Influence of Pomace Matrix and Cyclodextrin Encapsulation on Olive Pomace Polyphenols’ Bioaccessibility and Intestinal Permeability




Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Reagents
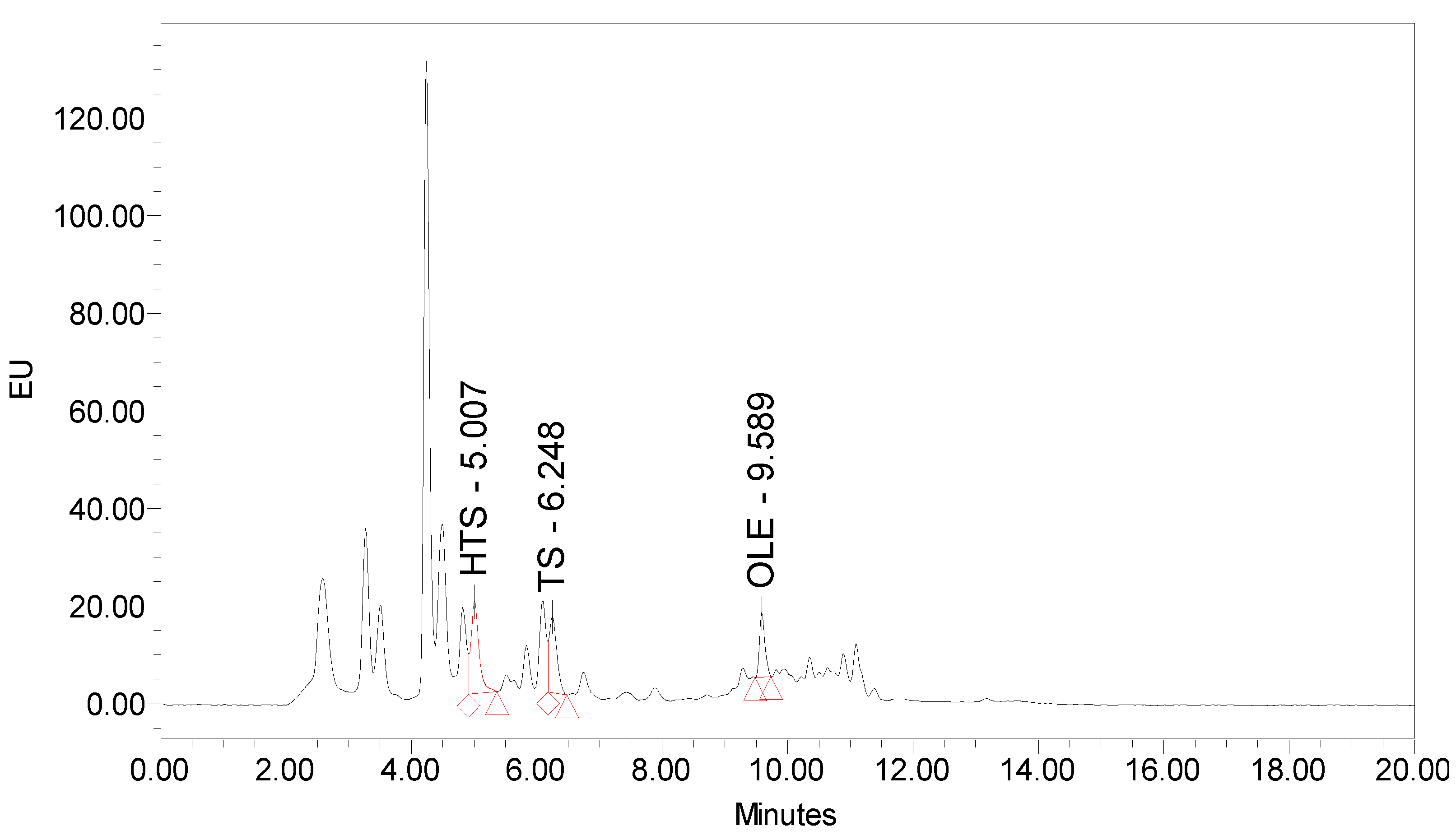
2.3. Quantification of Phenolic Components by HPLC-FLD and Determination of the Radical Scavenging Capacity by TEAC
2.4. In Vitro Simulated Gastrointestinal Digestion
2.5. In Vitro Study of Transepithelial Transport
2.6. Determination of Antioxidative Activity
2.7. Statistical Analysis
3. Results and Discussion
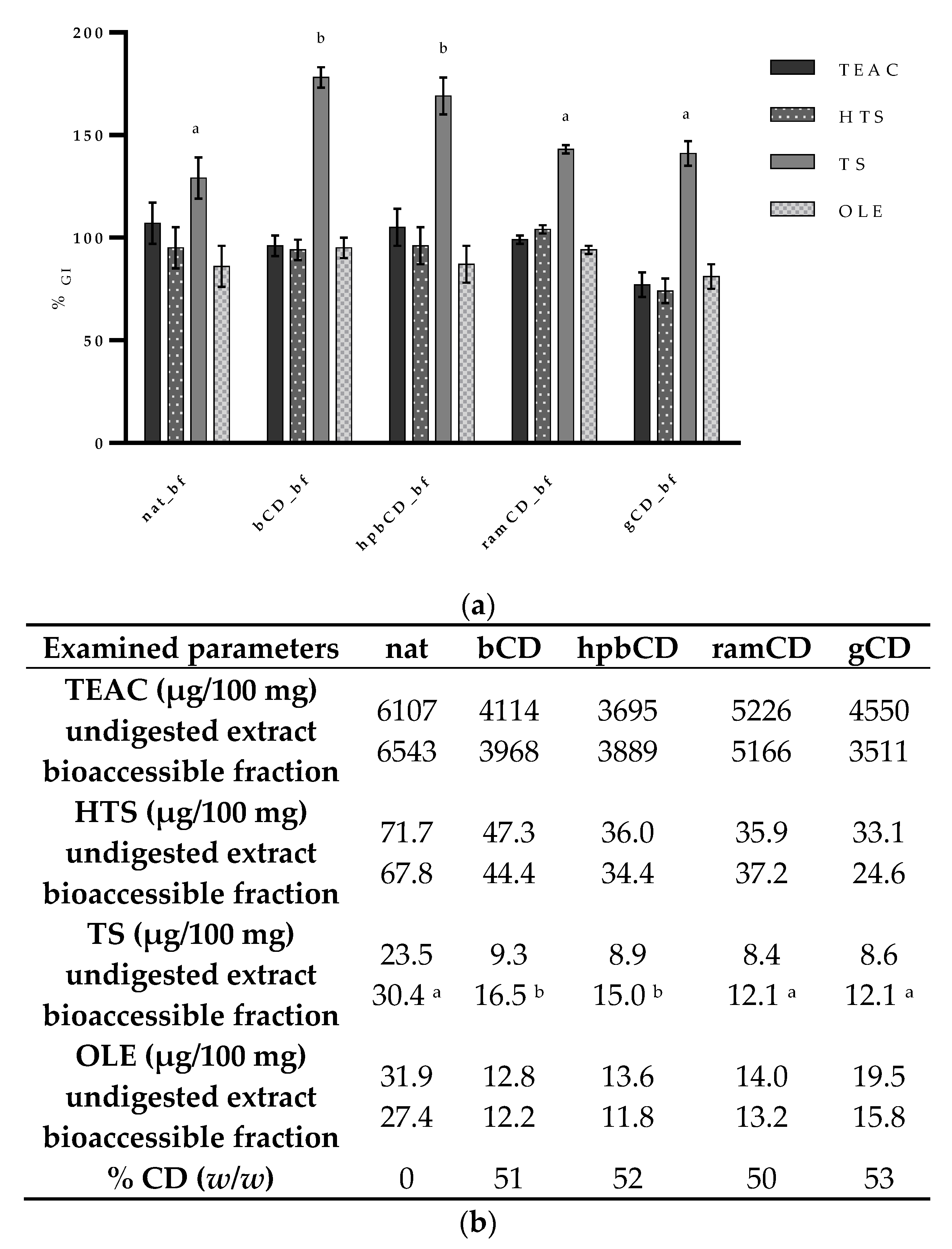
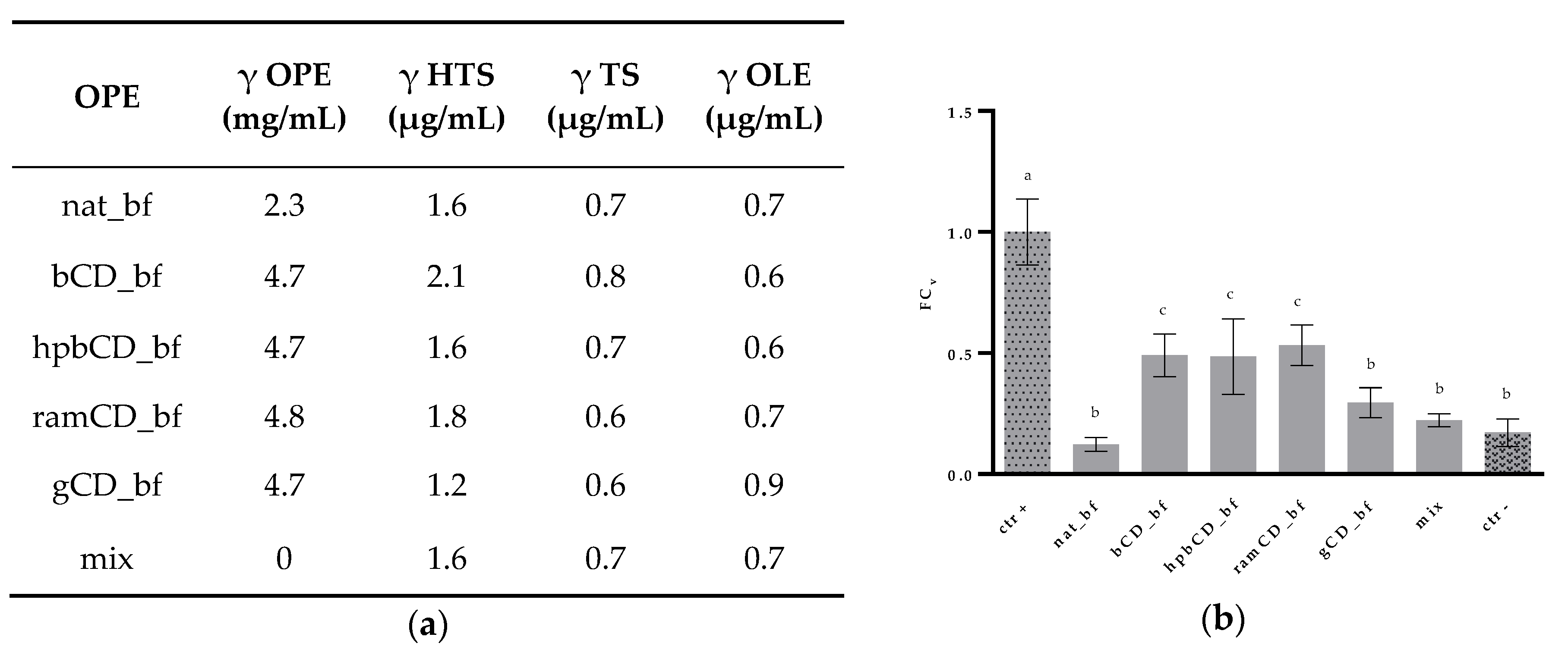
3.1. Composition of Olive Pomace Extracts
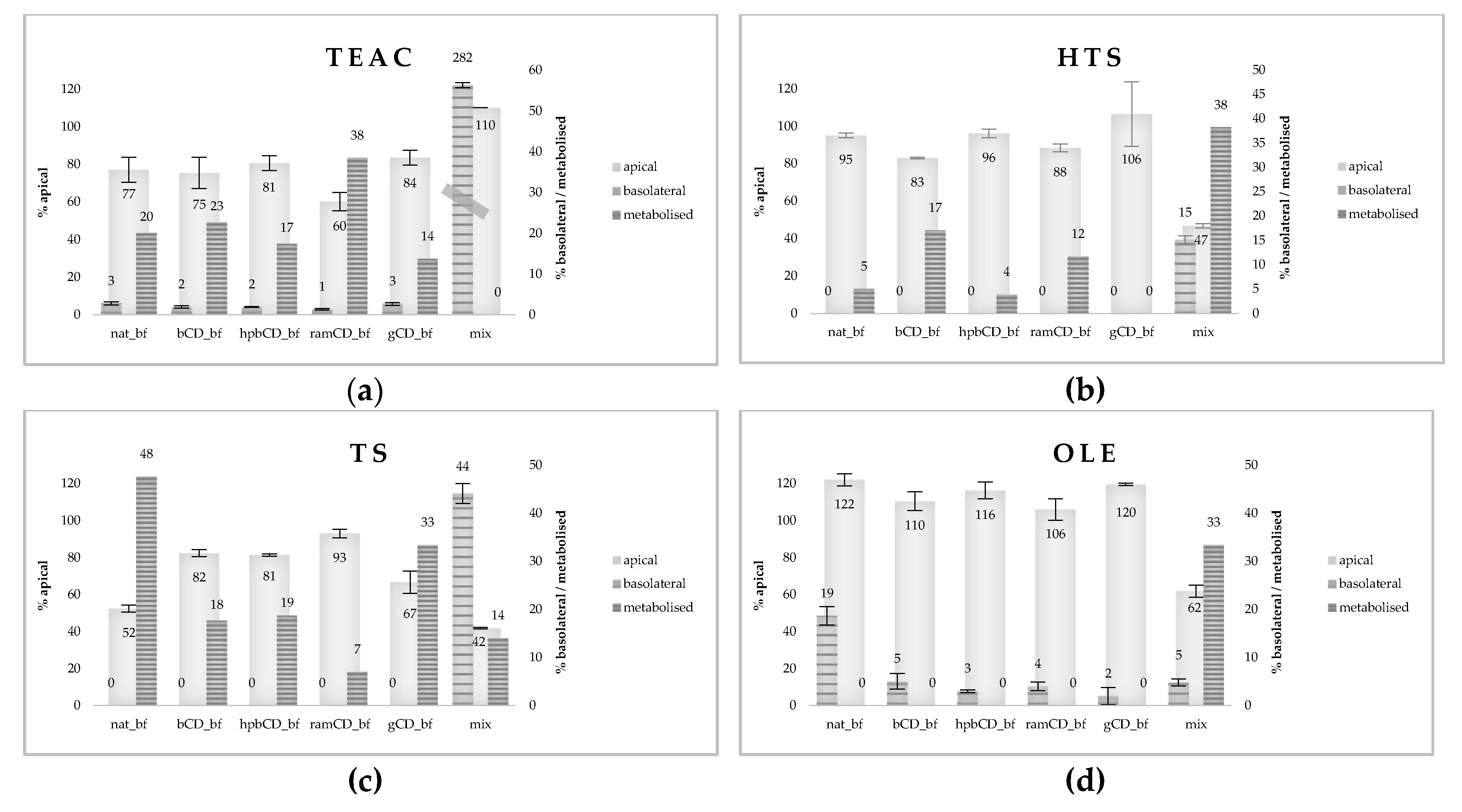
3.2. Bioaccessibility of Olive Pomace Polyphenols
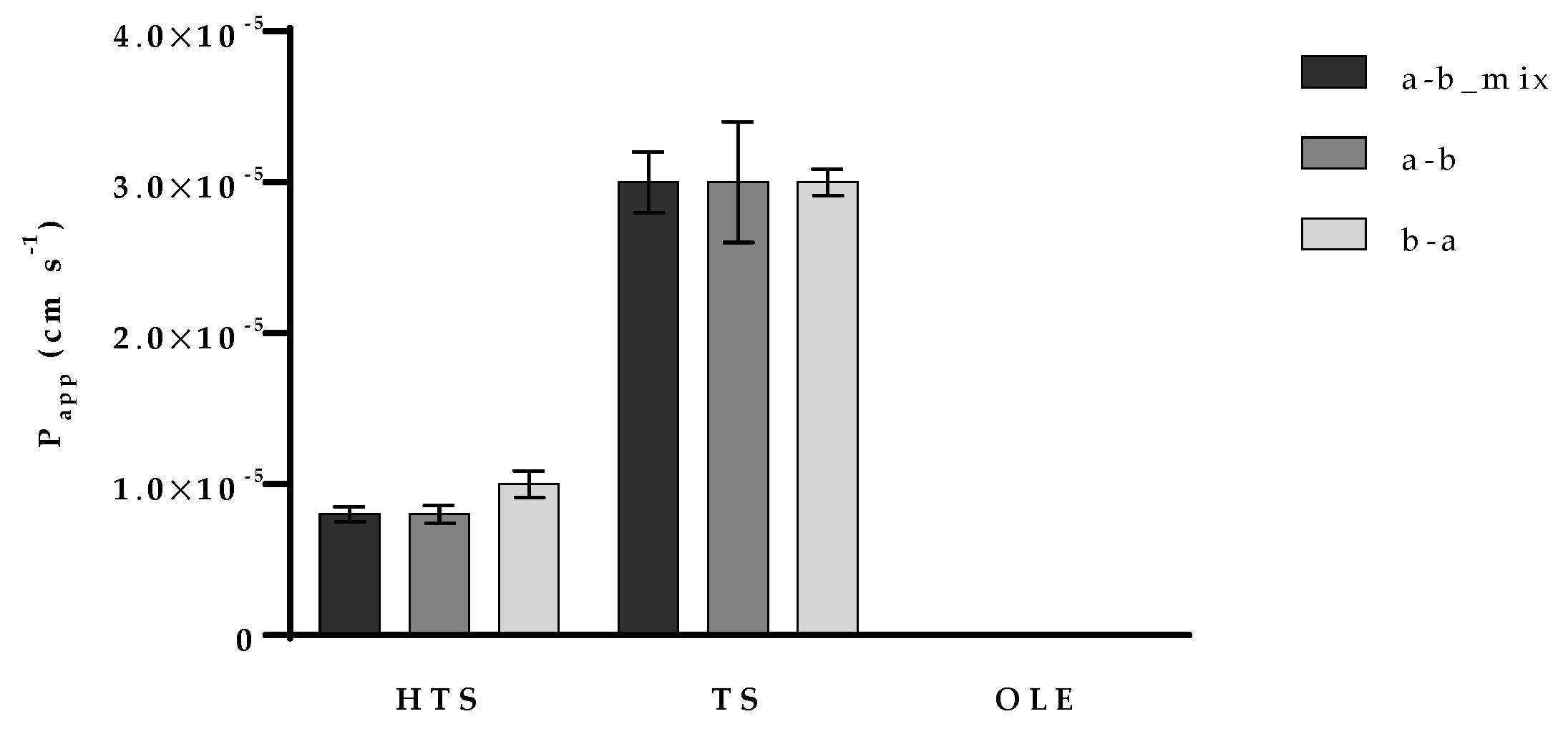
3.3. Permeability of OPE Polyphenols on Caco-2 Cell Monolayer
3.4. Antioxidative Effect of OPE on Caco-2 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ntougias, S.; Bourtzis, K.; Tsiamis, G. The microbiology of olive mill wastes. BioMed Res. Int. 2013, 2013, 784591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topi, D.; Beqiraj, I.; Seiti, B.; Halimi, E. Environmental impact from olive mills waste disposal, chemical analysis of solid wastes and wastewaters. J. Hyg. Eng. Des. 2014, 7, 44–48. [Google Scholar]
- Herrero, M.; Temirzoda, T.N.; Segura-Carretero, A.; Quirantes, R.; Plaza, M.; Ibañez, E. New possibilities for the valorization of olive oil by-products. J. Chromatogr. A 2011, 1218, 7511–7520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverría, F.; Ortiz, M.; Valenzuela, R.; Videla, L.; Echeverría, F.; Ortiz, M.; Valenzuela, R.; Videla, L.A. Hydroxytyrosol and cytoprotection: A projection for clinical interventions. Int. J. Mol. Sci. 2017, 18, 930. [Google Scholar] [CrossRef]
- EFSA. Oxidative Damage, Maintenance of Normal Blood HDL Cholesterol Concentrations Mainte; EFSA: Parma, Italy, 2011.
- Jurmanović, S.; Jug, M.; Safner, T.; Radić, K.; Domijan, A.-M.; Pedisić, S.; Šimić, S.; Jablan, J.; Vitali Čepo, D. Utilization of olive pomace as the source of polyphenols: Optimization of microwave-assisted extraction and characterization of spray-dried extract. J. Food Nutr. Res. 2019, 58, 51–62. [Google Scholar]
- Albahari, P.; Jug, M.; Radić, K.; Jurmanović, S.; Brnčić, M.; Brnčić, S.R.; Vitali Čepo, D. Characterization of olive pomace extract obtained by cyclodextrin-enhanced pulsed ultrasound assisted extraction. Lwt—Food Sci. Technol. 2018, 92, 22–31. [Google Scholar] [CrossRef]
- FDA. Subject: Notice of a GRAS Exclusion for Alpha-Cyclodextrin; FDA: White Oak, MD, USA, 2016; ISBN 7722990746.
- Corp, W.B. GRAS Notice 000074: BETA-CYCLODEXTRIN; HFS: Washington, DC, USA, 2001; Volume 28. [Google Scholar]
- Clissold, D.B. Notice of a GRAS Exemption for Gamma-Cyclodextrin; FDA: White Oak, MD, USA, 2000; pp. 1–49.
- Soler, A.; Romero, M.P.; Macià, A.; Saha, S.; Furniss, C.S.M.; Kroon, P.A.; Motilva, M.J. Digestion stability and evaluation of the metabolism and transport of olive oil phenols in the human small-intestinal epithelial Caco-2/TC7 cell line. Food Chem. 2010, 119, 703–714. [Google Scholar] [CrossRef]
- Rigacci, S.; Stefani, M. Nutraceutical properties of olive oil polyphenols. An itinerary from cultured cells through animal models to humans. Int. J. Mol. Sci. 2016, 17, 843. [Google Scholar] [CrossRef] [Green Version]
- Corona, G.; Tzounis, X.; Dessì, M.A.; Deiana, M.; Debnam, E.S.; Visioli, F.; Spencer, J.P.E. The fate of olive oil polyphenols in the gastrointestinal tract: Implications of gastric and colonic microflora-dependent biotransformation. Free Radic. Res. 2006, 40, 647–658. [Google Scholar] [CrossRef]
- Malapert, A.; Tomao, V.; Margier, M.; Nowicki, M.; Gleize, B.; Dangles, O.; Reboul, E. Β-Cyclodextrin Does Not Alter the Bioaccessibility and the Uptake By Caco-2 Cells of Olive By-Product Phenolic Compounds. Nutrients 2018, 10, 1653. [Google Scholar] [CrossRef] [Green Version]
- Malapert, A.; Tomao, V.; Dangles, O.; Reboul, E. Effect of foods and β-cyclodextrin on the bioaccessibility and the uptake by caco-2 cells of hydroxytyrosol from either a pure standard or alperujo. J. Agric. Food Chem. 2018, 66, 4614–4620. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, R.F.S.; Martins, J.T.; Duarte, C.M.M.; Vicente, A.A.; Pinheiro, A.C. Advances in nutraceutical delivery systems: From formulation design for bioavailability enhancement to efficacy and safety evaluation. Trends Food. Sci. Technol. 2018, 78, 270–291. [Google Scholar] [CrossRef] [Green Version]
- Vitali Čepo, D.; Radić, K.; Jurmanović, S.; Jug, M.; Rajković, M.G.; Pedisić, S.; Moslavac, T.; Albahari, P. Valorization of olive pomace-based nutraceuticals as antioxidants in chemical, food, and biological models. Molecules 2018, 23, 2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, S.B.; Duarte, F.Í.C.; Heimfarth, L.; Quintans, J.D.S.S.; Quintans-Júnior, L.J.; Júnior, V.F.D.V.; De Lima, Á.A.N. Cyclodextrin-drug inclusion complexes: In vivo and in vitro approaches. Int. J. Mol. Sci. 2019, 20, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansook, P.; Ogawa, N.; Loftsson, T. Cyclodextrins: Structure, physicochemical properties and pharmaceutical applications. Int. J. Pharm. 2018, 535, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zu, Y.; Zhao, X.; Wu, M.; Feng, Z.; Deng, Y.; Zu, C.; Wang, L. Preparation of inclusion complex of apigenin-hydroxypropyl-β-cyclodextrin by using supercritical antisolvent process for dissolution and bioavailability enhancement. Int. J. Pharm. 2016, 511, 921–930. [Google Scholar] [CrossRef]
- Soo, E.; Thakur, S.; Qu, Z.; Jambhrunkar, S.; Parekh, H.S.; Popat, A. Enhancing delivery and cytotoxicity of resveratrol through a dual nanoencapsulation approach. J. Colloid Interface Sci. 2016, 462, 368–374. [Google Scholar] [CrossRef] [Green Version]
- Sharayei, P.; Azarpazhooh, E.; Ramaswamy, H.S. Effect of microencapsulation on antioxidant and antifungal properties of aqueous extract of pomegranate peel. J. Food Sci. Technol. 2019. [Google Scholar] [CrossRef]
- Mady, F.M.; Ibrahim, S.R.M. Cyclodextrin-based nanosponge for improvement of solubility and oral bioavailability of Ellagic acid. Pak. J. Pharm. Sci. 2018, 31, 2069–2076. [Google Scholar]
- Rezaei, A.; Varshosaz, J.; Fesharaki, M.; Farhang, A.; Jafari, S.M. Improving the solubility and in vitro cytotoxicity (Anticancer activity) of ferulic acid by loading it into cyclodextrin nanosponges. Int. J. Nanomed. 2019, 14, 4589–4599. [Google Scholar] [CrossRef] [Green Version]
- Tsarbopoulos, A.; Gikas, E.; Papadopoulos, N.; Aligiannis, N.; Kafatos, A. Simultaneous determination of oleuropein and its metabolites in plasma by high-performance liquid chromatography. J. Chromatogr. B 2003, 785, 157–164. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Benink, H.A.; Worzlla, T.J.; Minor, L. Cell Viability Assays; Assay Guidance Manual; National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004; pp. 1–23.
- Hubatsch, I.; Ragnarsson, E.G.E.; Artursson, P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat. Protoc. 2007, 2, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.; Kalla, A.M.; Rajunaik, B.; Kumar, A. Food matrix: A new tool to enhance nutritional quality of food. J. Pharm. Phytochem. 2018, 7, 1011–1014. [Google Scholar]
- Visioli, F.; Galli, C.; Bornet, F.; Mattei, A.; Patelli, R.; Galli, G.; Caruso, D. Olive oil phenolics are dose-dependently absorbed in humans Francesco. FEBS Lett. 2000, 468, 159–160. [Google Scholar] [CrossRef]
- Seiquer, I.; Rueda, A.; Olalla, M.; Cabrera-Vique, C. Assessing the bioavailability of polyphenols and antioxidant properties of extra virgin argan oil by simulated digestion and Caco-2 cell assays. Comparative study with extra virgin olive oil. Food Chem. 2015, 188, 496–503. [Google Scholar] [CrossRef]
- Vissers, M.N.; Zock, P.L.; Roodenburg, A.J.C.; Leenen, R.; Katan, M.B. Olive oil phenols are absorbed in humans. J. Nutr. 2002, 132, 409–417. [Google Scholar] [CrossRef]
- Corona, G.; Spencer, J.; Dessì, M. Extra virgin olive oil phenolics: Absorption, metabolism, and biological activities in the GI tract. Toxicol. Ind. Health 2009, 25, 285–293. [Google Scholar] [CrossRef]
- Cardinali, A.; Linsalata, V.; Lattanzio, V.; Ferruzzi, M.G. Verbascosides from olive mill waste water: assessment of their bioaccessibility and intestinal uptake using an in vitro digestion/caco-2 model system. J. Food Sci. 2011, 76, H48–H54. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, G.; Zang, X.; Ye, F. Optimization, antioxidant activity and bile salts adsorption capacity of the aqueous enzymatic extract from rice bran. Czech Acad. Agric. Sci. 2018, 2018, 338–348. [Google Scholar]
- Garcia-padial, M.; Oharriz, M. Complexation of tyrosol with cyclodextrins Complexation of tyrosol with cyclodextrins. J. Incl. Phenom. Macrocycl. Chem. 2012, 75, 241–246. [Google Scholar] [CrossRef]
- Di Benedetto, R.; Varì, R.; Scazzocchio, B.; Filesi, C.; Santangelo, C.; Giovannini, C.; Matarrese, P.; D’Archivio, M.; Masella, R. Tyrosol, the major extra virgin olive oil compound, restored intracellular antioxidant defences in spite of its weak antioxidative effectiveness. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 535–545. [Google Scholar] [CrossRef]
- Markopoulos, C.; Vertzoni, M.; Agalias, A.; Magiatis, P.; Reppas, C. Stability of oleuropein in the human proximal gut. J. Pharm. Pharmacol. 2009, 61, 143–149. [Google Scholar] [CrossRef]
- Dinnella, C.; Minichino, P.; D’Andrea, A.M.; Monteleone, E. Bioaccessibility and antioxidant activity stability of phenolic compounds from extra-virgin olive oils during in vitro digestion. J. Agric. Food Chem. 2007, 55, 8423–8429. [Google Scholar] [CrossRef]
- Rosignoli, P.; Fuccelli, R.; Sepporta, M.V.; Fabiani, R. In vitro chemo-preventive activities of hydroxytyrosol: The main phenolic compound present in extra-virgin olive oil. Food Funct. 2016, 7, 301–307. [Google Scholar] [CrossRef]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef]
- D’Antuono, I.; Garbetta, A.; Ciasca, B.; Linsalata, V.; Minervini, F.; Lattanzio, V.M.T.; Logrieco, A.F.; Cardinali, A. Biophenols from table olive CV bella di cerignola: chemical characterization, bioaccessibility, and intestinal absorption. J. Agric. Food Chem. 2016, 64, 5671–5678. [Google Scholar] [CrossRef]
- Mateos, R.; Pereira-Caro, G.; Saha, S.; Cert, R.; Redondo-Horcajo, M.; Bravo, L.; Kroon, P.A. Acetylation of hydroxytyrosol enhances its transport across differentiated Caco-2 cell monolayers. Food Chem. 2011, 125, 865–872. [Google Scholar] [CrossRef]
- Manna, C.; Galletti, P.; Maisto, G.; Cucciolla, V.; D’Angelo, S.; Zappia, V. Transport mechanism and metabolism of olive oil hydroxytyrosol in Caco-2 cells. FEBS Lett. 2000, 470, 341–344. [Google Scholar] [CrossRef] [Green Version]
- Le Bourvellec, C.; Renard, C.M.G.C. Interactions between polyphenols and macromolecules: Quantification methods and mechanisms. Crit. Rev. Food Sci. Nutr. 2012, 52, 213–248. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, F.; Kiss, T.; Fenyvesi, E.; Szente, L.; Veszelka, S.; Deli, M.A.; Varadi, J.; Feher, P.; Ujhelyi, Z.; Tosaki, A. Randomly methylated β-cyclodextrin derivatives enhance taxol permeability through human intestinal epithelial caco-2 cell. J. Pharm. Sci. 2011, 100, 4734–4744. [Google Scholar] [CrossRef] [PubMed]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Morató, J.; Boronat, A.; Kotronoulas, A.; Pujadas, M.; Pastor, A.; Olesti, E.; Pérez-Mañá, C.; Khymenets, O.; Fitó, M.; Farré, M. Metabolic disposition and biological significance of simple phenols of dietary origin: Hydroxytyrosol and tyrosol. Drug Metab. Rev. 2016, 48, 218–236. [Google Scholar] [CrossRef] [Green Version]
- Pinho, E.; Grootveld, M.; Soares, G.; Henriques, M. Cyclodextrins as encapsulation agents for plant bioactive compounds. Carbohydr. Polym. 2014, 101, 121–135. [Google Scholar] [CrossRef] [Green Version]

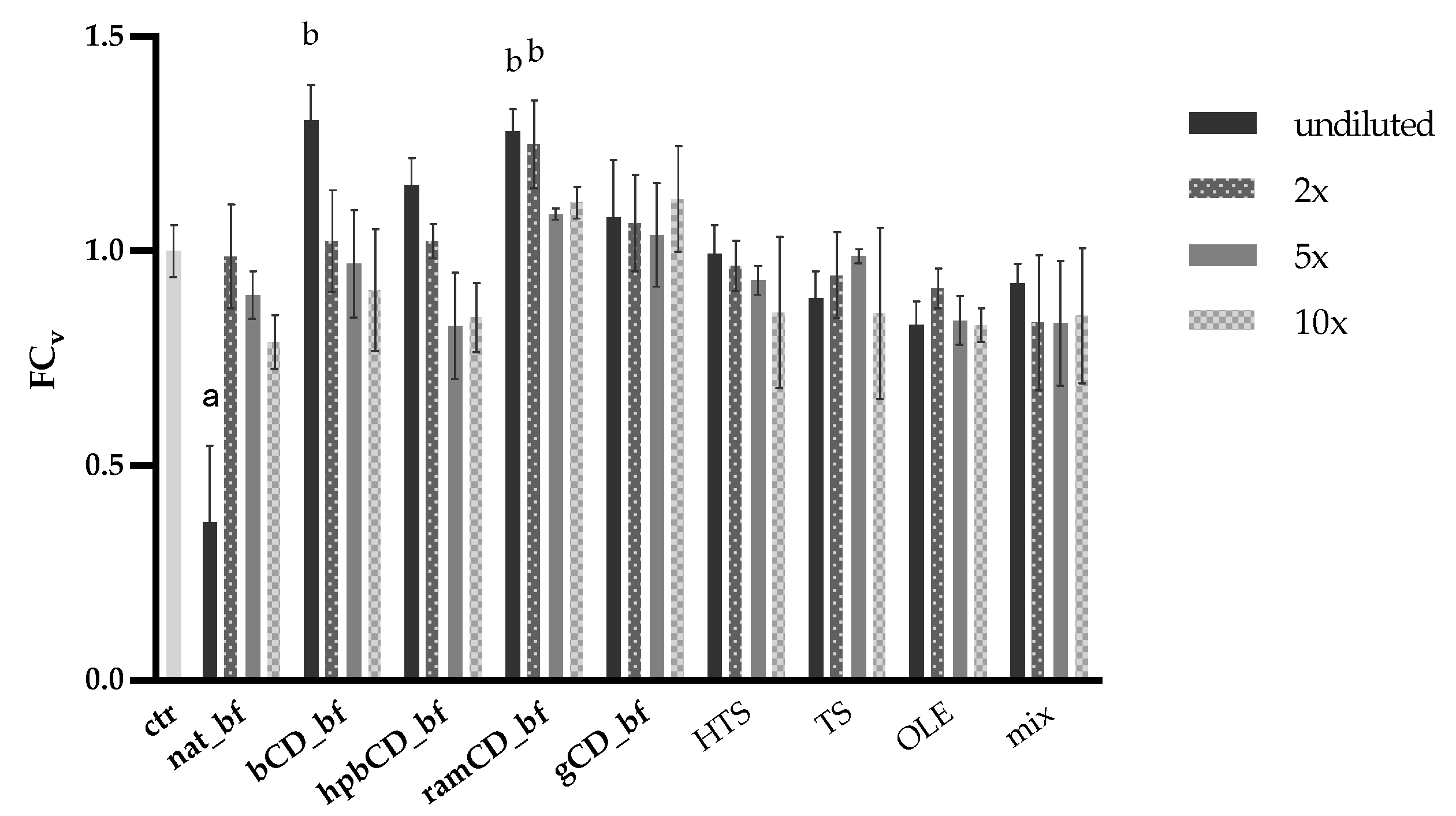




{kind=link}
{kind=link}
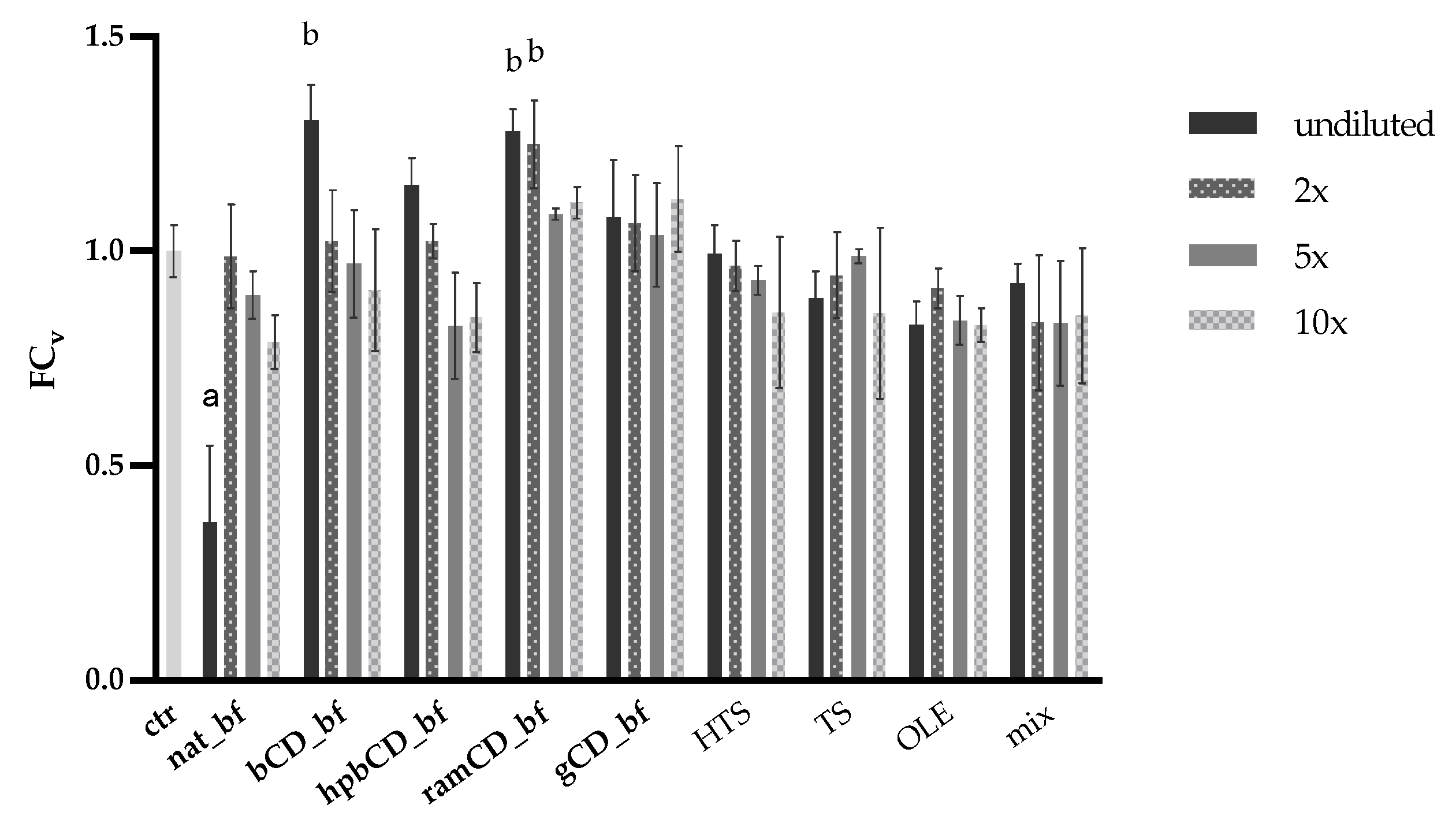{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OPE | HTS (µg/100 mg) | TS (µg/100 mg) | OLE (µg/100 mg) | TEAC (µg/100 mg) | % CD (w/w) |
|---|---|---|---|---|---|
| nat | 71.7 | 23.5 | 31.9 | 6107 | 0 |
| bCD | 47.3 | 9.3 | 12.8 | 4114 | 51 |
| hpbCD | 36.0 | 8.9 | 13.6 | 3695 | 52 |
| ramCD | 35.9 | 8.4 | 14.0 | 5226 | 50 |
| gCD | 33.1 | 8.6 | 19.5 | 4550 | 53 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radić, K.; Jurišić Dukovski, B.; Vitali Čepo, D. Influence of Pomace Matrix and Cyclodextrin Encapsulation on Olive Pomace Polyphenols’ Bioaccessibility and Intestinal Permeability. Nutrients 2020, 12, 669. https://doi.org/10.3390/nu12030669
Radić K, Jurišić Dukovski B, Vitali Čepo D. Influence of Pomace Matrix and Cyclodextrin Encapsulation on Olive Pomace Polyphenols’ Bioaccessibility and Intestinal Permeability. Nutrients. 2020; 12(3):669. https://doi.org/10.3390/nu12030669
Chicago/Turabian StyleRadić, Kristina, Bisera Jurišić Dukovski, and Dubravka Vitali Čepo. 2020. "Influence of Pomace Matrix and Cyclodextrin Encapsulation on Olive Pomace Polyphenols’ Bioaccessibility and Intestinal Permeability" Nutrients 12, no. 3: 669. https://doi.org/10.3390/nu12030669
APA StyleRadić, K., Jurišić Dukovski, B., & Vitali Čepo, D. (2020). Influence of Pomace Matrix and Cyclodextrin Encapsulation on Olive Pomace Polyphenols’ Bioaccessibility and Intestinal Permeability. Nutrients, 12(3), 669. https://doi.org/10.3390/nu12030669