PROVIT: Supplementary Probiotic Treatment and Vitamin B7 in Depression—A Randomized Controlled Trial

 , , , , and add
Show full author list
, , , , and add
Show full author list
Abstract
:1. Introduction
- Experience significantly higher improvement in psychiatric symptoms than individuals treated with placebo after one month;
- Show significant changes in intestinal barrier function (measured by zonulin) in comparison to treatment with placebo after one month;
- Have significant changes in microbiome analysis (alpha and beta diversity, global differential abundance, Piphillin-Analysis) in comparison to individuals with placebo treatment after one week as well as after one month.
2. Materials and Methods
2.1. Study Medication
2.2. Demographics and Scales of PROVIT
2.3. Statistics of Clinical Data
2.4. Zonulin Analysis
2.5. Microbiome Analysis
2.6. Bioinformatics
2.7. Statistical Analysis and Visualization of Microbiome Data
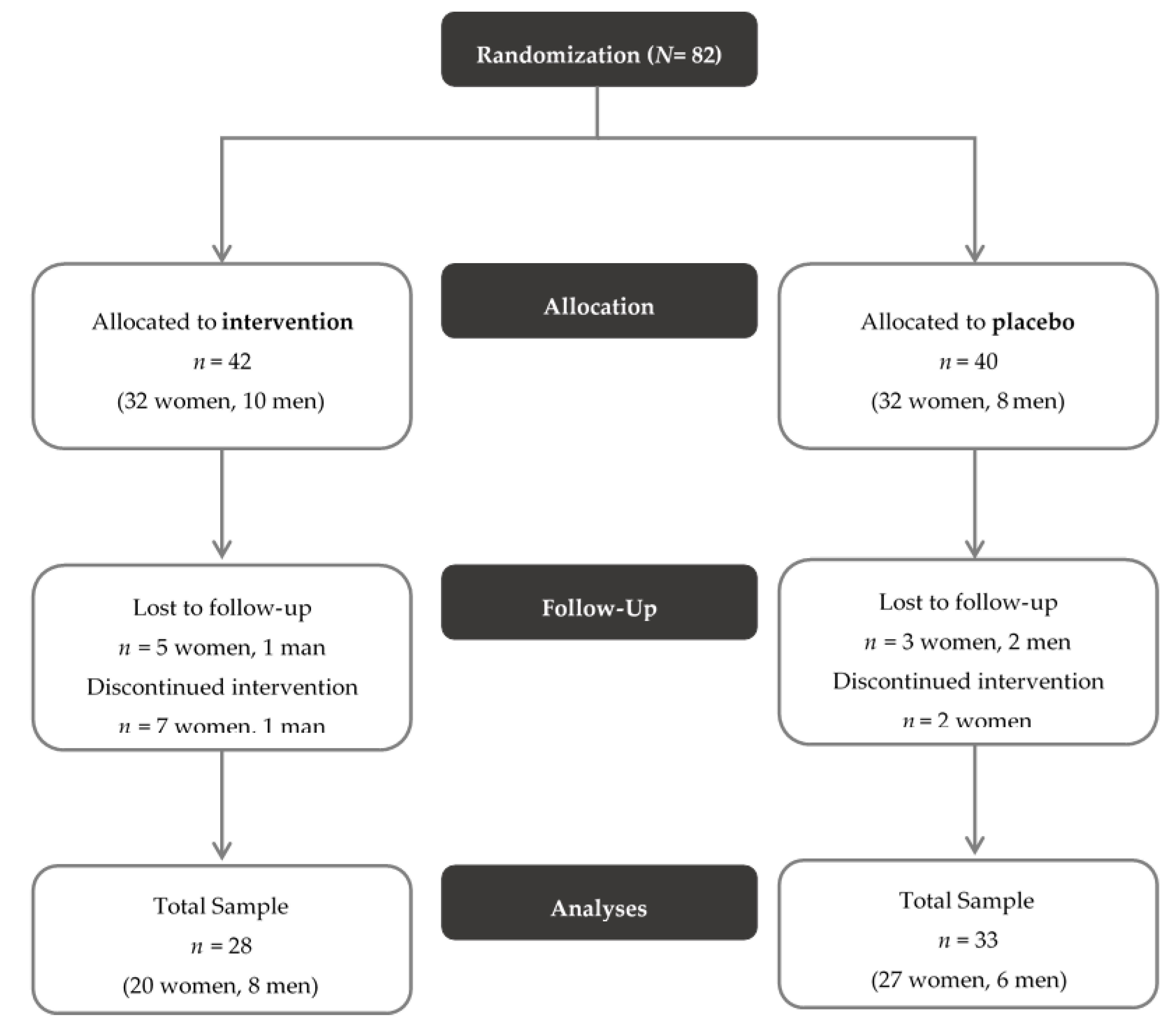
3. Results
3.1. Clinical Parameters
3.2. Anthropometric Data
3.3. Zonulin
3.4. Microbiome Analysis
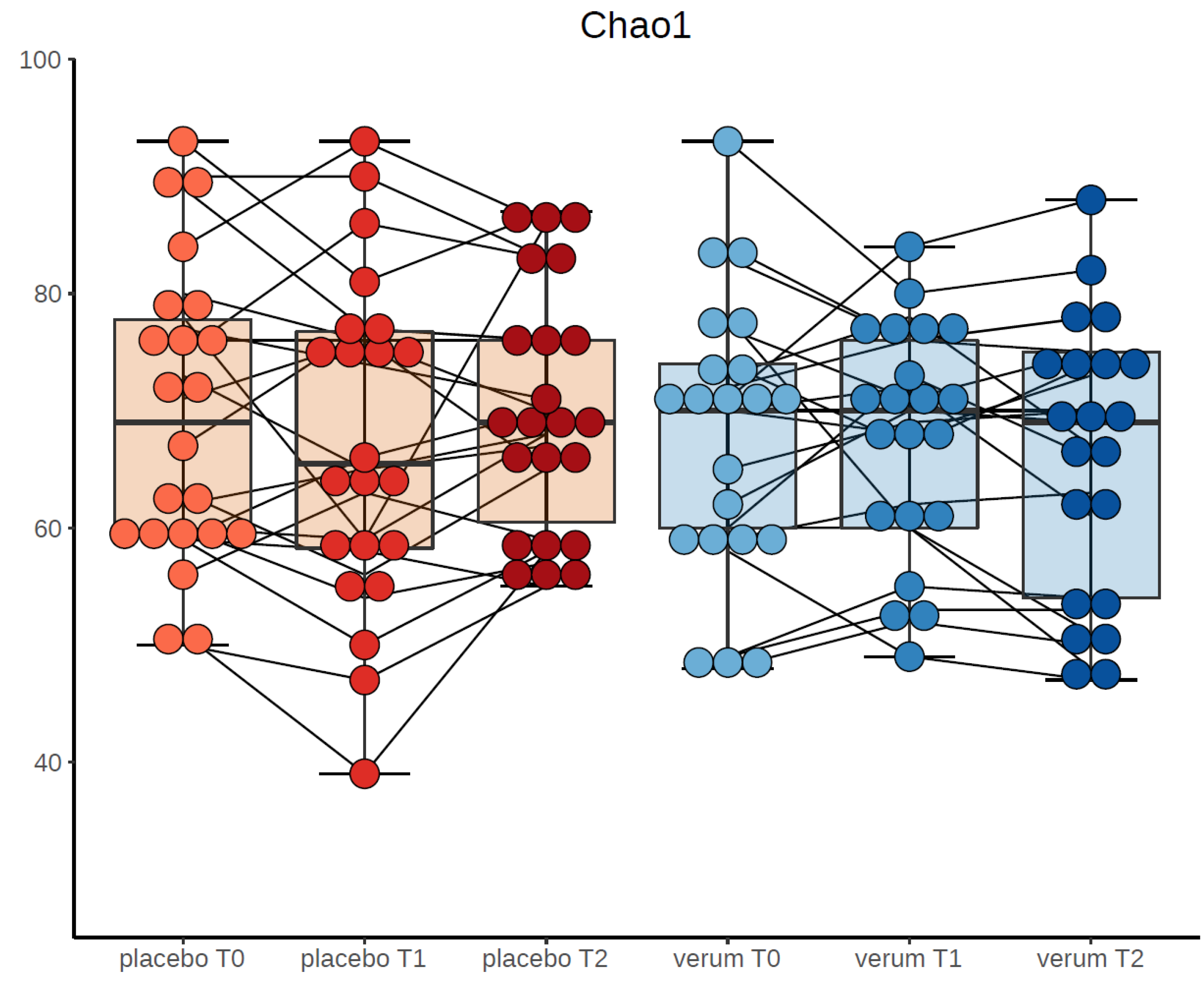
3.4.1. Alpha-Diversity
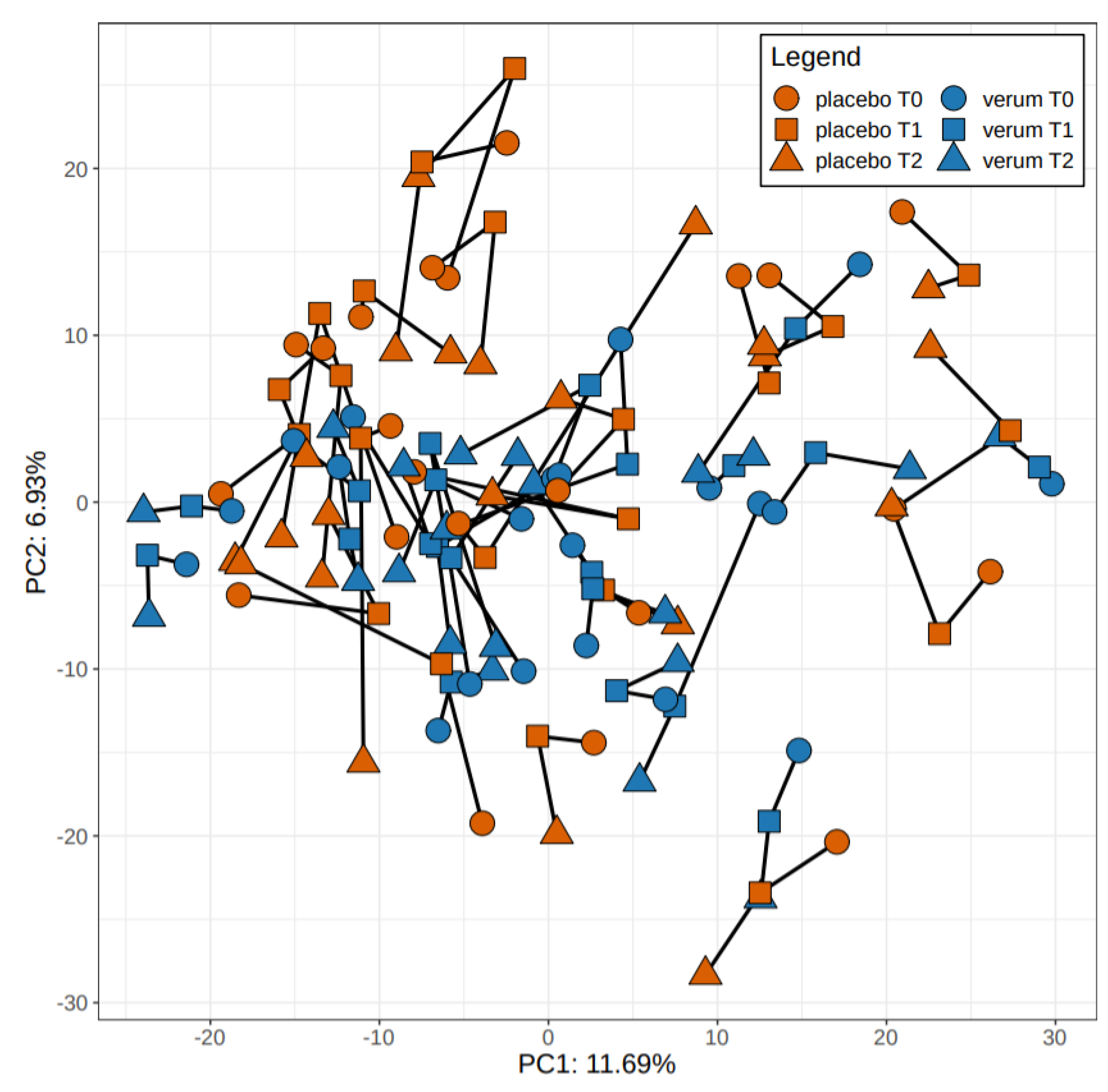
3.4.2. Beta-Diversity
3.4.3. Global Differential Abundance
3.4.4. Piphillin-Analysis
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bauer, M.E.; Teixeira, A.L. Inflammation in psychiatric disorders: What comes first? Ann. N. Y. Acad. Sci. 2019, 1437, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, C.; Fasano, A.; Zonulin, A. Regulator of epithelial and endothelial barrier Functions, and its involvement in chronic inflammatory diseases. Tissue Barriers 2016, 4, e1251384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, R.; Abdolmaleky, H.M.; Zhou, J.R. Microbiome, inflammation, epigenetic alterations, and mental diseases. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2017, 174, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Ait-Belgnaoui, A.; Colom, A.; Braniste, V.; Ramalho, L.; Marrot, A.; Cartier, C.; Houdeau, E.; Theodorou, V.; Tompkins, T. Probiotic gut effect prevents the chronic psychological stress-induced brain activity abnormality in mice. Neurogastroenterol. Motil. 2014, 26, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Frohlich, E.E.; Farzi, A.; Mayerhofer, R.; Reichmann, F.; Jacan, A.; Wagner, B.; Zinser, E.; Bordag, N.; Magnes, C.; Frohlich, E. Cognitive impairment by antibiotic-induced gut dysbiosis: Analysis of gut microbiota-brain communication. Brain Behav. Immun. 2016, 56, 140–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, C.J.K.; Milev, R. The effects of probiotics on depressive symptoms in humans: A systematic review. Ann. Gen. Psychiatry 2017, 16, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoban, A.E.; Stilling, R.M.; Moloney, G.; Shanahan, F.; Dinan, T.G.; Clarke, G.; Cryan, J.F. The microbiome regulates amygdala-dependent fear recall. Mol. Psychiatry 2017, 25, 1134–1144. [Google Scholar]
- Lowry, C.A.; Smith, D.G.; Siebler, P.H.; Schmidt, D.; Stamper, C.E.; Hassell, J.E., Jr.; Yamashita, P.S.; Fox, J.H.; Reber, S.O.; Brenner, L.A. The Microbiota, Immunoregulation, and Mental Health: Implications for Public Health. Curr. Environ. Health Rep. 2016, 3, 270–286. [Google Scholar] [CrossRef] [Green Version]
- Evrensel, A.; Ceylan, M.E. The gut-brain axis: The missing link in depression. Clin. Psychopharmacol. Neurosci. 2015, 13, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Medawar, E.; Huhn, S.; Villringer, A.; Witte, A.V. The effects of plant-based diets on the body and the brain: A systematic review. Transl. Psychiatry 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Madara, J.L. Regulation of the movement of solutes across tight junctions. Ann. Rev. Physiol. 1998, 60, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Not, T.; Wang, W.; Uzzau, S.; Berti, I.; Tommasini, A.; Goldblum, S.E.; Zonulin, A. Newly discovered modulator of intestinal permeability, and its expression in coeliac disease. Lancet 2000, 355, 1518–1519. [Google Scholar] [CrossRef]
- Huang, R.; Wang, K.; Hu, J. Effect of probiotics on depression: A systematic review and meta-analysis of randomized controlled trials. Nutrients 2016, 8, 483. [Google Scholar] [CrossRef]
- Kennedy, R.J.; Kirk, S.J.; Gardiner, K.R. Mucosal barrier function and the commensal flora. Gut 2002, 50, 441–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangiola, F.; Ianiro, G.; Franceschi, F.; Fagiuoli, S.; Gasbarrini, G.; Gasbarrini, A. Gut microbiota in autism and mood disorders. World J. Gastroenterol. 2016, 22, 361. [Google Scholar] [CrossRef]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psych. 2013, 74, 720–726. [Google Scholar] [CrossRef]
- Goh, K.K.; Liu, Y.W.; Kuo, P.H.; Chung, Y.C.E.; Lu, M.L.; Chen, C.H. Effect of probiotics on depressive symptoms: A meta-analysis of human studies. Psych. Res. 2019, 282, 112568. [Google Scholar] [CrossRef]
- Mörkl, S.; Butler, M.I.; Holl, A.; Cyran, J.F.; Dinan, T.G. Probiotics and the microbiota-gut-brain axis: Focus on psychiatry. Curr. Nutr. Rep. 2020. [Google Scholar] [CrossRef]
- Akkasheh, G.; Kashani-Poor, Z.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akbari, H.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z.; Esmaillzadeh, A. Clinical and metabolic response to probiotic administration in patients with major depressive disorder: A randomized, double-blind, placebo-controlled trial. Nutrition 2016, 32, 315–320. [Google Scholar] [CrossRef]
- Ghorbani, Z.; Nazari, S.; Etesam, F.; Nourimajd, S.; Ahmadpanah, M.; Jahromi, S.R. The effect of synbiotic as an adjuvant therapy to fluoxetine in moderate depression: A randomized multicenter trial. Arch. Neurosci. 2018, 5, e60507. [Google Scholar] [CrossRef] [Green Version]
- Kazemi, A.; Noorbala, A.A.; Azam, K.; Eskandari, M.H.; Djafarian, K. Effect of probiotic and prebiotic vs placebo on psychological outcomes in patients with major depressive disorder: A randomized clinical trial. Clin. Nutr. 2019, 38, 522–528. [Google Scholar] [CrossRef]
- Sanada, K.; Nakajima, S.; Kurokawa, S.; Barceló-Soler, A.; Ikuse, D.; Hirata, A.; Yoshizawa, A.; Tomizawa, J.; Salas-Valero, M.; Noda, Y. Gut microbiota and major depressive disorder: A systematic review and meta-analysis. J. Affect. Disord. 2020, 266, 1–13. [Google Scholar] [CrossRef]
- Rudzki, L.; Ostrowska, L.; Pawlak, D.; Malus, A.; Pawlak, K.; Waszkiewicz, N.; Szulc, A. Probiotic Lactobacillus Plantarum 299v decreases kynurenine concentration and improves cognitive functions in patients with major depression: A double-blind, randomized, placebo controlled study. Psychoneuroendocrinology 2019, 100, 213–222. [Google Scholar] [CrossRef]
- Reininghaus, E.Z.; Wetzlmair, L.C.; Fellendorf, F.T.; Platzer, M.; Queissner, R.; Birner, A.; Pilz, R.; Hamm, C.; Maget, A.; Koidl, C.; et al. The impact of probiotic supplements on cognitive parameters in euthymic individuals with bipolar disorder: A pilot study. Neuropsychobiolgy 2018, 79, 1–8. [Google Scholar] [CrossRef]
- Reiter, A.; Bengesser, S.A.; Hauschild, A.-C.; Birkl-Töglhofer, A.-M.; Fellendorf, F.T.; Platzer, M.; Färber, T.; Seidl, M.; Mendel, L.-M.; Unterweger, R. Interleukin-6 gene expression changes after a 4-week intake of a multispecies probiotic in major depressive disorder—Preliminary Results of the PROVIT Study. Nutrients 2020, 12, 2575. [Google Scholar] [CrossRef]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The mini-international neuropsychiatric interview (M.I.N.I.): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psych. 1998, 59 (Suppl. 20), 22–33. [Google Scholar]
- Hamilton, M.A. Rating scale for depression. J. Neurol. Neurosurg. Psych. 1960, 23, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Kuhner, C.; Burger, C.; Keller, F.; Hautzinger, M. Reliability and validity of the revised beck depression inventory (BDI-II). Results from GERMAN samples. Nervenarzt 2007, 78, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Franke, G.H.; Derogatis, L.R. Die Symptom-Checkliste von Derogatis: Manual. SCL-90-R; Deutsche Version; Beltz Test: Gottingen, Germany, 1995. [Google Scholar]
- Krüger, S.; Bräunig, P.; Shugar, G. Manie-Selbstbeurteilungsskala: MSS; Manual Beltz Test: Weinheim, Germany, 1997. [Google Scholar]
- Eypasch, E.; Wood-Dauphinee, S.; Williams, J.I.; Ure, B.; Neugebauer, E.; Troidl, H. The Gastrointestinal Quality of Life Index. A clinical index for measuring patient status in gastroenterologic surgery. Der Chirurg Zeitschrift für alle Gebiete der Operativen Medizin 1993, 64, 264–274. [Google Scholar]
- Roager, H.M.; Hansen, L.B.; Bahl, M.I.; Frandsen, H.L.; Carvalho, V.; Gøbel, R.J.; Dalgaard, M.D.; Plichta, D.R.; Sparholt, M.H.; Vestergaard, H. Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut. Nat. Microbiol. 2016, 1, 16093. [Google Scholar] [CrossRef]
- White, M.A.; Whisenhunt, B.L.; Williamson, D.A.; Greenway, F.L.; Netemeyer, R.G. Development and validation of the food-craving inventory. Obes. Res. 2002, 10, 107–114. [Google Scholar] [CrossRef]
- Klymiuk, I.; Bilgilier, C.; Stadlmann, A.; Thannesberger, J.; Kastner, M.T.; Högenauer, C.; Püspök, A.; Biowski-Frotz, S.; Schrutka-Kölbl, C.; Thallinger, G.G. The human gastric microbiome is predicated upon infection with helicobacter pylori. Front. Microbiol. 2017, 8, 2508. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.Z.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. Accurate taxonomy assignments from 16S rRNA sequences produced by highly parallel pyrosequencers. Nucl. Acids. Res. 2008, 36, e120. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.G.; Garrity, M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Sieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar]
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome datasets are compositional: And this is not optional. Front. Microbiol. 2017, 8, 2224. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; McMurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Package ‘Vegan’ Austria 2019; R Package Version 2.5-6; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [Green Version]
- Iwai, S.; Weinmaier, T.; Schmidt, B.L.; Albertson, D.G.; Poloso, N.J.; Dabbagh, K.; DeSantis, T.Z. Piphillin: Improved prediction of metagenomic content by direct inference from human microbiomes. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Ai, D.; Pan, H.; Li, X.; Gao, Y.; Liu, G.; Xia, L.C. Identifying gut microbiota associated with colorectal cancer using a zero-inflated lognormal model. Front. Microbiol. 2019, 10, 826. [Google Scholar] [CrossRef]
- Domingo, M.C.; Huletsky, A.; Boissinot, M.; Bernard, K.A.; Picard, F.J.; Bergeron, M.G. Ruminococcus gauvreauii sp. nov., a glycopeptide-resistant species isolated from a human faecal specimen. Int. J. Syst. Evol Microbiol. 2008, 58, 1393–1397. [Google Scholar] [CrossRef]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of human gut microbiota and short-chain fatty acids in response to dietary interventions with three fermentable fibers. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Painold, A.; Mörkl, S.; Kashofer, K.; Halwasch, B.; Dalkner, N.; Bengesser, S.; Birner, A.; Fellendorf, F.; Platzer, M.; Queissner, R. A step ahead: Exploring the gut microbiota in inpatients with bipolar disorder during a depressive episode. Bipolar Disord. 2019, 21, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Berni Canani, R.; Flint, H.J.; Salminen, S. Expert consensus document. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laursen, M.F.; Laursen, R.P.; Larnkjær, A.; Michaelsen, K.F.; Bahl, M.I.; Licht, T.R. Administration of two probiotic strains during early childhood does not affect the endogenous gut microbiota composition despite probiotic proliferation. BMC Microbiol. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martoni, C.J.; Evans, M.; Chow, C.E.T.; Chan, L.S.; Leyer, G. Impact of a probiotic product on bowel habits and microbial profile in participants with functional constipation: A randomized controlled trial. J. Dig. Dis. 2019, 20, 435–446. [Google Scholar] [CrossRef]
- Parra, M.; Stahl, S.; Hellmann, H. Vitamin B6 and its role in cell metabolism and physiology. Cells 2018, 7, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustafa, A.A.; Hewedi, D.H.; Eissa, A.M.; Frydecka, D.; Misiak, B. Homocysteine levels in schizophrenia and affective disorders-focus on cognition. Front. Behav. Neurosci. 2014, 8, 343. [Google Scholar] [CrossRef] [Green Version]
- Reininghaus, B.; Riedrich, K.; Dalkner, N.; Lehner, L.A.; Rieger, A.; Hamm, C.; Dorn, M.; Gradauer, L.; Hufnagl, A.; Mayr-Mauhart, M. Physical health in individuals with psychiatric disorders in Austria. J. Affect. Disord. 2019, 257, 38–44. [Google Scholar] [CrossRef]
- Slyepchenko, A.; Maes, M.; Jacka, F.N.; Köhler, C.A.; Barichello, T.; McIntyre, R.S.; Berk, M.; Grande, I.; Foster, J.A.; Vieta, E. Gut microbiota, bacterial translocation, and interactions with diet: Pathophysiological links between major depressive disorder and non-communicable medical comorbidities. Psychother. Psychosom. 2017, 86, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.J. Intestinal flora and endogenous vitamin synthesis. Eur. J. Cancer Prev. 1997, 6 (Suppl. 1), S43–S45. [Google Scholar] [CrossRef] [PubMed]
- Mörkl, S.; Wagner-Skacel, J.; Lahousen, T.; Lackner, S.; Holasek, S.J.; Bengesser, S.A.; Painold, A.; Holl, A.K.; Reininghaus, E.Z. The Role of Nutrition and the Gut-Brain Axis in Psychiatry: A Review of the Literature]. Neuropsychobiolgy 2018, 1–9. [Google Scholar] [CrossRef]
- Ohlsson, L.; Gustafsson, A.; Lavant, E.; Suneson, K.; Brundin, L.; Westrin, Å.; Ljunggren, L.; Lindqvist, D. Leaky gut biomarkers in depression and suicidal behavior. Acta Psychiatr. Scand. 2019, 139, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Stevens, B.R.; Goel, R.; Seungbum, K.; Richards, E.M.; Holbert, R.C.; Pepine, C.J.; Raizada, M.K. Increased human intestinal barrier permeability plasma biomarkers zonulin and FABP2 correlated with plasma LPS and altered gut microbiome in anxiety or depression. Gut 2018, 67, 1555–1557. [Google Scholar] [CrossRef]
- Mokkala, K.; Pussinen, P.; Houttu, N.; Koivuniemi, E.; Vahlberg, T.; Laitinen, K. The impact of probiotics and n-3 long-chain polyunsaturated fatty acids on intestinal permeability in pregnancy: A randomised clinical trial. Eur. J. Clin. Nutr. 2017, 71, 1455–1462. [Google Scholar] [CrossRef]
- De Roos, N.M.; van Hemert, S.; Rovers, J.M.P.; Smits, M.G.; Witteman, B.J.M. The effects of a multispecies probiotic on migraine and markers of intestinal permeability-results of a randomized placebo-controlled study. Eur. J. Clin. Nutr. 2017, 71, 1455–1462. [Google Scholar] [CrossRef]
- Amatya, N.; Garg, A.V.; Gaffen, S.L. IL-17 Signaling: The Yin and the Yang. Trends Immunol. 2017, 38, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Abusleme, L.; Hong, B.Y.; Hoare, A.; Konkel, J.E.; Diaz, P.I.; Moutsopoulos, N.M. Oral Microbiome characterization in murine models. Bio-protocol 2017, 7, e2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Tato, C.M.; Joyce-Shaikh, B.; Gulen, M.F.; Cayatte, C.; Chen, Y.; Blumenschein, W.M.; Judo, M.; Ayanoglu, G.; McClanahan, T.K. Interleukin-23-dependent IL-17 production regulates intestinal epithelial permeability. Immunity 2015, 43, 727–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karp, D.R.; Marthandan, N.; Marsh, S.G.; Ahn, C.; Arnett, F.C.; Deluca, D.S.; Diehl, A.D.; Dunivin, R.; Eilbeck, K.; Feolo, M. Novel sequence feature variant type analysis of the HLA genetic association in systemic sclerosis. Hum. Mol. Genet. 2010, 19, 707–719. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Description | Intervention Group | Placebo Group | Statistics | |
|---|---|---|---|---|
| (n = 28) | (n = 33) | |||
| N (%) | N (%) | χ2 | Sig (p) | |
| Sex (female) | 20 (71.4%) | 27 (81.8%) | 0.925 | 0.336 |
| Smoking (yes) | 9 (32.1%) | 19 (57.6%) | 3.946 | 0.047 |
| Mean (SD) | Mean (SD) | T | Sig (p) | |
| Age (years) | 43.00 (14.31) | 40.11 (11.45) | −0.876 | 0.384 |
| HAMD | 15.07 (6.32) | 14.73 (4.59) | 0.246 | 0.807 |
| BDI-II | 30.75 (8.40) | 32.60 (10.93) | −0.719 | 0.475 |
| BMI (kg/m2) | 26.29 (5.78) | 25.74 (7.29) | −0.319 | 0.751 |
| Waist-to-hip ratio | 0.86 (0.07) | 0.84 (0.10) | −0.739 | 0.463 |
| Education (years) | 11.04 (2.87) | 10.45 (2.05) | −0.919 | 0.362 |
| Illness duration (years) | 11.40 (13.52) | 11.15 (8.34) | −0.090 | 0.929 |
| Scores | Intervention Group (n = 28) | Placebo Group (n = 30) | Time | Group | Time * Group | |||
|---|---|---|---|---|---|---|---|---|
| Mean (SD) | Mean (SD) | F | p | F | p | F | p | |
| HAMD t0 | 15.07 (6.32) | 14.43 (4.41) | 47.853 | 0.000 | 0.482 | 0.490 | 0.036 | 0.850 |
| HAMD t2 | 9.11 (5.16) | 8.13 (6.16) | ||||||
| BDI-II t0 | 30.75 (8.40) | 32.6 (10.93) | 114.635 | 0.000 | 1.284 | 0.262 | 0.196 | 0.660 |
| BDI-II t2 | 15.11 (7.91) | 18.2 (11.53) | ||||||
| MSS t0 | 7.18 (5.67) | 8.37 (7.18) | 4.882 | 0.031 | 0.923 | .341 | 0.029 | 0.866 |
| MSS t2 | 5.43 (4.61) | 6.87 (5.78) | ||||||
| GSI t0 | 67.68 (5.68) | 67.20 (8.52) | 64.293 | 0.000 | 0.056 | 0.813 | 0.882 | 0.352 |
| GSI t2 | 58.36 (9.79) | 59.83 (10.76) | ||||||
| PST t0 | 66.93 (6.72) | 66.23 (8.89) | 34.866 | 0.000 | 0.001 | 0.978 | 0.476 | 0.493 |
| PST t2 | 59.68 (11.28) | 60.50 (10.80) | ||||||
| PSDI t0 | 65.71 (5.47) | 65.20 (8.48) | 58.700 | 0.000 | 0.001 | 0.977 | 0.233 | 0.631 |
| SCL-R PSDI t2 | 56.07 (9.18) | 56.70 (10.68) | ||||||
| GIQL t0 | 75.89 (17.07) | 76.37 (16.24) | 47.841 | 0.000 | 0.032 | 0.859 | 0.018 | 0.895 |
| GIQL t2 | 88.81 (17.85) | 89.80 (17.22) | ||||||
| Zonulin Concentrations | Intervention Group (n = 28) | Placebo Group (n = 31) | Time | Group | Time * Group | |||
|---|---|---|---|---|---|---|---|---|
| Mean (SD) | Mean (SD) | f | p | f | p | f | p | |
| Zonulin [ng/mL] t0 | 46.801 (15.957) | 52.007 (10.906) | 0.560 | 0.457 | 1.662 | 0.202 | 0.426 | 0.516 |
| Zonulin [ng/mL] t2 | 50.161 (15.909) | 52.236 (13.827) | ||||||
| KEGG Pathway | Probiotics Versus Placebo, T2, Effect Size | Name of Pathway |
|---|---|---|
| KO04657 | 0.463 | IL-17 signaling pathway |
| KO00780 | 0.432 | Biotin (Vitamin B7) metabolism |
| KO04910 | 0.424 | Insulin signaling pathway |
| KO00750 | 0.410 | Vitamin B6 metabolism |
| KO00500 | 0.404 | Starch and sucrose metabolism |
| KO05010 | 0.384 | Alzheimer disease |
| KO00730 | 0.370 | Thiamine (vitamin B1) metabolism |
| KO00400 | 0.368 | Phenylalanine, tyrosine and tryptophan biosynthesis |
| KO00190 | 0.363 | Oxidative phosphorylation |
| KO00770 | 0.362 | Pantothenate and CoA biosynthesis |
| KO01100 | 0.362 | Metabolic pathways |
| KO00760 | 0.359 | Nicotinate and nicotinamide metabolism |
| KO00471 | 0.358 | D-Glutamine and D-glutamate metabolism |
| KO01040 | 0.355 | Biosynthesis of unsaturated fatty acids |
| KO00620 | 0.349 | Pyruvate metabolism |
| KO00290 | 0.340 | Valine, leucine and isoleucine biosynthesis |
| KO00061 | 0.332 | Fatty acid biosynthesis |
| KO00010 | 0.326 | Glycolysis/Gluconeogenesis |
| KO04724 | 0.326 | Glutamatergic synapse |
| KO01212 | 0.323 | Fatty acid metabolism |
| KO00640 | 0.319 | Propanoate metabolism |
| KO00020 | 0.309 | Citrate cycle (TCA cycle) |
| KO04727 | 0.294 | GABAergic synapse |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reininghaus, E.Z.; Platzer, M.; Kohlhammer-Dohr, A.; Hamm, C.; Mörkl, S.; Bengesser, S.A.; Fellendorf, F.T.; Lahousen-Luxenberger, T.; Leitner-Afschar, B.; Schöggl, H.; et al. PROVIT: Supplementary Probiotic Treatment and Vitamin B7 in Depression—A Randomized Controlled Trial. Nutrients 2020, 12, 3422. https://doi.org/10.3390/nu12113422
Reininghaus EZ, Platzer M, Kohlhammer-Dohr A, Hamm C, Mörkl S, Bengesser SA, Fellendorf FT, Lahousen-Luxenberger T, Leitner-Afschar B, Schöggl H, et al. PROVIT: Supplementary Probiotic Treatment and Vitamin B7 in Depression—A Randomized Controlled Trial. Nutrients. 2020; 12(11):3422. https://doi.org/10.3390/nu12113422
Chicago/Turabian StyleReininghaus, Eva Z., Martina Platzer, Alexandra Kohlhammer-Dohr, Carlo Hamm, Sabrina Mörkl, Susanne A. Bengesser, Frederike T. Fellendorf, Theressa Lahousen-Luxenberger, Birgitta Leitner-Afschar, Helmut Schöggl, and et al. 2020. "PROVIT: Supplementary Probiotic Treatment and Vitamin B7 in Depression—A Randomized Controlled Trial" Nutrients 12, no. 11: 3422. https://doi.org/10.3390/nu12113422
APA StyleReininghaus, E. Z., Platzer, M., Kohlhammer-Dohr, A., Hamm, C., Mörkl, S., Bengesser, S. A., Fellendorf, F. T., Lahousen-Luxenberger, T., Leitner-Afschar, B., Schöggl, H., Amberger-Otti, D., Wurm, W., Queissner, R., Birner, A., Falzberger, V. S., Painold, A., Fitz, W., Brunnmayr, M., Rieger, A., ... Dalkner, N. (2020). PROVIT: Supplementary Probiotic Treatment and Vitamin B7 in Depression—A Randomized Controlled Trial. Nutrients, 12(11), 3422. https://doi.org/10.3390/nu12113422