Does Caesarean Section or Preterm Delivery Influence TGF-β2 Concentrations in Human Colostrum?

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Design
2.2. Study Population
2.3. Collection of Human Colostrum (HC)
2.4. Quantification of Transforming Growth Factor β (TGF-β2) in Human Colostrum
2.5. Statistical Analysis
2.6. Ethical Approval
3. Results
3.1. Description of Study Participants
3.2. Measurements of TGF-β2 Concentrations
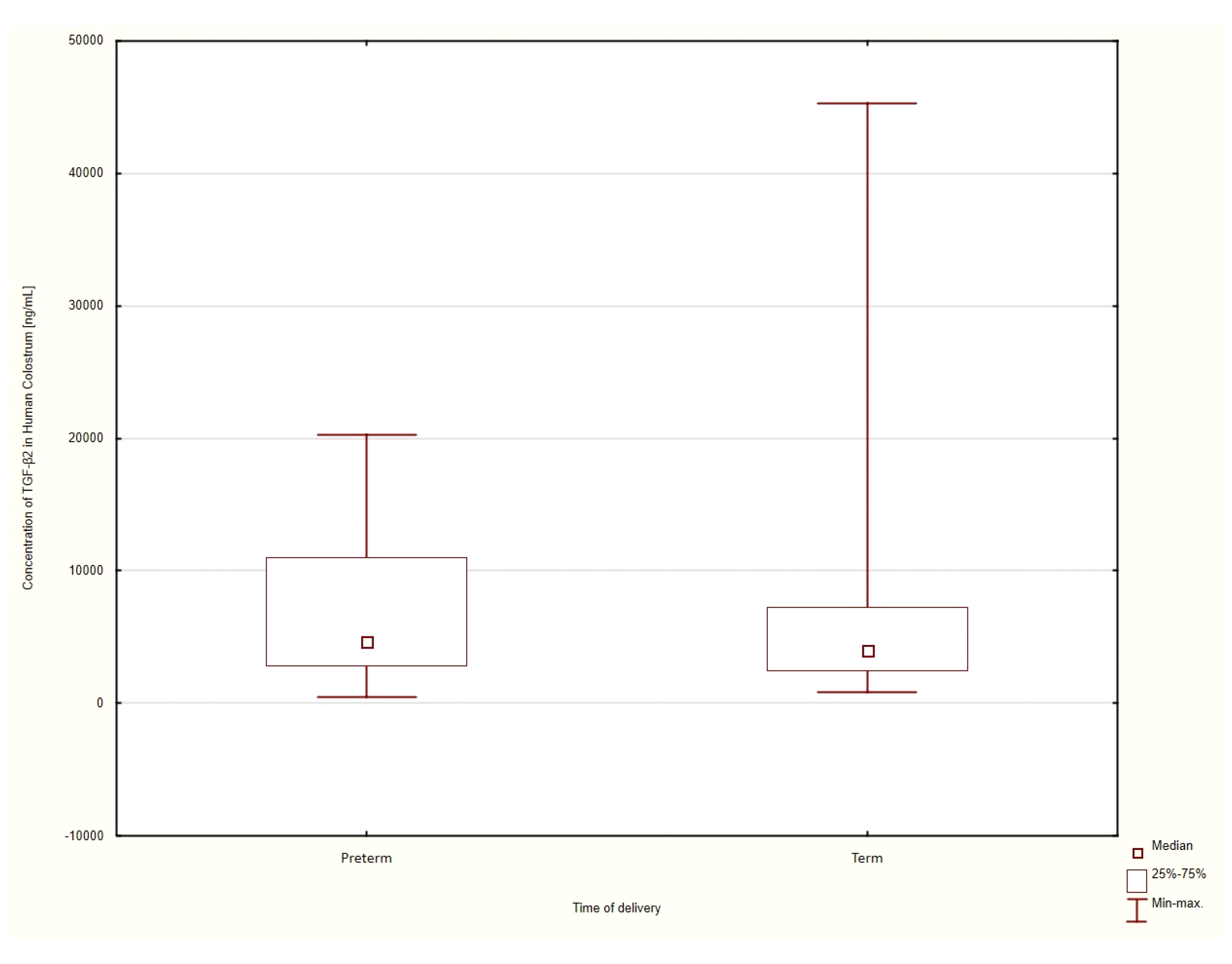
3.3. Gestational Age at the Time of Delivery
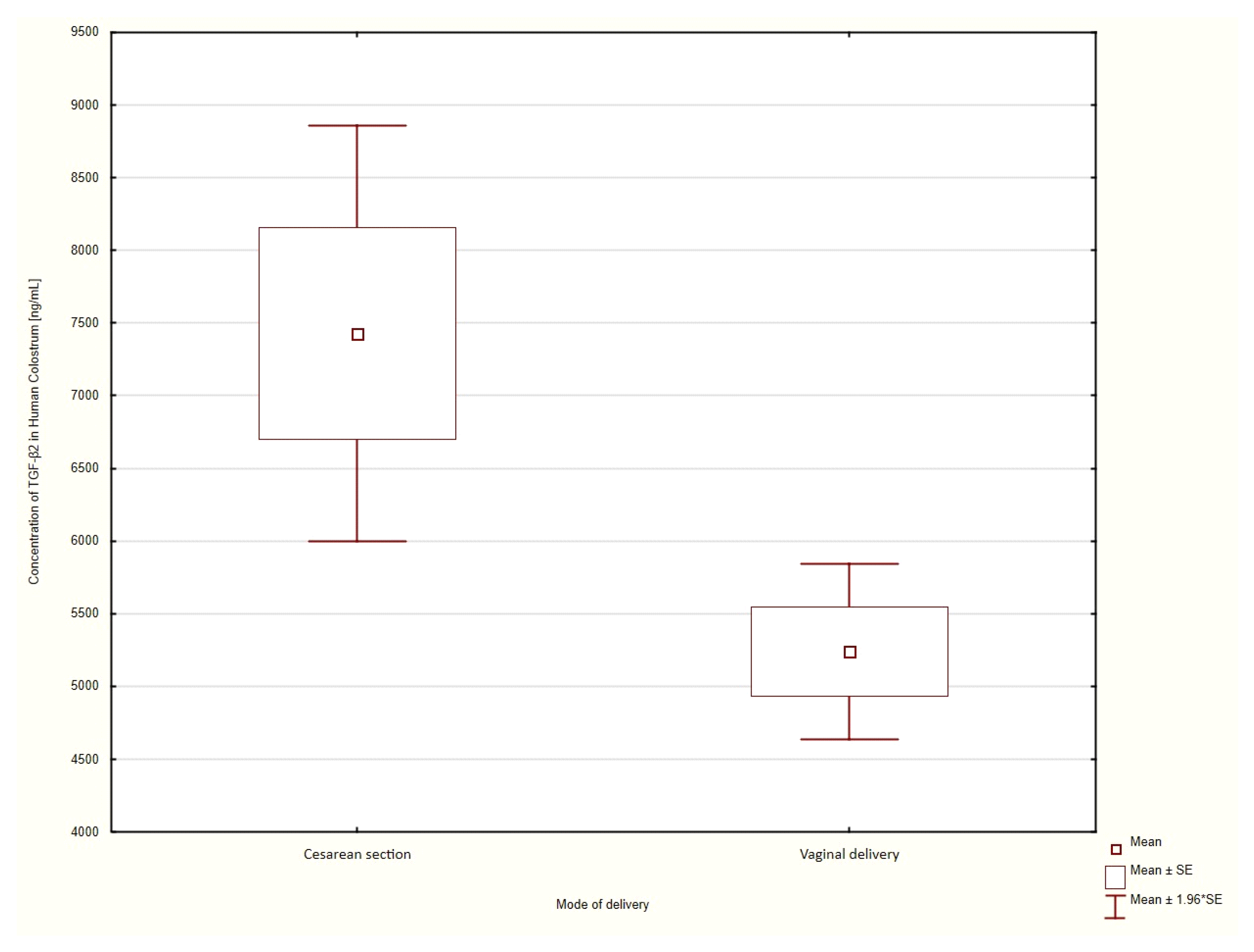
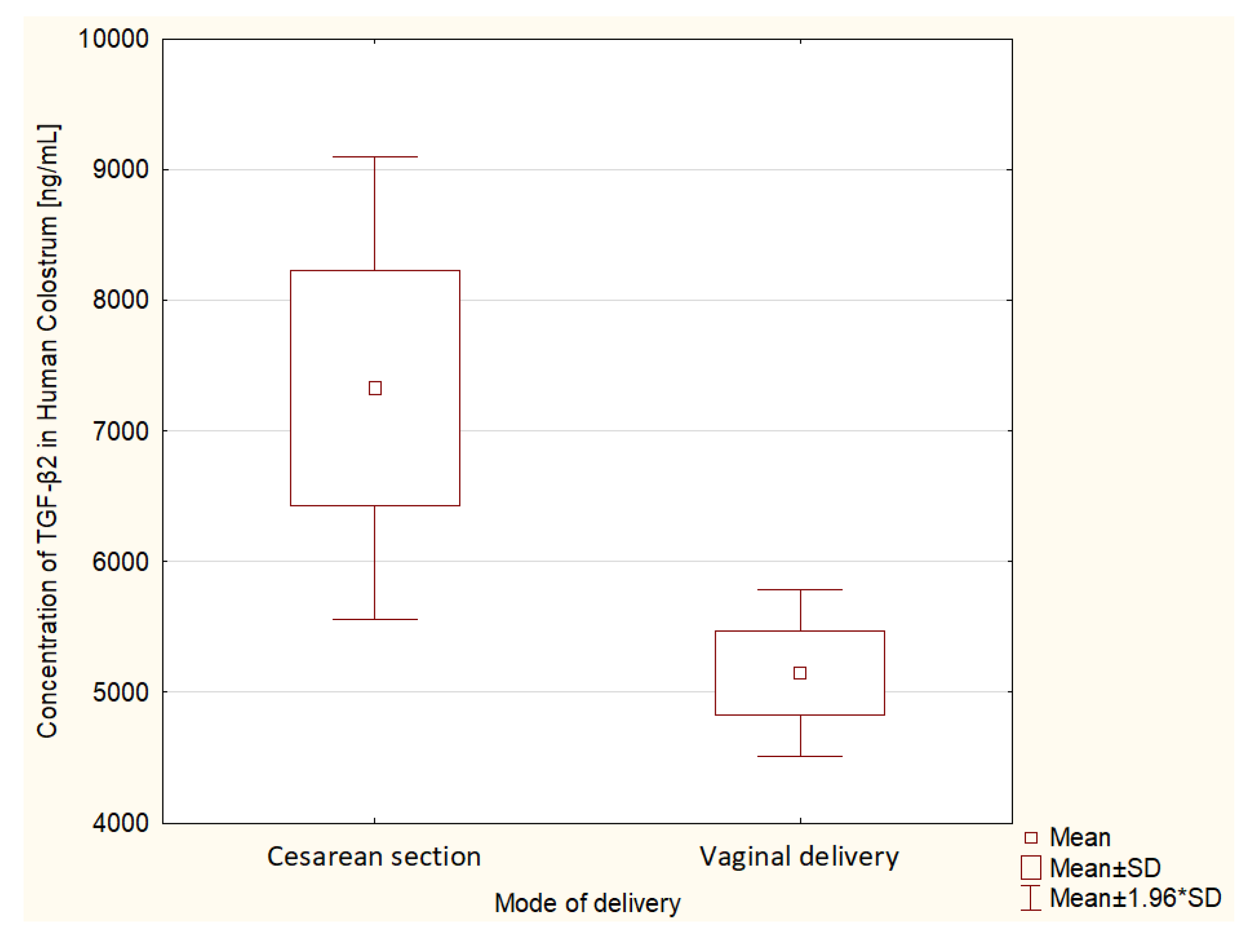
3.4. Mode of Delivery
Term Delivery
4. Discussion
5. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Castilho, S.D.; Barros Filho, A.A. The history of infant nutrition. J. Pediatr. (Rio J.) 2010, 86, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Fewtrell, M.S.; Mohd Shukri, N.H.; Wells, J.C.K. ‘Optimising’ breastfeeding: What can we learn from evolutionary, comparative and anthropological aspects of lactation? BMC Med. 2020, 18, 4. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Parks, O.B.; Good, M. A Review of the Immunomodulating Components of Maternal Breast Milk and Protection against Necrotizing Enterocolitis. Nutrients 2020, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Laouar, A. Maternal Leukocytes and Infant Immune Programming during Breastfeeding. Trends Immunol. 2020, 41, 225–239. [Google Scholar] [CrossRef]
- Matsumoto, N.; Yorifuji, T.; Nakamura, K.; Ikeda, M.; Tsukahara, H.; Doi, H. Breastfeeding and risk of food allergy: A nationwide birth cohort in Japan. Allergol. Int. 2020, 69, 91–97. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, K.M.; Jung, H.L.; Shim, J.Y.; Kim, D.S.; Shim, J.W. Relationship between Breastfeeding, Birth History, and Acute Pyelonephritis in Infants. J. Korean Med. Sci. 2020, 35, e32. [Google Scholar] [CrossRef]
- Munblit, D.; Abrol, P.; Sheth, S.; Chow, L.Y.; Khaleva, E.; Asmanov, A.; Lauriola, S.; Padovani, E.M.; Comberiati, P.; Boner, A.L.; et al. Levels of Growth Factors and IgA in the Colostrum of Women from Burundi and Italy. Nutrients 2018, 10, 1216. [Google Scholar] [CrossRef]
- Newburg, D.; Walker, W. Protection of the Neonate by the Innate Immune System of Developing Gut and of Human Milk. Pediatr. Res. 2007, 61, 2–8. [Google Scholar] [CrossRef]
- Munblit, D.; Peroni, D.G.; Boix-Amorós, A.; Hsu, P.S.; Land, B.V.; Gay, M.C.L.; Kolotilina, A.; Skevaki, C.; Boyle, R.J.; Collado, M.C.; et al. Human Milk and Allergic Diseases: An Unsolved Puzzle. Nutrients 2017, 9, 894. [Google Scholar] [CrossRef]
- Munblit, D.; Treneva, M.; Peroni, D.G.; Colicino, S.; Chow, L.; Dissanayeke, S.; Abrol, P.; Sheth, S.; Pampura, A.; Boner, A.L.; et al. Colostrum and Mature Human Milk of Women from London, Moscow, and Verona: Determinants of Immune Composition. Nutrients 2016, 8, 695. [Google Scholar] [CrossRef]
- Ahnfeldt, A.M.; Hyldig, N.; Li, Y.; Kappel, S.S.; Aunsholdt, L.; Sangild, P.T.; Zachariassen, G. FortiColos—A multicentre study using bovine colostrum as a fortifier to human milk in very preterm infants: Study protocol for a randomised controlled pilot trial. Trials 2019, 20, 279. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, J.S.; Bryan, D.L.; James, M.J.; Gibson, R.A. Cytokines (IL-1β, IL-6, TNF-α, TGF-β1, and TGF-β2) and Prostaglandin E2 in Human Milk during the First Three Months Postpartum. Pediatr. Res. 1999, 46, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Kalliomäki, M.; Ouwehand, A.; Arvilommi, H.; Kero, P.; Isolauri, E. Transforming growth factor-beta in breast milk: A potential regulator of atopic disease at an early age. J. Allergy Clin. Immunol. 1999, 104, 1251–1257. [Google Scholar] [CrossRef]
- Saito, S.; Yoshida, M.; Ichijo, M.; Ishizaka, S.; Tsujii, T. Transforming growth factor-beta (TGF-beta) in human milk. Clin. Exp. Immunol. 1993, 94, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Lu, L.; Nanthakumar, N.N.; Dubert-Ferrandon, A.; Walker, W.A. TGF-beta2 induces maturation of immature human intestinal epithelial cells and inhibits inflammatory cytokine responses induced via the NF-kappaB pathway. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 630–638. [Google Scholar] [CrossRef]
- Brenmoehl, J.; Ohde, D.; Wirthgen, E.; Hoeflich, A. Cytokines in milk and the role of TGF-beta. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 47–56. [Google Scholar] [CrossRef]
- Maheshwari, A.; Kelly, D.R.; Nicola, T.; Ambalavanan, N.; Jain, S.K.; Murphy-Ullrich, J.; Athar, M.; Shimamura, M.; Bhandari, V.; Aprahamian, C.; et al. TGF-β2 suppresses macrophage cytokine production and mucosal inflammatory responses in the developing intestine. Gastroenterology 2011, 140, 242–253. [Google Scholar] [CrossRef]
- Frost, B.L.; Jilling, T.; Lapin, B.; Maheshwari, A.; Caplan, M.S. Maternal breast milk transforming growth factor-beta and feeding intolerance in preterm infants. Pediatr. Res. 2014, 76, 386–393. [Google Scholar] [CrossRef]
- Khaleva, E.; Gridneva, Z.; Geddes, D.T.; Oddy, W.H.; Colicino, S.; Blyuss, O.; Boyle, R.J.; Warner, J.O.; Munblit, D. Transforming growth factor beta in human milk and allergic outcomes in children: A systematic review. Clin. Exp. Allergy 2019, 49, 1201–1213. [Google Scholar] [CrossRef]
- Sitarik, A.R.; Bobbitt, K.R.; Havstad, S.L.; Fujimura, K.E.; Levin, A.M.; Zoratti, E.M.; Kim, H.; Woodcroft, K.J.; Wegienka, G.; Ownby, D.R.; et al. Breast Milk Transforming Growth Factor β Is Associated with Neonatal Gut Microbial Composition. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 60–67. [Google Scholar] [CrossRef]
- Xiong, W.; Frasch, S.C.; Thomas, S.M.; Bratton, D.L.; Henson, P.M. Induction of TGF-β1 synthesis by macrophages in response to apoptotic cells requires activation of the scavenger receptor CD36. PloS ONE 2013, 8, e72772. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [PubMed]
- Cacho, N.T.; Lawrence, R.M. Innate Immunity and Breast Milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Campos-Alberto, E.; Yamaide, F.; Nakano, T.; Ohnisi, H.; Kawamoto, M.; Kawamoto, N.; Matsui, E.; Kondo, N.; Kohno, Y.; et al. TGF-β Concentration in Breast Milk is Associated With the Development of Eczema in Infants. Front. Pediatr. 2018, 6, 162. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.Y.; Flavell, R.A. TGF-beta and regulatory T cell in immunity and autoimmunity. J. Clin. Immunol. 2008, 28, 647–659. [Google Scholar] [CrossRef]
- Robertson, I.B.; Rifkin, D.B. Regulation of the bioavailability of TGF-b and TGF-b-related proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a021907. [Google Scholar] [CrossRef]
- Taylor, A.W. Review of the activation of TGF-β in immunity. J. Leukoc. Biol. 2009, 85, 29–33. [Google Scholar] [CrossRef]
- Ogawa, J.; Sasahara, A.; Yoshida, T.; Sira, M.M.; Futatani, T.; Kanegane, H.; Miyawaki, T. Role of transforming growth factor-beta in breast milk for initiation of IgA production in newborn infants. Early Hum. Dev. 2004, 77, 67–75. [Google Scholar] [CrossRef]
- Namachivayam, K.; Blanco, C.L.; Frost, B.L.; Reeves, A.A.; Jagadeeswaran, R.; MohanKumar, K.; Safarulla, A.; Mandal, P.; Garzon, S.A.; Raj, J.U.; et al. Preterm human milk contains a large pool of latent TGF-β, which can be activated by exogenous neuraminidase. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G1055–G1065. [Google Scholar] [CrossRef]
- Walraven, M.; Beelen, R.H.; Ulrich, M.M. Transforming growth factor-β (TGF-β) signaling in healthy human fetal skin: A descriptive study. J. Dermatol. Sci. 2015, 78, 117–124. [Google Scholar] [CrossRef]
- Gilbert, R.; Vickaryous, M.K.; Viloria-Petit, A.M. Signalling by Transforming Growth Factor Beta Isoforms in Wound Healing and Tissue Regeneration. J. Dev. Biol. 2016, 4, 21. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| Informed consent to participate in the study | Lack of informed consent to participate in the study |
| Vaginal delivery | Assisted vaginal delivery |
| Surgical delivery | Active mother’s infection |
| Adequate colostrum lactation for the collection of samples | Breast infection/inflammations |
| Single pregnancy | Chronic antibiotic therapy |
| Autoimmune diseases | |
| Gestational diabetes | |
| HELLP * syndrome | |
| Preeclampsia and eclampsia | |
| Multiple pregnancy | |
| Smoking |
| Number of Patients | Mean Gestational Age ± Standard Deviation (SD) | Mean TGF-β2 Concentration in HC [ng/mL] | SD | |
|---|---|---|---|---|
| Extremely preterm, ≤28 WG | 3 | 27.00 ± 1.73 | 3685 | ±1032 |
| Very preterm, 29–32 WG | 6 | 26.50 ± 1.70 | 4235 | ±664 |
| Late preterm, 33–366/7 WG | 39 | 34.95 ± 2.94 | 6026 | ±622 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kociszewska-Najman, B.; Sibanda, E.; Radomska-Leśniewska, D.M.; Taradaj, K.; Kociołek, P.; Ginda, T.; Gruszfeld, M.; Jankowska-Steifer, E.; Pietrzak, B.; Wielgoś, M.; et al. Does Caesarean Section or Preterm Delivery Influence TGF-β2 Concentrations in Human Colostrum? Nutrients 2020, 12, 1095. https://doi.org/10.3390/nu12041095
Kociszewska-Najman B, Sibanda E, Radomska-Leśniewska DM, Taradaj K, Kociołek P, Ginda T, Gruszfeld M, Jankowska-Steifer E, Pietrzak B, Wielgoś M, et al. Does Caesarean Section or Preterm Delivery Influence TGF-β2 Concentrations in Human Colostrum? Nutrients. 2020; 12(4):1095. https://doi.org/10.3390/nu12041095
Chicago/Turabian StyleKociszewska-Najman, Bożena, Elopy Sibanda, Dorota M. Radomska-Leśniewska, Karol Taradaj, Patrycja Kociołek, Tomasz Ginda, Monika Gruszfeld, Ewa Jankowska-Steifer, Bronisława Pietrzak, Mirosław Wielgoś, and et al. 2020. "Does Caesarean Section or Preterm Delivery Influence TGF-β2 Concentrations in Human Colostrum?" Nutrients 12, no. 4: 1095. https://doi.org/10.3390/nu12041095
APA StyleKociszewska-Najman, B., Sibanda, E., Radomska-Leśniewska, D. M., Taradaj, K., Kociołek, P., Ginda, T., Gruszfeld, M., Jankowska-Steifer, E., Pietrzak, B., Wielgoś, M., & Malejczyk, J. (2020). Does Caesarean Section or Preterm Delivery Influence TGF-β2 Concentrations in Human Colostrum? Nutrients, 12(4), 1095. https://doi.org/10.3390/nu12041095