doi.org/10.3390/nu12010075
APA StyleRaber, M., Wu, J., Donnella, H., Knouse, P., Pise, M., Munsell, M., Liu, D., & Chandra, J. (2020). Cellular Oxidative Stress in Pediatric Leukemia and Lymphoma Patients Undergoing Treatment Is Associated with Protein Consumption. Nutrients, 12(1), 75. https://doi.org/10.3390/nu12010075