Progressive Metabolic Dysfunction and Nutritional Variability Precedes Necrotizing Enterocolitis



 and
and
Abstract
:1. Introduction
2. Materials and Methods
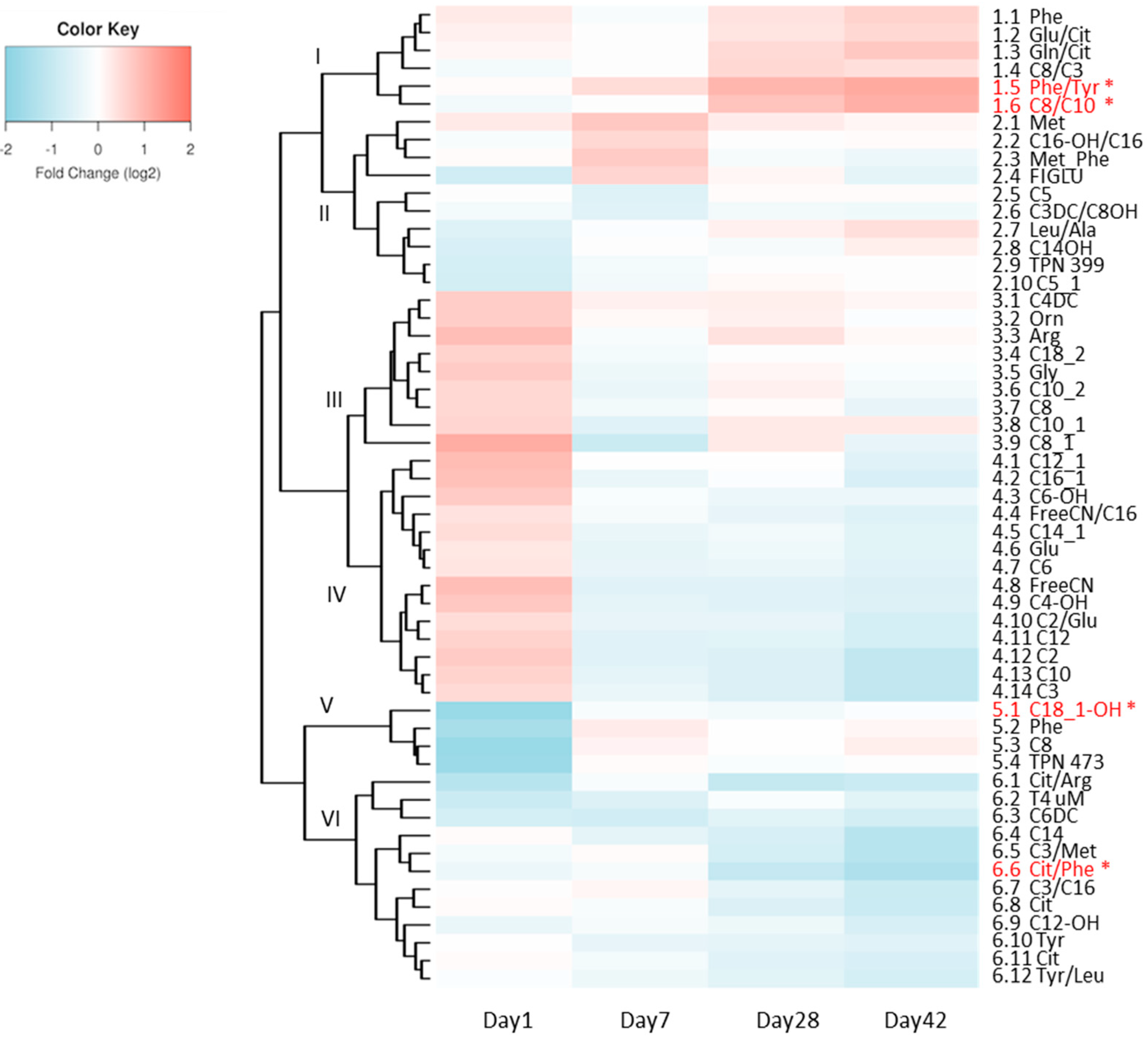
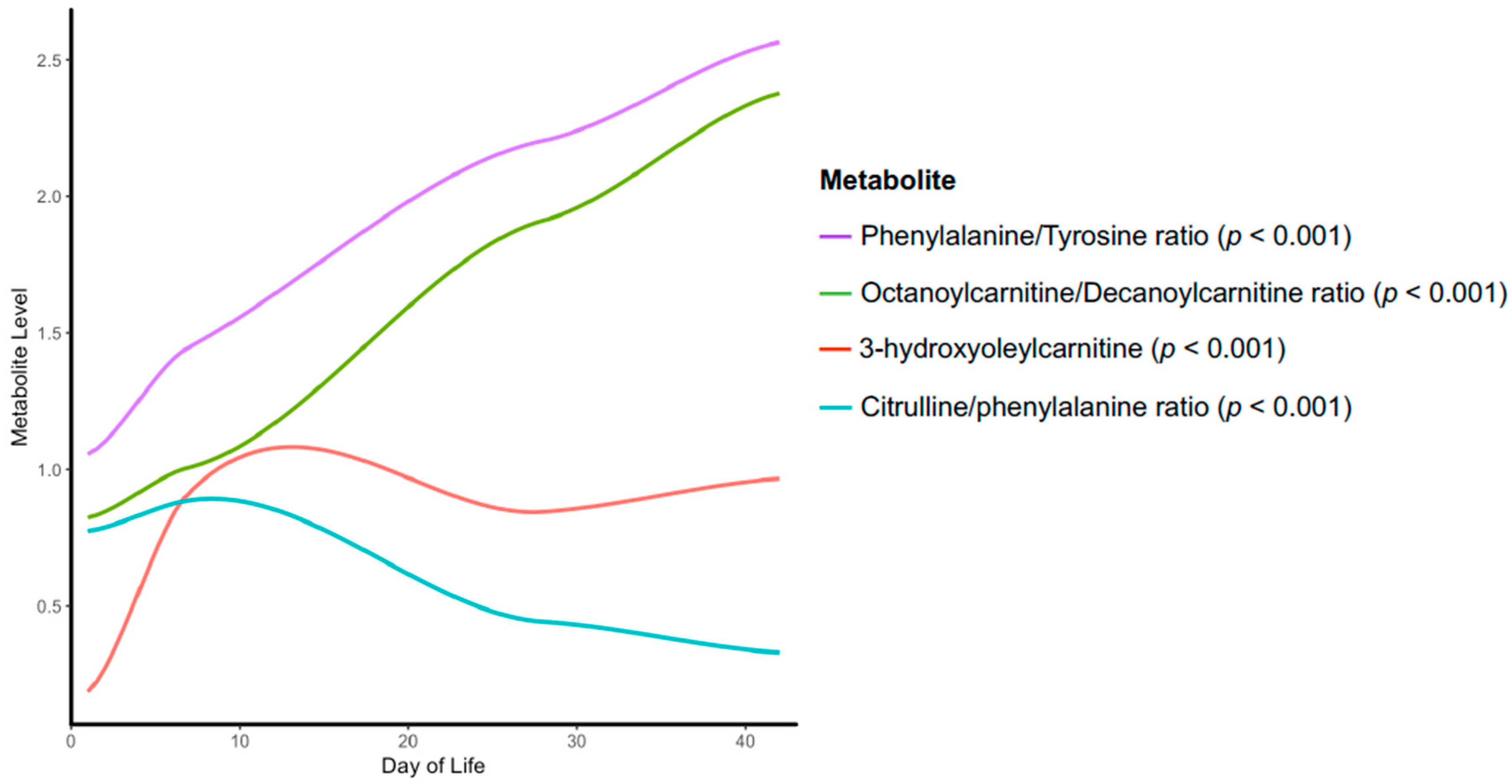
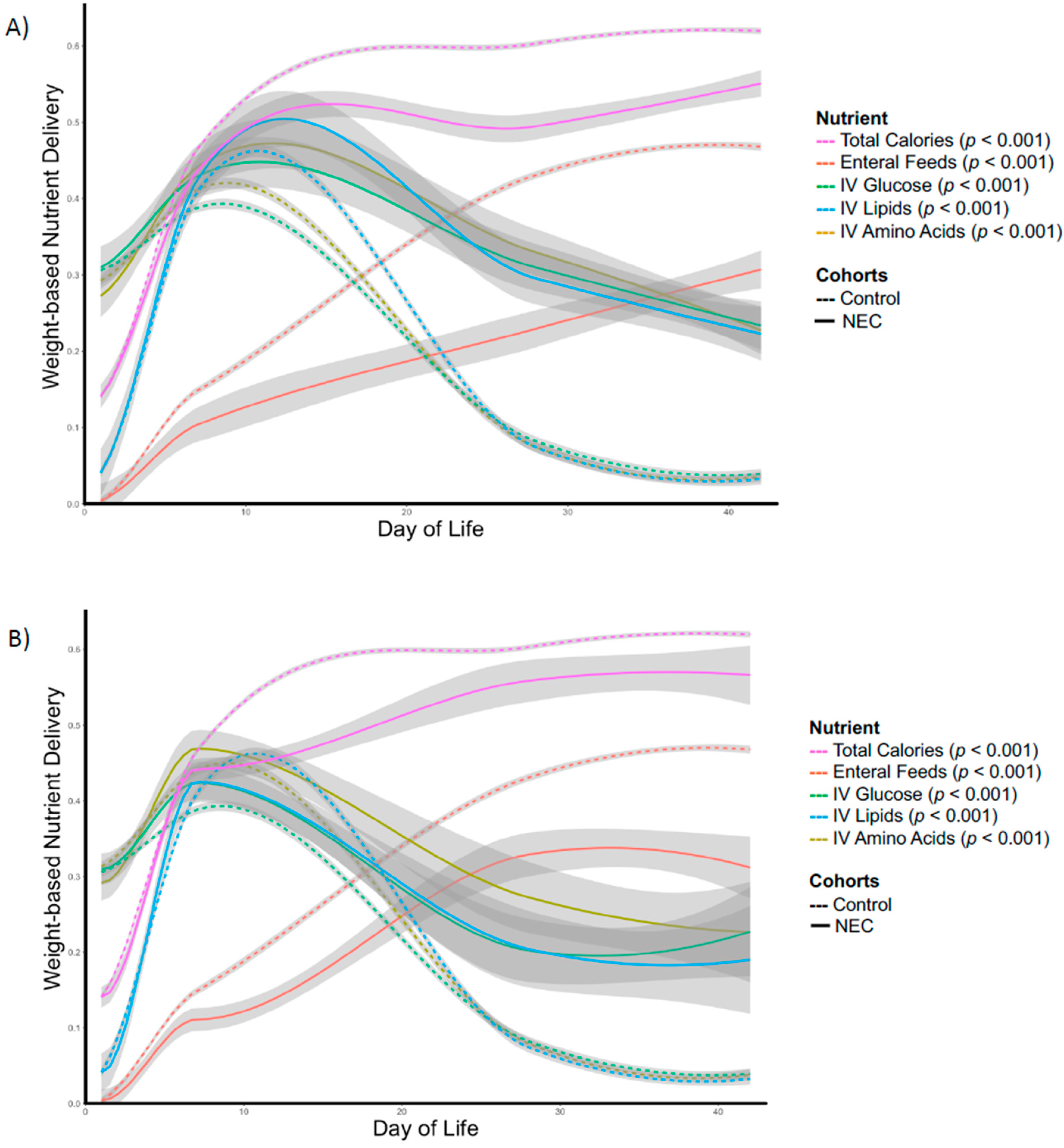
3. Results
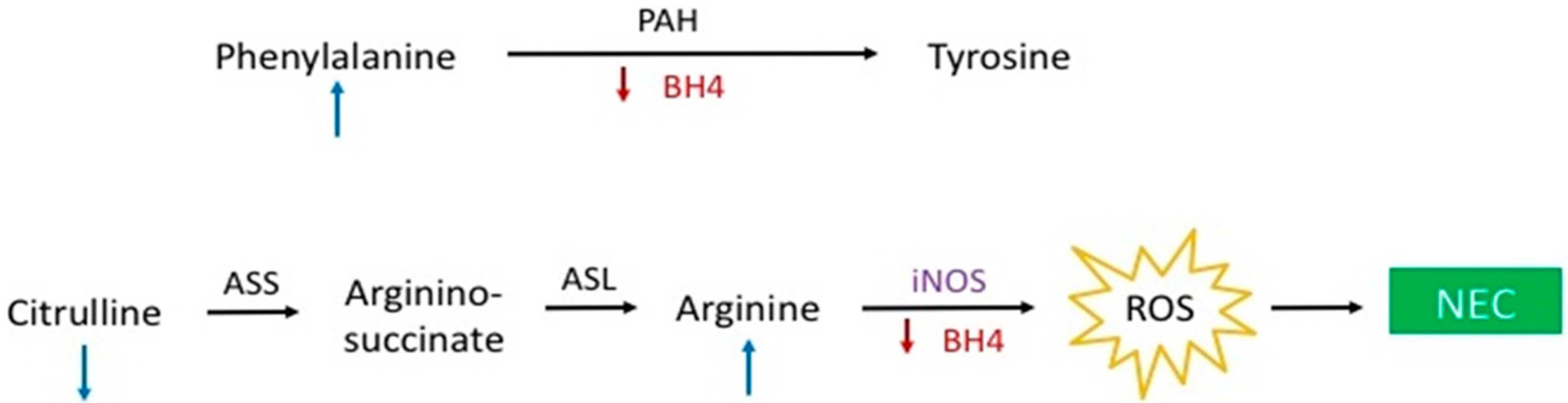
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meyburg, J. Acylcarnitine Profiles of Preterm Infants Over the First Four Weeks of Life. Pediatr. Res. 2002, 52, 720–723. [Google Scholar] [CrossRef]
- Oladipo, O.O.; Weindel, A.L.; Saunders, A.N.; Dietzen, D. Impact of premature birth and critical illness on neonatal range of plasma amino acid concentrations determined by LC-MS/MS. Mol. Genet. Metab. 2011, 104, 476–479. [Google Scholar] [CrossRef]
- Autmizguine, J.; Hornik, C.P.; Benjamin, D.K.; Laughon, M.M.; Clark, R.H.; Cotten, C.M.; Cohen-Wolkowiez, M.; Smith, P.B. Anaerobic Antimicrobial Therapy After Necrotizing Enterocolitis in VLBW Infants. Pediatrics 2014, 135, e117–e125. [Google Scholar] [CrossRef] [Green Version]
- Blanco, C.L.; Gong, A.K.; Green, B.K.; Falck, A.; Schoolfield, J.; Liechty, E.A. Early Changes in Plasma Amino Acid Concentrations during Aggressive Nutritional Therapy in Extremely Low Birth Weight Infants. J. Pediatr. 2011, 158, 543–548.e1. [Google Scholar] [CrossRef]
- Meyburg, J.; Schulze, A.; Kohlmueller, D.; Linderkamp, O.; Mayatepek, E. Postnatal Changes in Neonatal Acylcarnitine Profile. Pediatr. Res. 2001, 49, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Alul, F.Y.; Cook, D.E.; A Shchelochkov, O.; Fleener, L.G.; Berberich, S.L.; Murray, J.C.; Ryckman, K.K. The heritability of metabolic profiles in newborn twins. Heredity 2012, 110, 253–258. [Google Scholar] [CrossRef] [Green Version]
- McCallie, K.R.; Lee, H.C.; Mayer, O.; Cohen, R.S.; Hintz, S.R.; Rhine, W.D. Improved outcomes with a standardized feeding protocol for very low birth weight infants. J. Perinatol. 2011, 31, S61–S67. [Google Scholar] [CrossRef]
- Ehrenkranz, R.A.; Das, A.; Wrage, L.A.; Poindexter, B.B.; Higgins, R.D.; Stoll, B.J.; Oh, W.; Eunice Kennedy Shriver National Institute of Child Health and Human Development Neonatal Research Network. Early nutrition mediates the influence of severity of illness on extremely LBW infants. Pediatr. Res. 2011, 69, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Alvarez, R.; Almeida, A.; Medina, J.M. Oxidative Stress in Preterm Rat Brain Is Due to Mitochondrial Dysfunction. Pediatr. Res. 2002, 51, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.O.; Pinto, J.; Barros, A.; Morais, E.; Duarte, D.; Negrão, F.; Pita, C.; Almeida, M.D.C.; Carreira, I.M.; Spraul, M.; et al. Newborn Urinary Metabolic Signatures of Prematurity and Other Disorders: A Case Control Study. J. Proteome Res. 2015, 15, 311–325. [Google Scholar] [CrossRef] [Green Version]
- Baregamian, N.; Song, J.; Bailey, C.E.; Papaconstantinou, J.; Evers, B.M.; Chung, D.H. Tumor necrosis factor-α and apoptosis signal-regulating kinase 1 control reactive oxygen species release, mitochondrial autophagy and c-Jun N-terminal kinase/p38 phosphorylation during necrotizing enterocolitis. Oxidative Med. Cell. Longev. 2009, 2, 297–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrazi, A.; Branca, M.F.; Sodhi, C.P.; Good, M.; Yamaguchi, Y.; Egan, C.E.; Lu, P.; Jia, H.; Shaffiey, S.; Lin, J.; et al. Toll-like Receptor 4-mediated Endoplasmic Reticulum Stress in Intestinal Crypts Induces Necrotizing Enterocolitis*. J. Boil. Chem. 2014, 289, 9584–9599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Good, M.; Sodhi, C.P.; Yamaguchi, Y.; Jia, H.; Lu, P.; Fulton, W.B.; Martin, L.Y.; Prindle, T.; Nino, D.F.; Zhou, Q.; et al. The human milk oligosaccharide 2′-fucosyllactose attenuates the severity of experimental necrotising enterocolitis by enhancing mesenteric perfusion in the neonatal intestine. Br. J. Nutr. 2016, 116, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylvester, K.G.; Kastenberg, Z.J.; Moss, R.L.; Enns, G.M.; Cowan, T.M.; Shaw, G.M.; Stevenson, D.K.; Sinclair, T.J.; Scharfe, C.; Ryckman, K.K.; et al. Acylcarnitine Profiles Reflect Metabolic Vulnerability for Necrotizing Enterocolitis in Newborns Born Premature. J. Pediatr. 2016, 181, 80–85.e1. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.H.; Kelleher, A.S.; Chace, D.H.; Spitzer, A.R. Gestational Age and Age at Sampling Influence Metabolic Profiles in Premature Infants. Pediatrics 2014, 134, e37–e46. [Google Scholar] [CrossRef] [Green Version]
- Horbar, J.D.; Ehrenkranz, R.A.; Badger, G.J.; Edwards, E.M.; Morrow, K.A.; Soll, R.F.; Buzas, J.S.; Bertino, E.; Gagliardi, L.; Bellu’, R. Weight Growth Velocity and Postnatal Growth Failure in Infants 501 to 1500 Grams: 2000–2013. Pediatrics 2015, 136, e84–e92. [Google Scholar] [CrossRef] [Green Version]
- Patel, V.B.; Engstrom, J.L.; Meier, P.P.; Kimura, R.E. Accuracy of Methods for Calculating Postnatal Growth Velocity for Extremely Low Birth Weight Infants. Pediatrics 2005, 116, 1466–1473. [Google Scholar] [CrossRef]
- Zamora, S.A.; Amin, H.J.; McMillan, D.D.; Kubes, P.; Fick, G.H.; Butzner, J.; Parsons, H.G.; Scott, R. Plasma L-arginine concentrations in premature infants with necrotizing enterocolitis. J. Pediatr. 1997, 131, 226–232. [Google Scholar] [CrossRef]
- Becker, R.M.; Wu, G.; Galanko, J.A.; Chen, W.; Maynor, A.R.; Bose, C.L.; Rhoads, J. Reduced serum amino acid concentrations in infants with necrotizing enterocolitis. J. Pediatr. 2000, 137, 785–793. [Google Scholar] [CrossRef]
- Grishin, A.V.; Bowling, J.; Bell, B.; Wang, J.; Ford, H.R. Roles of nitric oxide and intestinal microbiota in the pathogenesis of necrotizing enterocolitis. J. Pediatr. Surg. 2015, 51, 13–17. [Google Scholar] [CrossRef]
- Ploder, M.; Neurauter, G.; Spittler, A.; Schroecksnadel, K.; Roth, E.; Fuchs, D. Serum phenylalanine in patients post trauma and with sepsis correlate to neopterin concentrations. Amino Acids 2007, 35, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Weinmann, A.; Post, M.; Pan, J.; Rafii, M.; O’Connor, D.L.; Unger, S.; Pencharz, P.; Belik, J. Tetrahydrobiopterin Is Present in High Quantity in Human Milk and Has a Vasorelaxing Effect on Newborn Rat Mesenteric Arteries. Pediatr. Res. 2011, 69, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Ehrenkranz, R.A.; Dusick, A.M.; Vohr, B.; Wright, L.L.; Wrage, L.A.; Poole, W.K. Growth in the Neonatal Intensive Care Unit Influences Neurodevelopmental and Growth Outcomes of Extremely Low Birth Weight Infants. Pediatrics 2006, 117, 1253–1261. [Google Scholar] [CrossRef] [Green Version]
- Ehrenkranz, R.A.; Younes, N.; A Lemons, J.; A Fanaroff, A.; Donovan, E.F.; Wright, L.L.; Katsikiotis, V.; Tyson, J.E.; Oh, W.; Shankaran, S.; et al. Longitudinal growth of hospitalized very low birth weight infants. Pediatrics 1999, 104, 280–289. [Google Scholar] [CrossRef]
- Vanderveen, D.K.; Martin, C.R.; Mehendale, R.; Allred, E.N.; Dammann, O.; Leviton, A. Early Nutrition and Weight Gain in Preterm Newborns and the Risk of Retinopathy of Prematurity. PLoS ONE 2013, 8, e64325. [Google Scholar] [CrossRef]
- Sjöström, E.S.; Lundgren, P.; Öhlund, I.; Holmström, G.; Hellström, A.; Domellöf, M. Low energy intake during the first 4 weeks of life increases the risk for severe retinopathy of prematurity in extremely preterm infants. Arch. Dis. Child.-Fetal Neonatal Ed. 2015, 101, F108–F113. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day Post-Delivery | Metabolite | OR | 95% CI | p-Value |
|---|---|---|---|---|
| Day 1 | Alanine | 1.002 | 1.0004–1.004 | 0.01 |
| Phenylalanine | 1.01 | 1.001–1.01 | 0.01 | |
| Free Carnitine | 1.02 | 1.002–1.04 | 0.03 | |
| C16 | 1.60 | 1.04–2.44 | 0.03 | |
| Arginine | 1.01 | 1.001–1.02 | 0.03 | |
| C14:1/C16 | 0.87 | 0.75–0.98 | 0.04 | |
| Citrulline/Phenylalanine | 0.63 | 0.40–0.96 | 0.04 | |
| Day 7 | Methionine/Phenylalanine | 1.07 | 1.01–1.12 | 0.01 |
| Methionine | 1.01 | 1.001–1.02 | 0.02 | |
| Citrulline/Arginine | 0.92 | 0.84–0.98 | 0.02 | |
| Arginine | 1.04 | 1.003–1.07 | 0.03 | |
| C12-DC | 0.85 | 0.73–0.98 | 0.03 | |
| Leucine/Alanine | 0.88 | 0.78–0.99 | 0.04 | |
| Day 28 | C5/C4 | 1.23 | 1.06–1.43 | 0.01 |
| C2 | 0.90 | 0.80–0.99 | 0.04 | |
| C4-OH | 0.27 | 0.07–0.85 | 0.04 | |
| Day 42 | C8/C16 | 1.93 | 1.14–3.41 | 0.02 |
| C4DC | 3.42 | 1.11–10.59 | 0.03 | |
| C10:1 | 1.12 | 1.01–1.25 | 0.03 | |
| C12-OH | 2.48 | 1.04–6.39 | 0.4 | |
| C10:2 | 1.54 | 1.01–2.40 | 0.04 |
| Characteristics | NEC * (n = 73) | Control (n = 814) | p-Value (NEC vs. Control) |
|---|---|---|---|
| Females, n (%) | 39 (53.4%) | 406 (49.9%) | 0.56 |
| Race, n (%) | 0.54 | ||
| Black | 27 (37.0%) | 197 (24.2%) | |
| White | 29 (39.7%) | 496(60.9%) | |
| Asian | 2 (2.7%) | 23 (2.8%) | |
| Other | 15 (20.5%) | 98 (12.0%) | |
| Birth weight (g), median (IQR) | 880 (709, 1180) | 1090 (860, 1303) | <0.001 |
| Gestational age (weeks), median (IQR) | 26 (25, 29) | 28 (27, 30) | <0.001 |
| Age at NEC diagnosis (days), median (IQR) | 20.5 (12.75, 28) | -- | -- |
| Apgar at 1 minute (minutes), mean (IGR) | 5.2 (4, 7) | 5.2 (3, 7) | 0.91 |
| Apgar at 5 minutes (minutes), mean (IQR) | 7.2 (6, 9) | 7.3 (7, 9) | 0.47 |
| Antenatal Steroids, n (%) | 65 (89.0%) | 758 (88.8%) | 0.94 |
| Transfused, n (%) | 66 (90.4%) | 462 (54.1%) | <0.001 |
| Postnatal steroids, n (%) | 20 (27.4%) | 92 (10.8%) | <0.001 |
| Late onset sepsis, n (%) | 23 (31.5%) | 88 (10.3%) | <0.001 |
| Retinopathy of prematurity, n (%) | 26 (36.6%) | 211 (24.9%) | 0.030 |
| Intraventricular hemorrhage, n (%) | 16 (21.9%) | 184 (21.5%) | 0.94 |
| Bronchopulmonary dysplasia, n (%) | 32 (43.8%) | 292 (34.2%) | 0.097 |
| Patent ductus ateriosus, n (%) | 35 (48.6%) | 343 (40.5%) | 0.18 |
| Day 1 | Day 7 | Day 28 | Day 42 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NEC | Control | p-Value | NEC | Control | p-Value | NEC | Control | p-Value | NEC | Control | p-Value | |
| Total Calories (kcal/kg/d) | 33.8 | 33.9 | 0.48 | 87.5 | 91.6 | 0.056 | 108.3 | 116.2 | 0.026 | 109.6 | 119.1 | 0.059 |
| IV Amino Acids (g/kg/d) | 1.7 | 1.8 | 0.077 | 2.7 | 2.6 | 0.14 | 1.6 | 0.5 | <0.001 | 1.3 | 0.2 | <0.001 |
| IV Lipids (g/kg/d) | 0.2 | 0.2 | 0.47 | 2.0 | 1.9 | 0.30 | 1.0 | 0.3 | 0.002 | 0.9 | 0.2 | <0.001 |
| IV Glucose (g/kg/d) | 7.2 | 7.1 | 0.35 | 9.9 | 9.1 | 0.069 | 4.7 | 1.9 | 0.003 | 5.3 | 0.0 | <0.001 |
| Enteral Feeds (mL/kg/d) | 1.1 | 1.1 | 0.42 | 33.7 | 45.0 | 0.025 | 100.0 | 131.0 | 0.003 | 94.8 | 142.2 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinclair, T.J.; Ye, C.; Chen, Y.; Zhang, D.; Li, T.; Ling, X.B.; Cohen, H.J.; Shaw, G.M.; Stevenson, D.K.; Chace, D.; et al. Progressive Metabolic Dysfunction and Nutritional Variability Precedes Necrotizing Enterocolitis. Nutrients 2020, 12, 1275. https://doi.org/10.3390/nu12051275
Sinclair TJ, Ye C, Chen Y, Zhang D, Li T, Ling XB, Cohen HJ, Shaw GM, Stevenson DK, Chace D, et al. Progressive Metabolic Dysfunction and Nutritional Variability Precedes Necrotizing Enterocolitis. Nutrients. 2020; 12(5):1275. https://doi.org/10.3390/nu12051275
Chicago/Turabian StyleSinclair, Tiffany J., Chengyin Ye, Yunliang Chen, Dongyan Zhang, Tian Li, Xuefeng Bruce Ling, Harvey J. Cohen, Gary M. Shaw, David K. Stevenson, Donald Chace, and et al. 2020. "Progressive Metabolic Dysfunction and Nutritional Variability Precedes Necrotizing Enterocolitis" Nutrients 12, no. 5: 1275. https://doi.org/10.3390/nu12051275
APA StyleSinclair, T. J., Ye, C., Chen, Y., Zhang, D., Li, T., Ling, X. B., Cohen, H. J., Shaw, G. M., Stevenson, D. K., Chace, D., Clark, R. H., & Sylvester, K. G. (2020). Progressive Metabolic Dysfunction and Nutritional Variability Precedes Necrotizing Enterocolitis. Nutrients, 12(5), 1275. https://doi.org/10.3390/nu12051275