Effects of a Low-Fat Vegan Diet on Gut Microbiota in Overweight Individuals and Relationships with Body Weight, Body Composition, and Insulin Sensitivity. A Randomized Clinical Trial



Abstract
:1. Introduction
2. Materials and Methods
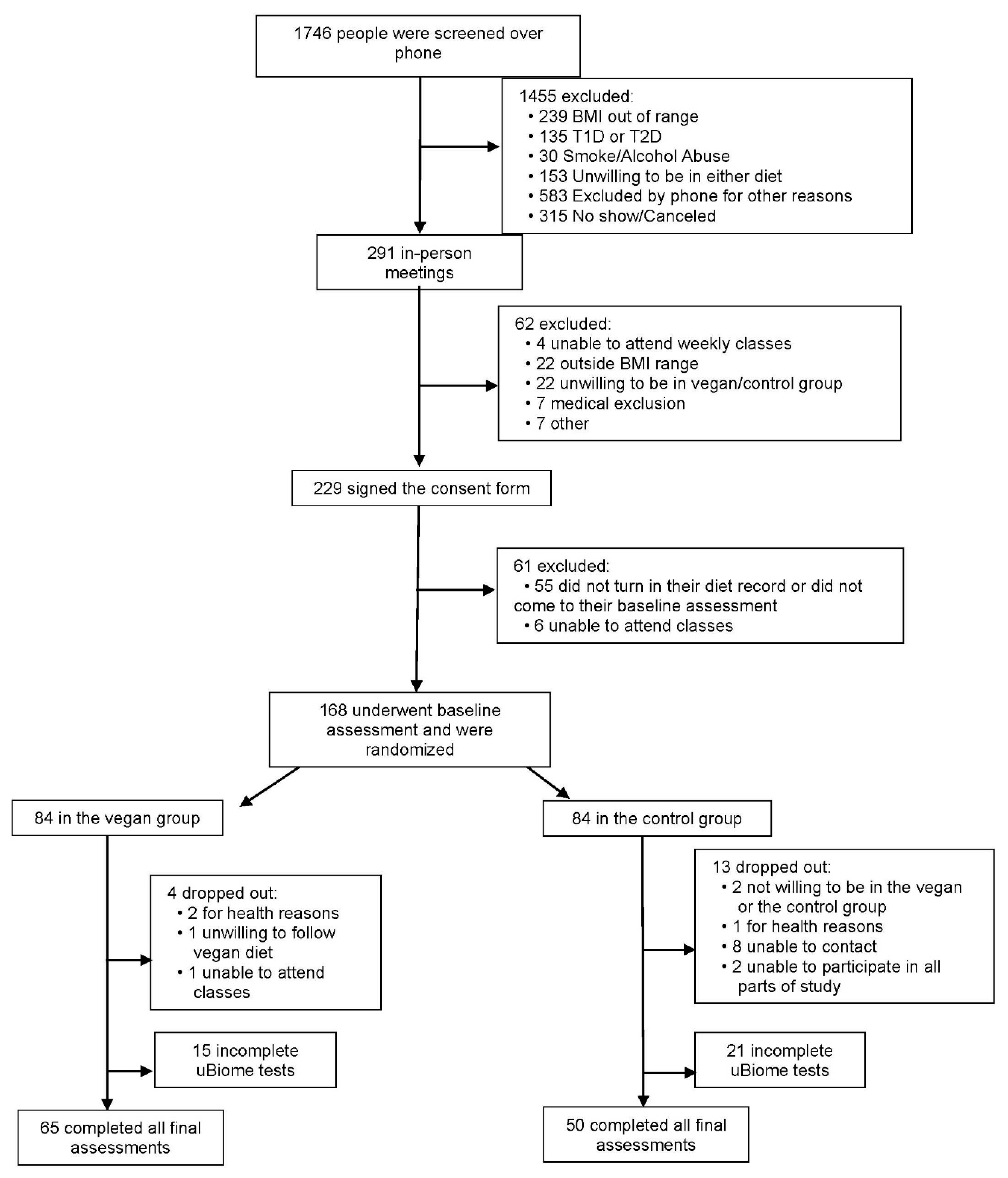
2.1. Participants and Methods
2.2. Study Design
2.3. Randomization and Study Groups
2.4. Dietary Intake and Physical Activity
2.5. Gut Microbiota Composition
2.6. Anthropometric and Metabolic Measurements
2.7. Statistical Analysis
3. Results
3.1. Characteristics of the Study Participants
3.2. Body Weight, Body Composition, and Insulin Sensitivity
3.3. Gut Microbiota Composition
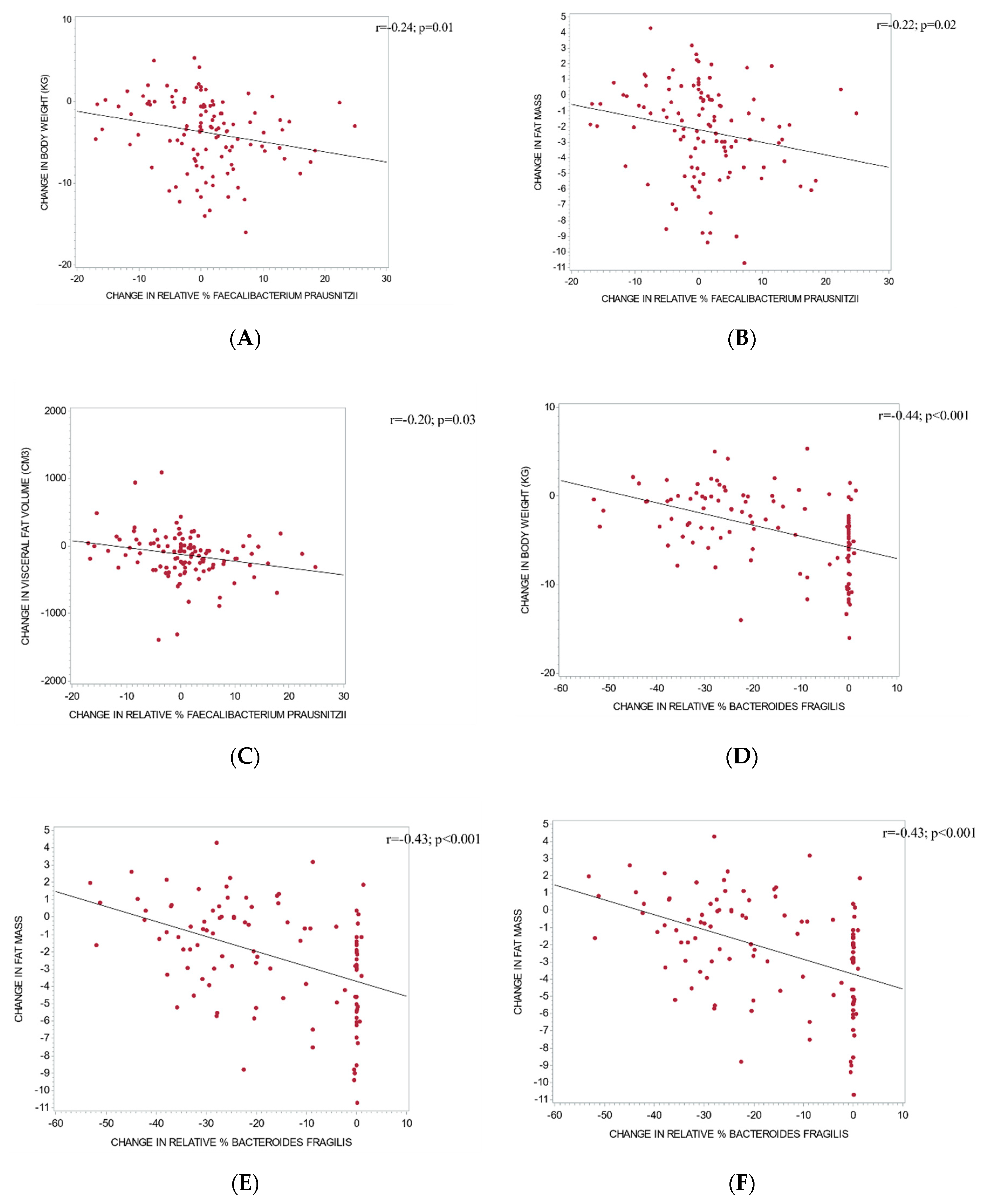
3.4. The Relationship between Changes in Gut Microbiota and Metabolic Outcomes
4. Discussion
4.1. Bacteroidetes and Diet
4.2. Firmicutes to Bacteroidetes Ratio, Diet, and Body Weight
4.3. Prevotella, Diet, and Body Weight
4.4. Faecalibacterium, Diet, and Body Weight
4.5. Bacteroides Fragilis, Diet, and Body Weight
4.6. Possible Mechanisms
4.7. Study Strengths and Limitations
4.8. Practical Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Khan, M.T.; Nieuwdorp, M.; Bäckhed, F. Microbial modulation of insulin sensitivity. Cell Metab. 2014, 20, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Ley, R.E. Obesity and the human microbiome. Curr. Opin. Gastroenterol. 2010, 26, 5–11. [Google Scholar] [CrossRef]
- Arora, T.; Sharma, R. Fermentation potential of the gut microbiome: Implications for energy homeostasis and weight management. Nutr. Rev. 2011, 69, 99–106. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Bäckhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Glick-Bauer, M.; Yeh, M.-C. The health advantage of a vegan diet: Exploring the gut microbiota connection. Nutrients 2014, 6, 4822–4838. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.-Y.; Huang, C.-C.; Hu, F.B.; Chavarro, J.E. Vegetarian Diets and Weight Reduction: A Meta-Analysis of Randomized Controlled Trials. J. Gen. Intern. Med. 2016, 31, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Tonstad, S.; Butler, T.; Yan, R.; Fraser, G.E. Type of Vegetarian Diet, Body Weight, and Prevalence of Type 2 Diabetes. Diabetes Care 2009, 32, 791–796. [Google Scholar] [CrossRef] [Green Version]
- Kahleova, H.; Petersen, K.F.; Shulman, G.I.; Alwarith, J.; Rembert, E.; Tura, A.; Hill, M.; Holubkov, R.; Barnard, N.D. Effect of a low-fat vegan diet on body weight, insulin sensitivity, postprandial metabolism, and intramyocellular and hepatocellular lipids in overweight adults: A randomized clinical trial. JAMA Network Open 2020. accepted for publication. [Google Scholar]
- Schakel, S.F.; Sievert, Y.A.; Buzzard, I.M. Sources of data for developing and maintaining a nutrient database. J. Am. Diet. Assoc. 1988, 88, 1268–1271. [Google Scholar] [PubMed]
- Hagströmer, M.; Oja, P.; Sjöström, M. The International Physical Activity Questionnaire (IPAQ): A study of concurrent and construct validity. Public Health Nutr. 2006, 9, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Almonacid, D.E.; Kraal, L.; Ossandon, F.J.; Budovskaya, Y.V.; Cardenas, J.P.; Bik, E.M.; Goddard, A.D.; Richman, J.; Apte, Z.S. 16S rRNA gene sequencing and healthy reference ranges for 28 clinically relevant microbial taxa from the human gut microbiome. PLoS ONE 2017, 12, e0176555. [Google Scholar] [CrossRef] [Green Version]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and fast clustering method for amplicon-based studies. PeerJ 2014, 2, e593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids. Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McCoy, C.O.; Iv, F.A.M. Abundance-weighted phylogenetic diversity measures distinguish microbial community states and are robust to sampling depth. PeerJ 2013, 1, e157. [Google Scholar] [CrossRef] [Green Version]
- Tura, A.; Chemello, G.; Szendroedi, J.; Göbl, C.; Færch, K.; Vrbíková, J.; Pacini, G.; Ferrannini, E.; Roden, M. Prediction of clamp-derived insulin sensitivity from the oral glucose insulin sensitivity index. Diabetologia 2018, 61, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Matijašić, B.B.; Obermajer, T.; Lipoglavšek, L.; Grabnar, I.; Avguštin, G.; Rogelj, I. Association of dietary type with fecal microbiota in vegetarians and omnivores in Slovenia. Eur. J. Nutr. 2014, 53, 1051–1064. [Google Scholar] [CrossRef]
- Liszt, K.; Zwielehner, J.; Handschur, M.; Hippe, B.; Thaler, R.; Haslberger, A.G. Characterization of bacteria, clostridia and Bacteroides in faeces of vegetarians using qPCR and PCR-DGGE fingerprinting. Ann. Nutr. Metab. 2009, 54, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Li, X.H.; Chen, W.N. Similarities and differences in gut microbiome composition correlate with dietary patterns of Indian and Chinese adults. AMB Express 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, I.; Kim, J.; Duffy, P.R.; Schlegel, V.L.; Walter, J. Resistant Starches Types 2 and 4 Have Differential Effects on the Composition of the Fecal Microbiota in Human Subjects. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdam, F.J.; Fuentes, S.; de Jonge, C.; Zoetendal, E.G.; Erbil, R.; Greve, J.W.; Buurman, W.A.; de Vos, W.M.; Rensen, S.S. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity: Obese Gut Microbiota and Inflammation. Obesity 2013, 21, E607–E615. [Google Scholar] [CrossRef] [PubMed]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17. [Google Scholar] [CrossRef] [Green Version]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity (Silver Spring) 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associated with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef] [Green Version]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans123. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-S.; Hwang, S.-S.; Park, E.-J.; Bae, J.-W. Strict vegetarian diet improves the risk factors associated with metabolic diseases by modulating gut microbiota and reducing intestinal inflammation. Environ. Microbiol. Rep. 2013, 5, 765–775. [Google Scholar] [CrossRef]
- Gong, J.; Yang, C. Advances in the methods for studying gut microbiota and their relevance to the research of dietary fiber functions. Food Res. Int. 2012, 48, 916–929. [Google Scholar] [CrossRef]
- Christensen, L.; Roager, H.M.; Astrup, A.; Hjorth, M.F. Microbial enterotypes in personalized nutrition and obesity management. Am. J. Clin. Nutr. 2018, 108, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Ferrocino, I.; Di Cagno, R.; De Angelis, M.; Turroni, S.; Vannini, L.; Bancalari, E.; Rantsiou, K.; Cardinali, G.; Neviani, E.; Cocolin, L. Fecal Microbiota in Healthy Subjects Following Omnivore, Vegetarian and Vegan Diets: Culturable Populations and rRNA DGGE Profiling. PLoS ONE 2015, 10, e0128669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, J.; Carbonero, F.; Zoetendal, E.G.; DeLany, J.P.; Wang, M.; Newton, K.; Gaskins, H.R.; O’Keefe, S.J.D. Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am. J. Clin. Nutr. 2013, 98, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abell, G.C.J.; Cooke, C.M.; Bennett, C.N.; Conlon, M.A.; McOrist, A.L. Phylotypes related to Ruminococcus bromii are abundant in the large bowel of humans and increase in response to a diet high in resistant starch. FEMS Microbiol. Ecol. 2008, 66, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Candela, M.; Biagi, E.; Soverini, M.; Consolandi, C.; Quercia, S.; Severgnini, M.; Peano, C.; Turroni, S.; Rampelli, S.; Pozzilli, P.; et al. Modulation of gut microbiota dysbioses in type 2 diabetic patients by macrobiotic Ma-Pi 2 diet. Br. J. Nutr. 2016, 116, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Furet, J.-P.; Kong, L.-C.; Tap, J.; Poitou, C.; Basdevant, A.; Bouillot, J.-L.; Mariat, D.; Corthier, G.; Doré, J.; Henegar, C.; et al. Differential adaptation of human gut microbiota to bariatric surgery-induced weight loss: Links with metabolic and low-grade inflammation markers. Diabetes 2010, 59, 3049–3057. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lian, F.; Zhao, L.; Zhao, Y.; Chen, X.; Zhang, X.; Guo, Y.; Zhang, C.; Zhou, Q.; Xue, Z.; et al. Structural modulation of gut microbiota during alleviation of type 2 diabetes with a Chinese herbal formula. ISME J. 2015, 9, 552–562. [Google Scholar] [CrossRef]
- LEE, Y.-K. Effects of Diet on Gut Microbiota Profile and the Implications for Health and Disease. Biosci. Microbiota Food Health 2013, 32, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Saarinen, M.T.; Lahtinen, S.J.; Sørensen, J.F.; Tiihonen, K.; Ouwehand, A.C.; Rautonen, N.; Morgan, A. Treatment of bran containing bread by baking enzymes; effect on the growth of probiotic bacteria on soluble dietary fiber extract in vitro. Biosci. Biotechnol. Biochem. 2012, 76, 1135–1139. [Google Scholar] [CrossRef] [Green Version]
- Rios-Covian, D.; Sánchez, B.; Salazar, N.; Martínez, N.; Redruello, B.; Gueimonde, M.; de Los Reyes-Gavilán, C.G. Different metabolic features of Bacteroides fragilis growing in the presence of glucose and exopolysaccharides of bifidobacteria. Front. Microbiol. 2015, 6, 825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Björkman, A.; Cai, K.; Liu, G.; Wang, C.; Li, Y.; Xia, H.; Sun, L.; Kristiansen, K.; Wang, J.; et al. Impact of a 3-Months Vegetarian Diet on the Gut Microbiota and Immune Repertoire. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Wang, D.; Ren, H.; Cai, K.; Chen, P.; Fang, C.; Shi, Z.; Zhang, P.; Wang, J.; Yang, H.; et al. Effect of Caloric Restriction on BMI, Gut Microbiota, and Blood Amino Acid Levels in Non-Obese Adults. Nutrients 2020, 12, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomova, A.; Bukovsky, I.; Rembert, E.; Yonas, W.; Alwarith, J.; Barnard, N.D.; Kahleova, H. The Effects of Vegetarian and Vegan Diets on Gut Microbiota. Front. Nutr. 2019, 6, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnard, N.D.; Gloede, L.; Cohen, J.; Jenkins, D.J.A.; Turner-McGrievy, G.; Green, A.A.; Ferdowsian, H. A low-fat vegan diet elicits greater macronutrient changes, but is comparable in adherence and acceptability, compared with a more conventional diabetes diet among individuals with type 2 diabetes. J. Am. Diet. Assoc. 2009, 109, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Spiegelman, D.; Rimm, E.B.; Rosner, B.A.; Stampfer, M.J.; Barnett, J.B.; Chavarro, J.E.; Rood, J.C.; Harnack, L.J.; Sampson, L.K.; et al. Relative Validity of Nutrient Intakes Assessed by Questionnaire, 24-Hour Recalls, and Diet Records Compared With Urinary Recovery and Plasma Concentration Biomarkers: Findings for Women. Am. J. Epidemiol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, R.; Mainali, R.; Ahmadi, S.; Wang, S.; Singh, R.; Kavanagh, K.; Kitzman, D.W.; Kushugulova, A.; Marotta, F.; Yadav, H. Gut microbiome and aging: Physiological and mechanistic insights. Nutr. Healthy Aging 2018, 4, 267–285. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Park, K. Adherence to a Vegetarian Diet and Diabetes Risk: A Systematic Review and Meta-Analysis of Observational Studies. Nutrients 2017, 9, 603. [Google Scholar] [CrossRef] [Green Version]
- Viguiliouk, E.; Kendall, C.W.; Kahleová, H.; Rahelić, D.; Salas-Salvadó, J.; Choo, V.L.; Mejia, S.B.; Stewart, S.E.; Leiter, L.A.; Jenkins, D.J.; et al. Effect of vegetarian dietary patterns on cardiometabolic risk factors in diabetes: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2018, 38, 1133–1145. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | Vegan Group (n = 84) | Control Group (n = 84) | Statistic |
|---|---|---|---|
| Age (years) | 52.9 ± 11.7 | 57.5 ± 10.2 | p = 0.01 |
| Sex (number, %) | |||
| Male | 15 (17.9) | 10 (11.9) | p = 0.28 |
| Female | 69 (82.1) | 74 (88.1) | |
| BMI (kg/m2) | 32.6 ± 3.7 | 33.6 ± 3.8 | p = 0.10 |
| Race, (number, %) | |||
| White | 42 (50.0) | 41 (48.8) | p = 1.00 |
| Black | 40 (47.6) | 39 (46.4) | |
| Asian, Pacific Islander | 1 (1.2) | 2 (2.4) | |
| Did not disclose | 1 (1.2) | 2 (2.4) | |
| Ethnicity, (number, %) | |||
| Non-Hispanic | 64 (76.2) | 69 (82.1) | p = 0.74 |
| Hispanic | 5 (5.9) | 4 (4.8) | |
| Did not disclose | 15 (17.9) | 11 (13.1) | |
| Marital status | |||
| Not married | 44 (52.4) | 42 (50.0) | p = 0.27 |
| Married | 39 (46.4) | 35 (41.7) | |
| Did not disclose | 1 (1.2) | 7 (8.3) | |
| Education | |||
| High school | 8 (9.5) | 11 (13.1) | p = 0.20 |
| Associates | 0 | 1 (1.2) | |
| College | 23 (27.4) | 29 (34.5) | |
| Graduate degree | 53 (63.1) | 43 (51.2) | |
| Occupation | |||
| Service occupation | 22 (26.2) | 14 (16.7) | p = 0.51 |
| Technical, sales, administrative | 19 (22.6) | 22 (26.2) | |
| Professional or managerial | 23 (27.4) | 23 (27.4) | |
| Retired | 12 (14.3) | 18 (21.4) | |
| Other | 8 (9.5) | 7 (8.3) | |
| Medications (%) | |||
| Lipid-lowering therapy | 17 (20.2) | 18 (21.4) | p = 0.85 |
| Antihypertensive therapy | 21 (25.0) | 25 (29.8) | p = 0.49 |
| Thyroid medications | 10 (11.9) | 7 (8.3) | p = 0.44 |
| Control Group | Vegan Group | Treatment Effect | p Value a | p Value b | |||
|---|---|---|---|---|---|---|---|
| Baseline | Week 16 | Baseline | Week 16 | ||||
| Total Physical activity (metabolic equivalents) | 2840 (2064–3616) | 2107 (1555–2658) * | 2982 (1696–4267) | 2000 (1394–2606) | −248 (−1549 to +1053) | 0.71 | 0.87 |
| Dietary Intake | |||||||
| Caloric Intake (kcal/day) | 1726 (1606–1847) | 1692 (1562–1821) | 1827 (1689–1965) | 1294 (1212–1376) *** | −498 (−696 to −300) | <0.001 | <0.001 |
| Total Fat (g/day) | 72.2 (66.1–78.3) | 71.5 (63.9–79.0) | 74.1 (67.9–80.4) | 24.3 (21.7–26.9) *** | −49.1 (−58.5 to −39.8) | <0.001 | <0.001 |
| Total Carbohydrate (g/day) | 204 (187–221) | 196 (178–214) | 222 (202–241) | 236 (219–254) | +22.6 (−6.5 to +51.7) | 0.13 | 0.17 |
| Total Protein (g/day) | 67.2 (62.2–72.1) | 68.9 (63.1–74.6) | 69.0 (63.9–74.1) | 42.9 (40.0–45.7) *** | −27.9 (−36.0 to −19.7) | <0.001 | <0.001 |
| Animal Protein (g/day) | 37.5 (32.5–42.4) | 39.3 (33.4–45.3) | 38.5 (33.7–43.2) | 1.2 (0.5–1.9) *** | −39.1 (−46.4 to −31.9) | <0.001 | <0.001 |
| Veg Protein (g/day) | 29.7 (26.8–32.6) | 29.5 (26.0–33.0) | 30.6 (27.5–33.7) | 41.7 (39.0–44.3) *** | +11.3 (+6.6 to +16.0) | <0.001 | <0.001 |
| Cholesterol (mg/day) | 227 (190–263) | 243 (201–285) | 227 (196–258) | 5.1 (3.3–6.9) *** | −238 (−285 to −191) | <0.001 | <0.001 |
| Total SFA (g/day) | 21.7 (19.3–24.0) | 21.9 (18.9–24.9) | 23.3 (20.4–26.1) | 4.8 (4.2–5.4) *** | −18.7 (−22.7 to −14.8) | <0.001 | <0.001 |
| Total MUFA (g/day) | 26.7 (24.1–29.3) | 25.8 (23.0–28.5) | 26.4 (24.2–28.6) | 7.9 (7.0–8.7) *** | −17.6 (−21.2 to −14.0) | <0.001 | <0.001 |
| Total PUFA (g/day) | 17.9 (16.4–19.5) | 17.8 (15.8–19.9) | 18.4 (16.7–20.2) | 9.0 (7.9–10.2) *** | −9.3 (−12.1 to −6.6) | <0.001 | <0.001 |
| Total Dietary Fiber (g/day) | 23.1 (21.0–25.2) | 23.1 (20.7–25.4) | 24.0 (21.6–26.5) | 33.2 (30.8–35.6) *** | +9.2 (+5.6 to +12.8) | <0.001 | <0.001 |
| Soluble Fiber (g/day) | 6.0 (5.5–6.5) | 6.6 (6.0–7.2) * | 6.9 (6.2–7.5) | 8.5 (7.8–9.2) *** | +1.0 (+0.1 to +2.0) | 0.03 | 0.01 |
| Insoluble Fiber (g/day) | 17.0 (15.3–18.8) | 16.4 (14.5–18.2) | 17.1 (15.1–19.0) | 24.6 (22.7–26.4) *** | +8.2 (+5.2 to +11.1) | <0.001 | <0.001 |
| Anthropo-Metabolic Outcomes | |||||||
| Weight (kg) | 93.4 (90.1–96.7) | 92.9 (89.6–96.3) | 92.9 (89.7–96.1) | 86.5 (83.5–89.5) *** | −5.9 (−7.0 to −4.9) | <0.001 | <0.001 |
| BMI (kg/m2) | 33.6 (32.6–34.5) | 33.4 (32.4–34.4) | 32.6 (31.8–33.5) | 30.5 (29.6–31.3) *** | −2.0 (−2.4 to −1.6) | <0.001 | <0.001 |
| Fat Mass (kg) | 42.0 (39.7–44.2) | 41.7 (39.4–44.1) | 39.8 (37.7–42.0) | 35.7 (33.6–37.9) *** | −3.9 (−4.6 to −3.1) | <0.001 | <0.001 |
| VAT Volume (cm3) | 1590 (1365–1814) | 1589 (1360–1818) | 1511 (1291–1732) | 1271 (1084–1457) *** | −240 (−345 to −135) | <0.001 | <0.001 |
| PREDIM (mg/min/kg) | 4.4 (4.0–4.8) | 4.2 (3.8–4.6) | 4.0 (3.7–4.3) | 4.8 (4.4–5.1) *** | +0.83 (+0.48 to +1.2) | <0.001 | <0.001 |
| Gut Microbiota Composition | |||||||
| Diversity | 1.6 (1.5–1.7) | 1.8 (1.7–1.9) *** | 1.7 (1.6–1.8) | 1.7 (1.6–1.8) | −0.20 (−0.34 to −0.06) | 0.0043 | 0.003 |
| Firmicutes | 60,997 (41,121–80,874) | 60,599 (51,612–69,586) | 52,038 (41,221–62,856) | 64,734 (53,368–76,100) | +13,094 (−11,808 to +37,996) | 0.30 | 0.42 |
| Firmicutes % | 52.2 (48.7–55.7) | 49.8 (46.6–52.9) | 55.4 (51.7–59.1) | 54.6 (51.4–57.9) | +1.7 (−2.7 to +6.1) | 0.45 | 0.72 |
| Bacteroidetes | 41,690 (33,401–49,980) | 53,871 (41,691–66,051) | 31,944 (24,747–39,141) | 42,855 (34,088–51,622) * | −1270 (−17,950 to +15,409) | 0.88 | 1.00 |
| Bacteroidetes % | 37.6 (34.1–41.1) | 38.3 (34.9–41.7) | 31.6 (28.4–34.8) | 35.1 (31.4–38.8) | +2.8 (−2.0 to +7.5) | 0.25 | 0.06 |
| Enterobacteriaceae | 600 (−109–1308) | 709 (−153–1570) | 293 (135–451) | 683 (−79.0–1444) | +280.6 (−545 to +1106) | 0.50 | 0.44 |
| Enterobacteriaceae % | 0.47 (0.02–0.93) | 0.66 (0.02–1.3) | 0.73 (0.03–1.4) | 0.47 (0.02–0.92) | −0.44 (−1.3 to +0.45) | 0.33 | 0.25 |
| Firmicutes:Bacteroidetes ratio | 3.0 (0.38–5.6) | 2.0 (0.92–3.1) | 2.4 (1.6–3.1) | 2.3 (1.7–2.9) | +0.90 (−0.76 to +2.6) | 0.28 | 0.26 |
| Butyrate producing bacteria | 41,781 (34,410–49,152) | 34358 (27,310–41,407) * | 37,169 (29,540–44,799) | 38,143 (30,371–45,915) | +8396 (−4458 to +21249) | 0.20 | 0.32 |
| Butyrate producing bacteria % | 22.0 (19.5–24.5) | 19.4 (16.8–21.9) | 23.4 (21.5–25.2) | 21.2 (19.2–23.2) | +0.48 (−4.0 to +5.0) | 0.83 | 0.74 |
| Prevotella | 240 (104–376) | 554 (49.4–1059) | 224 (111–336) | 402 (169–636) | −135 (−711 to +439) | 0.64 | 0.61 |
| Prevotella % | 0.35 (0.09–0.60) | 1.15 (0.08–2.2) | 0.69 (0.01–1.4) | 0.84 (0.02–1.7) | −0.65 (−2.1 to +0.8) | 0.39 | 0.56 |
| Akkermansia | 1089 (495–1684) | 5073 (66.0–10080) | 2256 (640–3871) | 3215 (1873–4557) | −3024 (−8211 to +2163) | 0.25 | 0.26 |
| Akkermansia % | 1.4 (0.74–2.1) | 2.4 (0.85–4.0) | 2.1 (1.0–3.1) | 2.3 (1.5–3.2) | −0.74 (−2.6 to +1.1) | 0.43 | 0.40 |
| Faecalibacterium prausnitzii | 6935 (4905–8966) | 7142 (4502–9783) | 6304 (4127–8481) | 12405 (8417–16394) * | +5895 (+506 to +11283) | 0.03 | 0.17 |
| Faecalibacterium prausnitzii % | 7.2 (5.2–9.2) | 5.3 (3.8–6.9) | 5.5 (4.3–6.8) | 8.8 (7.0–10.6) *** | +5.1 (+2.4 to +7.9) | 0.0003 | 0.002 |
| Bacteroides fragilis | 31212 (23918–38505) | 602 (−275–1478) *** | 10641 (5513–15769) | 524 (−19.0–1067) *** | +20493 (+11790 to +29195) | <.001 | <.001 |
| Bacteroides fragilis % | 27.1 (23.6–30.6) | 0.3 (0.0–0.60) *** | 8.3 (5.1–11.5) | 0.40 (0.10–0.70) *** | +18.9 (+14.2 to +23.7) | <.001 | <.001 |
| Clostridium | 844 (529–1160) | 956 (705–1206) | 631 (432–829) | 880 (694–1066) | +138 (−245 to +521) | 0.48 | 0.62 |
| Clostridium % | 0.72 (0.54–0.90) | 0.74 (0.58–0.90) | 0.68 (0.55–0.81) | 0.76 (0.63–0.88) | +0.05 (−0.17 to +0.27) | 0.63 | 0.71 |
| Methanobrevibacter | 71.4 (22.9–120) | 506 (156–856) * | 299 (37.1–560) | 826 (−2.8–1655) | +93.1 (−817 to +1003) | 0.84 | 0.51 |
| Methanobrevibacter % | 0.09 (0.03–0.14) | 0.35 (0.14–0.57) ** | 0.23 (0.07–0.39) | 0.57 (0.18–0.96) | +0.07 (−0.35 to +0.49) | 0.74 | 0.25 |
| Eubacterium | 0.42 (0.05–0.79) | 2.0 (0.37–3.7) * | 1.0 (0.08–2.0) | 0.9 (0.03–1.8) | −1.7 (−3.5 to +0.09) | 0.06 | 0.05 |
| Eubacterium % | 0.001 (0.00002–0.001) | 0.002 (0.0002–0.003) | 0.0009 (0.0001–0.002) | 0.002 (−0.0005–0.004) | −0.0003 (−0.002 to +0.002) | 0.77 | 0.88 |
| Bifidobacterium | 1254 (412–2096) | 1712 (764–2659) | 1278 (738–1818) | 2313 (1286–3339) * | +577 (−700 to +1854) | 0.37 | 0.89 |
| Bifidobacterium % | 1.4 (0.48–2.3) | 1.8 (0.35–3.3) | 1.3 (0.79–1.8) | 1.7 (1.1–2.3) | −0.05 (−0.91 to +0.81) | 0.91 | 0.64 |
| Proteobacteria | 5278 (3672–6883) | 5195 (3305–7086) | 3561 (2503–4619) | 4165 (2798–5533) | +686 (−1622 to +2994) | 0.56 | 0.78 |
| Proteobacteria % | 4.9 (3.7–6.0) | 4.0 (3.0–5.1) | 4.4 (3.2–5.5) | 3.1 (2.4–3.9) * | −0.40 (−2.0 to +1.2) | 0.61 | 0.58 |
| Actinobacteria | 2736 (1395–4077) | 2247 (1261–3233) | 2459 (1716–3203) | 2729 (1658–3800) | +758 (−1071 to +2588) | 0.41 | 0.85 |
| Actinobacteria % | 3.1 (1.9–4.4) | 2.9 (1.2–4.7) | 2.6 (2.0–3.1) | 2.4 (1.6–3.2) | +0.05 (−1.4 to +1.5) | 0.94 | 0.91 |
| Ruminococcaceae | 15,842 (11,727–19,956) | 15775 (12,261–19,289) | 18,807 (14,337–23,277) | 22,265 (17,204–27,326) | +3525 (−4962 to +12011) | 0.41 | 0.96 |
| Ruminococcaceae % | 15.4 (13.0–17.8) | 12.9 (10.5–15.2) | 19.0 (16.7–21.3) | 18.5 (16.2–20.8) | +2.1 (−1.4 to +5.5) | 0.24 | 0.66 |
| Lachnospiraceae | 26,182 (20,879–31,485) | 18,224 (12,241–24,207) ** | 22,871 (18,631–27,111) | 15,881 (11,212–20,550) * | +968 (−7106 to +9042) | 0.81 | 0.79 |
| Lachnospiraceae % | 24.4 (22.1–26.7) | 15.4 (11.3–19.4) *** | 24.8 (22.5–27.1) | 16.9 (13.6–20.2) *** | +1.2 (−4.6 to +7.0) | 0.69 | 0.91 |
| Roseburia | 5993 (4634–7351) | 5639 (4367–6912) | 5821 (4255–7387) | 6649 (4953–8345) | +1181 (−1469 to +3831) | 0.38 | 0.34 |
| Roseburia % | 5.8 (4.7–6.9) | 5.1 (4.1–6.1) | 6.4 (5.3–7.5) | 6.0 (4.8–7.2) | +0.28 (−1.6 to +2.2) | 0.77 | 0.90 |
| Anaerostipes | 1502 (1020–1985) | 2061 (1496–2625) | 1481 (1103–1858) | 2203 (1614–2792) * | +164 (−772 to +1100) | 0.73 | 0.45 |
| Anaerostipes % | 1.4 (1.1–1.8) | 1.5 (1.2–1.8) | 1.6 (1.2–1.9) | 1.8 (1.4–2.1) | +0.10 (−0.48 to +0.69) | 0.72 | 0.33 |
| Megasphaera | 324 (−264–912) | 334 (−270–938) | 60.7 (19.5–102) | 123 (−9.2–255) | +51.8 (−58.0 to +162) | 0.35 | 0.80 |
| Megasphaera % | 0.72 (−0.67–2.1) | 0.62 (−0.42–1.7) | 0.12 (0.00–0.25) | 0.19 (−0.05–0.42) | +0.16 (−0.24 to +0.56) | 0.42 | 0.67 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahleova, H.; Rembert, E.; Alwarith, J.; Yonas, W.N.; Tura, A.; Holubkov, R.; Agnello, M.; Chutkan, R.; Barnard, N.D. Effects of a Low-Fat Vegan Diet on Gut Microbiota in Overweight Individuals and Relationships with Body Weight, Body Composition, and Insulin Sensitivity. A Randomized Clinical Trial. Nutrients 2020, 12, 2917. https://doi.org/10.3390/nu12102917
Kahleova H, Rembert E, Alwarith J, Yonas WN, Tura A, Holubkov R, Agnello M, Chutkan R, Barnard ND. Effects of a Low-Fat Vegan Diet on Gut Microbiota in Overweight Individuals and Relationships with Body Weight, Body Composition, and Insulin Sensitivity. A Randomized Clinical Trial. Nutrients. 2020; 12(10):2917. https://doi.org/10.3390/nu12102917
Chicago/Turabian StyleKahleova, Hana, Emilie Rembert, Jihad Alwarith, Willy N. Yonas, Andrea Tura, Richard Holubkov, Melissa Agnello, Robynne Chutkan, and Neal D. Barnard. 2020. "Effects of a Low-Fat Vegan Diet on Gut Microbiota in Overweight Individuals and Relationships with Body Weight, Body Composition, and Insulin Sensitivity. A Randomized Clinical Trial" Nutrients 12, no. 10: 2917. https://doi.org/10.3390/nu12102917
APA StyleKahleova, H., Rembert, E., Alwarith, J., Yonas, W. N., Tura, A., Holubkov, R., Agnello, M., Chutkan, R., & Barnard, N. D. (2020). Effects of a Low-Fat Vegan Diet on Gut Microbiota in Overweight Individuals and Relationships with Body Weight, Body Composition, and Insulin Sensitivity. A Randomized Clinical Trial. Nutrients, 12(10), 2917. https://doi.org/10.3390/nu12102917