The Effects of Aronia melanocarpa Extract on Testosterone-Induced Benign Prostatic Hyperplasia in Rats, and Quantitative Analysis of Major Constituents Depending on Extract Conditions

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. A. Melanocarpa Extraction
2.3. LC/MS Analysis of Aronia Extract
2.4. High Performance Liquid Chromatography Analysis of Phenolic Compounds
2.5. Animals
2.6. Induction of BPH and Treatment
- Group Sham: non-BPH-induced and received oral 0.5% CMC-Na
- Group VC: BPH-induced and received oral 0.5% CMC-Na
- Group PC: BPH-induced and received saw palmetto (100 mg/kg body weight, oral gavage)
- Group T1: BPH-induced and received 100% ethanol/low temperature (30 °C) A. melanocarpa extract (100 mg/kg body weight, oral gavage).
- Group T2: BPH-induced and received 100% ethanol/high temperature (100 °C) A. melanocarpa extract (100 mg/kg body weight, oral gavage).
- Group T3: BPH-induced and received 60% ethanol/low temperature (30 °C) A. melanocarpa extract (100 mg/kg body weight, oral gavage).
- Group T4: BPH-induced and received 60% ethanol/high temperature (100 °C) extracted A. melanocarpa (100 mg/kg body weight, oral gavage).
2.7. Tissue, Blood Collection, and Biochemical Analysis
2.8. Real-Time Reverse Transcriptase Polymerase Chain Reaction
2.9. Prostate Index (PI) and 5AR and DHT Expressions in Prostate Tissues
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effects of A. Melanocarpa Extracts on PI and Serum Liver Enzymes in the TP-Induced BPH Murine Model
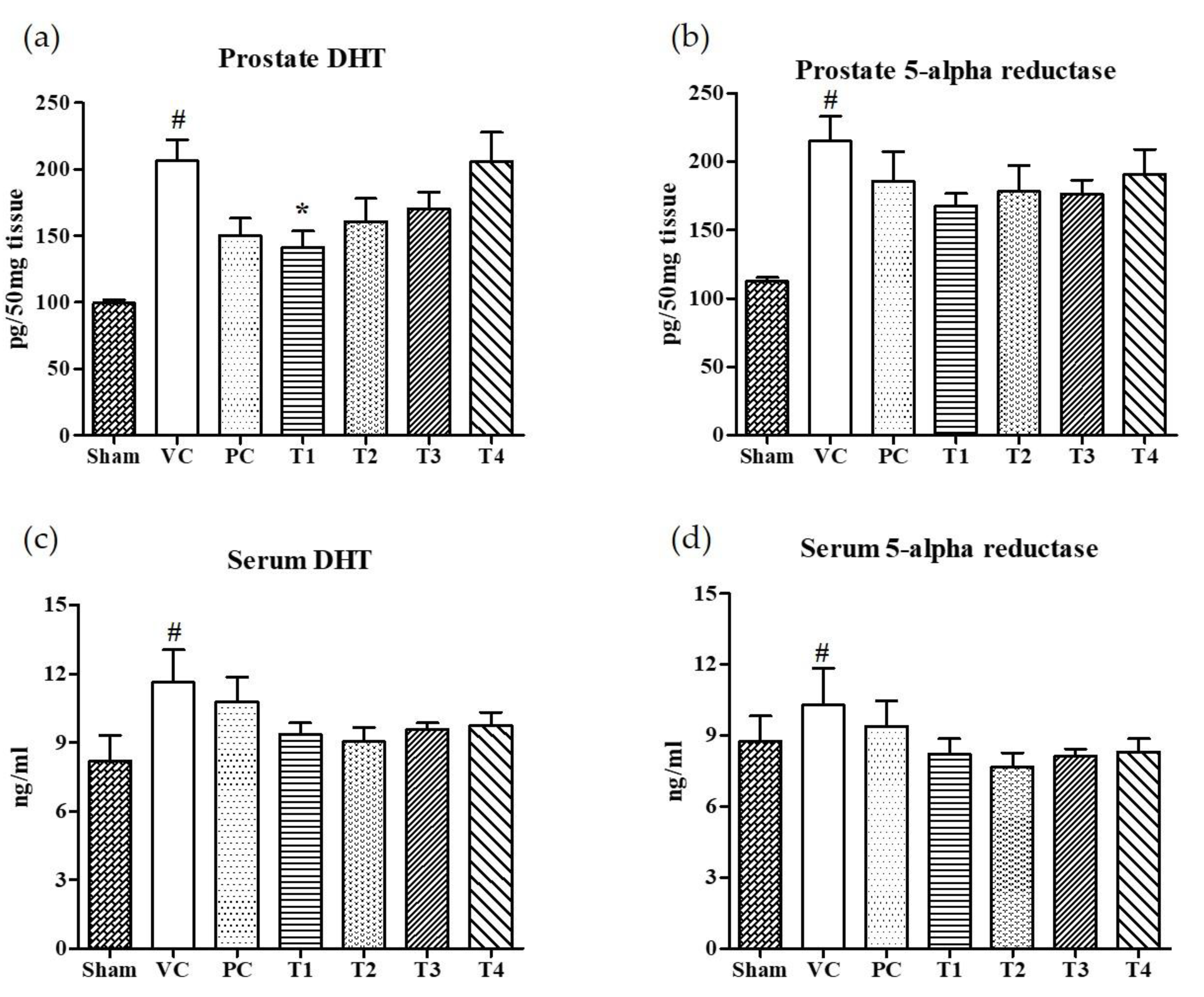
3.2. Effects of A. Melanocarpa Extracts on the Expression Levels of DHT and 5AR in Serum and Prostate Tissues
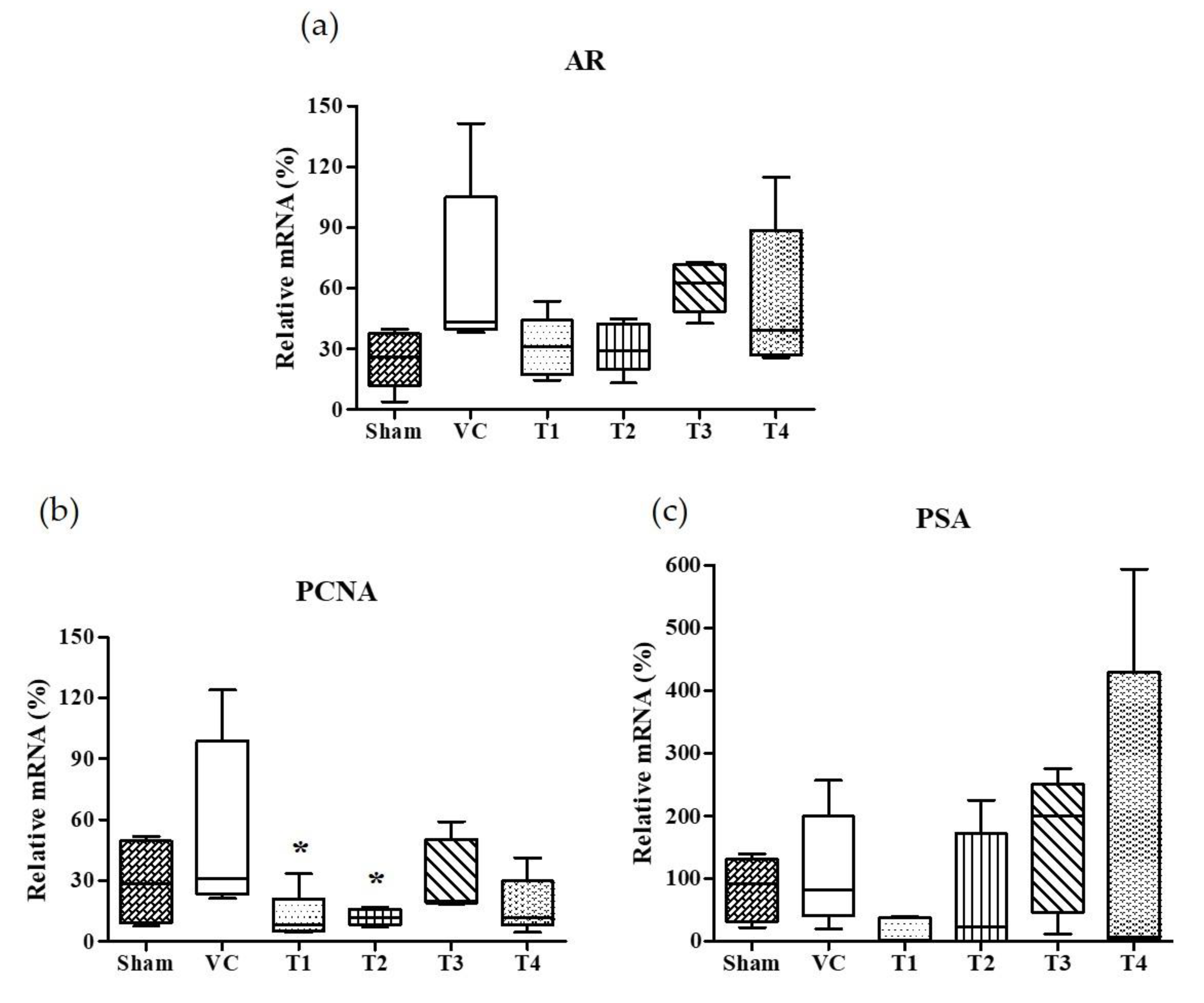
3.3. Effects of A. Melanocarpa Extracts on the mRNA Expressions of AR, PSA, and PCNA
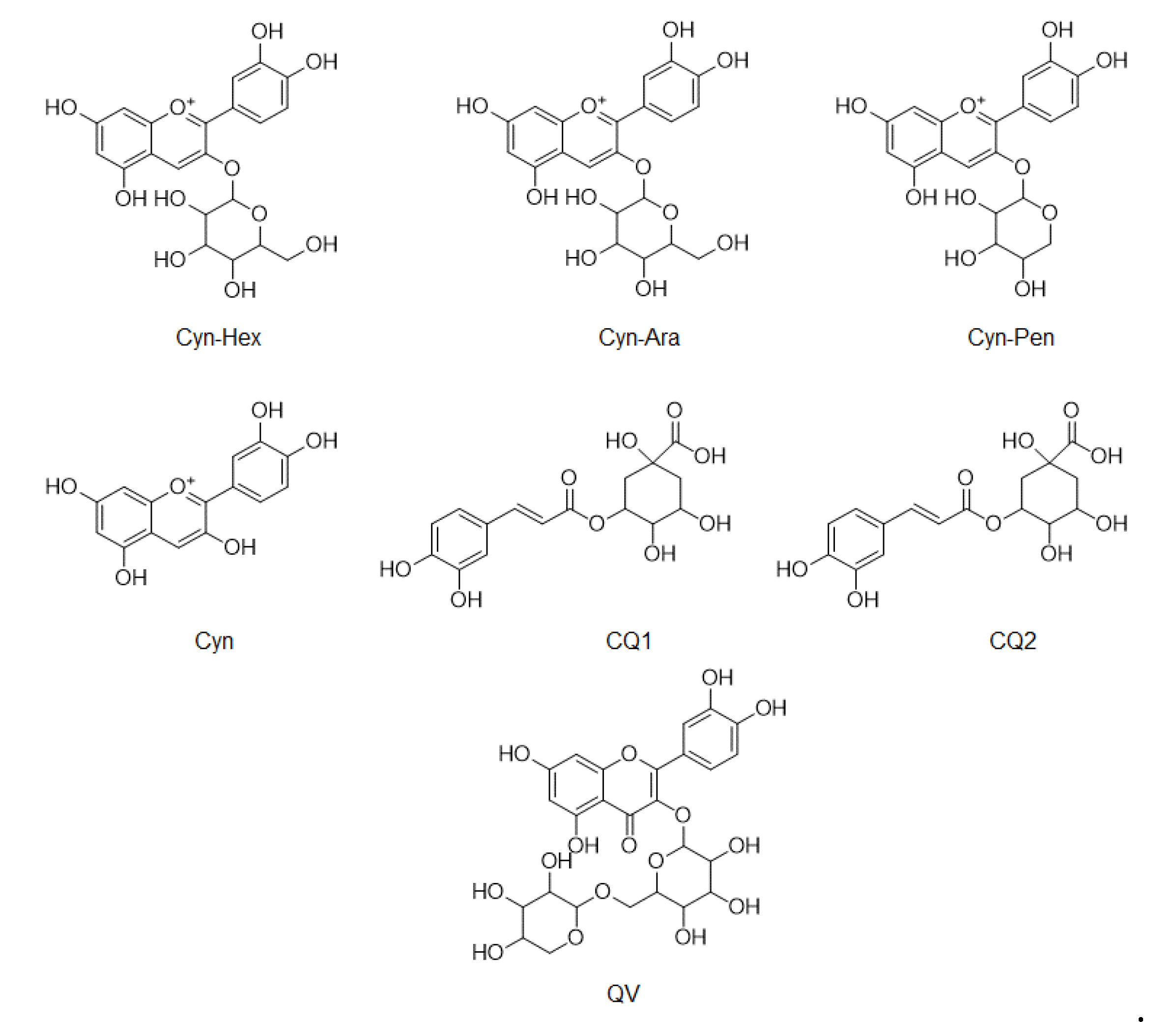
3.4. Comparison of Phenolic Compounds Abundance in A. Melanocarpa Extracts by LC/MS Analysis
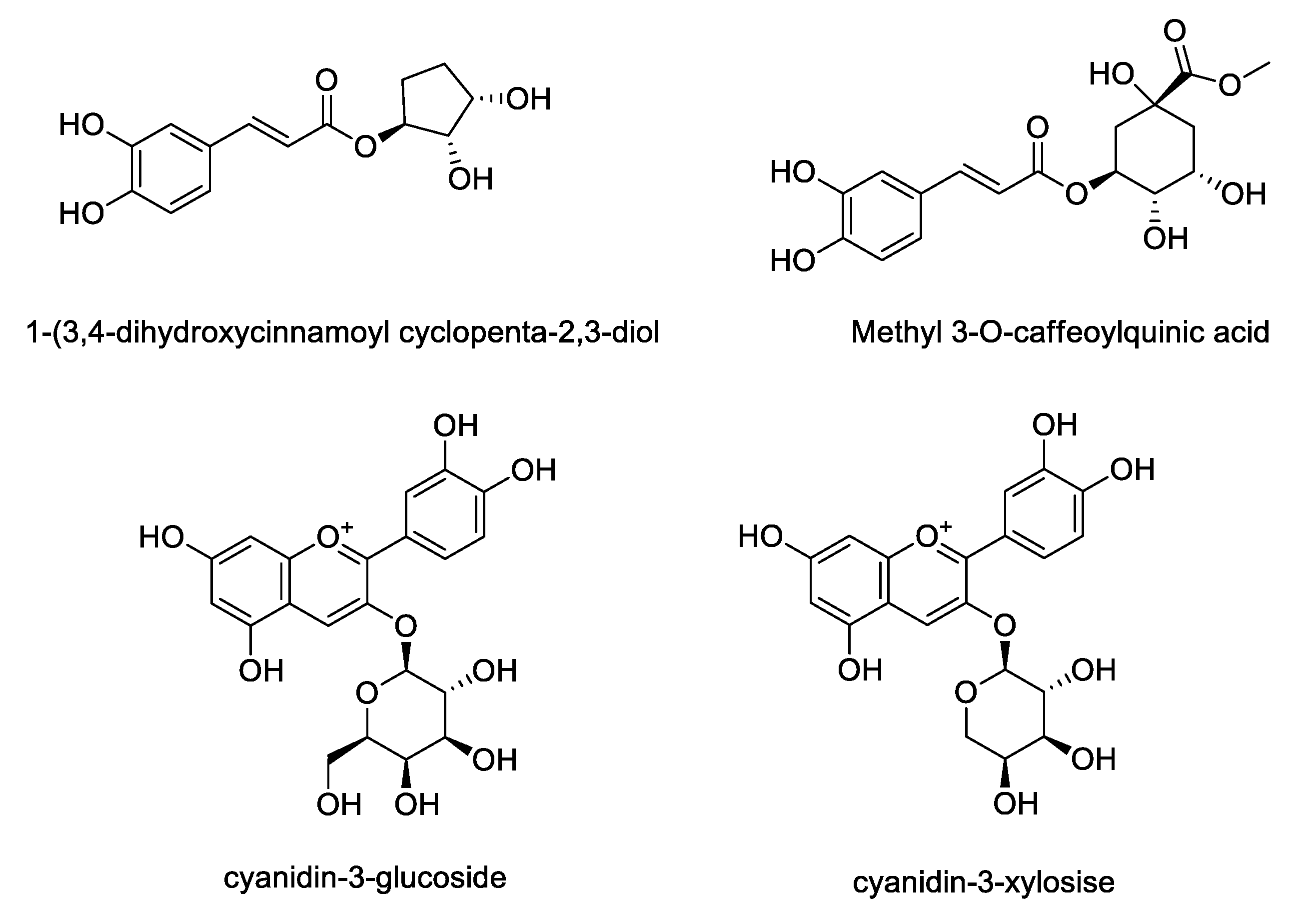
3.5. Quantification of Main Anthocyanin and Phenolics in A. Melanocarpa Extracts Using HPLC
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, A.; Neal, D. Benign prostatic hyperplasia. Lancet 2003, 361, 1359–1367. [Google Scholar] [CrossRef] [Green Version]
- McConnell, J.D.; Bruskewitz, R.; Walsh, P.; Andriole, G.; Lieber, M.; Holtgrewe, H.L.; Albertsen, P.; Roehrborn, C.G.; Nickel, J.C.; Wang, D.Z. The effect of finasteride on the risk of acute urinary retention and the need for surgical treatment among men with benign prostatic hyperplasia. N. Engl. J. Med. 1998, 338, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Penna, G.; Fibbi, B.; Amuchastegui, S.; Cossetti, C.; Aquilano, F.; Laverny, G.; Gacci, M.; Crescioli, C.; Maggi, M.; Adorini, L. Human benign prostatic hyperplasia stromal cells as inducers and targets of chronic immuno-mediated inflammation. J. Immunol. 2009, 182, 4056–4064. [Google Scholar] [CrossRef] [PubMed]
- Claus, S.; Berges, R.; Senge, T.; Schulze, H. Cell kinetic in epithelium and stroma of benign prostatic hyperplasia. J. Urol. 1997, 158, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Cunha, G.R.; Ricke, W.; Thomson, A.; Marker, P.C.; Risbridger, G.; Hayward, S.W.; Wang, Y.Z.; Donjacour, A.A.; Kurita, T. Hormonal, cellular, and molecular regulation of normal and neoplastic prostatic development. J. Steroid Biochem. Mol. Biol. 2004, 92, 221–236. [Google Scholar] [CrossRef]
- Andriole, G.; Bruchovsky, N.; Chung, L.W.; Matsumoto, A.M.; Rittmaster, R.; Roehrborn, C.; Russell, D.; Tindall, D. Dihydrotestosterone and the prostate: The scientific rationale for 5α-reductase inhibitors in the treatment of benign prostatic hyperplasia. J. Urol. 2004, 172, 1399–1403. [Google Scholar] [CrossRef]
- Gloyna, R.E.; Siiteri, P.K.; Wilson, J.D. Dihydrotestosterone in prostatic hypertrophy. II. The formation and content of dihydrotestosterone in the hypertrophic canine prostate and the effect of dihydrotestosterone on prostate growth in the dog. J. Clin. Investig. 1970, 49, 1746–1753. [Google Scholar] [CrossRef]
- Mooradian, A.D.; Morley, J.E.; Korenman, S.G. Biological actions of androgens. Endocr. Rev. 1987, 8, 1–28. [Google Scholar] [CrossRef]
- Carson, C.; Rittmaster, R. The role of dihydrotestosterone in benign prostatic hyperplasia. Urology 2003, 61, 2–7. [Google Scholar] [CrossRef]
- Wilson, J.D. The role of 5a-reduction in steroid hormone physiology. Reprod. Fertil. Dev. 2001, 13, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Rittmaster, R.S. 5α-Reductase inhibitors. J. Androl. 1997, 18, 582–587. [Google Scholar] [PubMed]
- Kokotkiewicz, A.; Jaremicz, Z.; Luczkiewicz, M. Aronia plants: A review of traditional use, biological activities, and perspectives for modern medicine. J. Med. Food 2010, 13, 255–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcheva-Kuzmanova, S.; Borisova, P.; Galunska, B.; Krasnaliev, I.; Belcheva, A. Hepatoprotective effect of the natural fruit juice from Aronia melanocarpa on carbon tetrachloride-induced acute liver damage in rats. Exp. Toxicol. Pathol. 2004, 56, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Valcheva-Kuzmanova, S.; Kuzmanov, K.; Mihova, V.; Krasnaliev, I.; Borisova, P.; Belcheva, A. Antihyperlipidemic effect of Aronia melanocarpa fruit juice in rats fed a high-cholesterol diet. Plant Foods Hum. Nutr. 2007, 62, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Naruszewicz, M.; Łaniewska, I.; Millo, B.; Dłużniewski, M. Combination therapy of statin with flavonoids rich extract from chokeberry fruits enhanced reduction in cardiovascular risk markers in patients after myocardial infraction (MI). Atherosclerosis 2007, 194, e179–e184. [Google Scholar] [CrossRef] [PubMed]
- Handeland, M.; Grude, N.; Torp, T.; Slimestad, R. Black chokeberry juice (Aronia melanocarpa) reduces incidences of urinary tract infection among nursing home residents in the long term-a pilot study. Nutr. Res. 2014, 34, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Kirakosyan, A.; Seymour, E.M.; Wolforth, J.; McNish, R.; Kaufman, P.B.; Bolling, S.F. Tissue bioavailability of anthocyanins from whole tart cherry in healthy rats. Food Chem. 2015, 171, 26–31. [Google Scholar] [CrossRef]
- Slimestad, R.; Torskangerpoll, K.; Nateland, H.S.; Johannessen, T.; Giske, N.H. Flavonoids from black chokeberries, Aronia melanocarpa. J. Food Comp. Anal. 2005, 18, 61–68. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Wrolstad, R.E. Anthocyanin pigments—Bioactivity and coloring properties. J. Food Sci. 2004, 69, 419–421. [Google Scholar] [CrossRef]
- Kırca, A.; Özkan, M.; Cemeroğlu, B. Effects of temperature, solid content and pH on the stability of black carrot anthocyanins. Food Chem. 2007, 101, 212–218. [Google Scholar] [CrossRef]
- Kırca, A.; Özkan, M.; Cemeroğlu, B. Stability of black carrot anthocyanins in various fruit juices and nectars. Food Chem. 2006, 97, 598–605. [Google Scholar] [CrossRef]
- Erlandson, J.A.; Wrolstad, R.E. Degradation of anthocyanins at limited water concentration. J. Food Sci. 1972, 37, 592–598. [Google Scholar] [CrossRef]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Stability of anthocyanin-based aqueous extracts of Andean purple corn and redfleshed sweet potato compared to synthetic and natural colorants. Food Chem. 2004, 86, 69–77. [Google Scholar] [CrossRef]
- Starr, M.S.; Francis, F.J. Oxygen and ascorbic acid effect on the relative stability of four anthocyanin pigments in cranberry juice. Food Technol. 1968, 22, 91–93. [Google Scholar]
- Tsai, P.J.; Hsieh, Y.Y.; Huang, T.C. Effect of sugar on anthocyanin degradation and water mobility in a roselle anthocyanin model system using 17O NMR. J. Agric. Food Chem. 2004, 52, 3097–3099. [Google Scholar] [CrossRef]
- Türkyılmaz, M.; Yemis, O.; Özkan, M. Clarification and pasteurisation effects on monomeric anthocyanins and percent polymeric colour of black carrot (Daucus carota L.) juice. Food Chem. 2012, 134, 1052–1058. [Google Scholar] [CrossRef]
- Molina, A.G.; Hernández-Martínez, R.A.; Cortez-Valadez, M.; García-Hernández, F.; Estevez, M. Effects of tetraethyl orthosilicate (teos) on the light and temperature stability of a pigment from Beta vulgaris and its potential food industry applications. Molecule 2014, 19, 17985–18002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, C.; Patel, A.V.; Calle, E.E.; Jacobs, E.J.; Chao, A.; Thun, M.J. Body mass index, height, and prostate cancer mortality in two large cohorts of adult men in the United States. Cancer Epidemiol. Biomark. Prev. 2001, 10, 345–353. [Google Scholar]
- Bent, S.; Kane, C.; Shinohara, K.; Neuhaus, J.; Hudes, E.S.; Goldberg, H.; Avins, A.L. Saw palmetto for benign prostatic hyperplasia. N. Engl. J. Med. 2006, 354, 557–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittmaster, R.S. 5α-reductase inhibitors in benign prostatic hyperplasia and prostate cancer risk reduction. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, G.; Rittmaster, R.S.; Klocker, H. Dihydrotestosterone and the concept of 5α-reductase inhibition in human benign prostatic hyperplasia. Eur. Urol. 2000, 37, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Gormley, G.J.; Stoner, E.; Bruskewitz, R.C.; Imperato-McGinley, J.; Walsh, P.C.; McConnell, J.D.; Andriole, G.L.; Geller, J.; Bracken, B.R.; Tenover, J.S. The effect of finasteride in men with benign prostatic hyperplasia. N. Engl. J. Med. 1992, 327, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Traish, A.M.; Hassani, J.; Guay, A.T.; Zitzmann, M.; Hansen, M.L. Adverse side effects of 5α-reductase inhibitors therapy: Persistent diminished libido and erectile dysfunction and depression in a subset of patients. J. Sex. Med. 2011, 8, 872–884. [Google Scholar] [CrossRef]
- Preuss, H.G.; Marcusen, C.; Regan, J.; Klimberg, I.W.; Welebir, T.A.; Jones, W.A. Randomized trial of a combination of natural products (cernitin, saw palmetto, B-sitosterol, vitamin E) on symptoms of benign prostatic hyperplasia (BPH). Int. Urol. Nephrol. 2001, 33, 217–225. [Google Scholar] [CrossRef]
- Steenkamp, V.; Gouws, M.C.; Gulumian, M.; Elgorashi, E.E.; Van Staden, J. Studies on antibacterial, anti-inflammatory and antioxidant activity of herbal remedies used in the treatment of benign prostatic hyperplasia and prostatitis. J. Ethnopharmacol. 2006, 103, 71–75. [Google Scholar] [CrossRef]
- Wadsworth, T.L.; Worstell, T.R.; Greenberg, N.M.; Roselli, C.E. Effects of dietary saw palmetto on the prostate of transgenic adenocarcinoma of the mouse prostate model (TRAMP). Prostate 2007, 67, 661–673. [Google Scholar] [CrossRef]
- Van Coppenolle, F.; Le Bourhis, X.; Carpentier, F.; Delaby, G.; Cousse, H.; Raynaud, J.P.; Dupouy, J.P.; Prevarskaya, N. Pharmacological effects of the lipidosterolic extract of Serenoa repens (Permixon) on rat prostate hyperplasia induced by hyperprolactinemia: Comparison with finasteride. Prostate 2000, 43, 49–58. [Google Scholar] [CrossRef]
- Talpur, N.; Echard, B.; Bagchi, D.; Bagchi, M.; Preuss, H.G. Comparison of Saw palmetto (extract and whole berry) and cernitin on prostate growth in rats. Mol. Cell. Biochem. 2003, 250, 21–26. [Google Scholar] [CrossRef]
- Olapade, E.O.; Olapade, E.O., Jr.; Olapade, C.O.; Olapade, O.C. Phytomedicines for the treatment of benign prostatic hyperplasia without surgery in Nigeria. Acta Hortic. 2003, 597, 231–234. [Google Scholar] [CrossRef]
- Gelmann, E.P. Molecular biology of the androgen receptor. J. Clin. Oncol. 2002, 20, 3001–3015. [Google Scholar] [CrossRef] [PubMed]
- Simental, J.A.; Sar, M.; Lane, M.V.; French, F.S.; Wilson, E.M. Transcriptional activation and nuclear targeting signals of the human androgen receptor. J. Biol. Chem. 1991, 266, 510–518. [Google Scholar] [PubMed]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev. 2004, 25, 276–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacs, J.T.; Isaacs, W.B. Androgen receptor outwits prostate cancer drugs. Nat. Med. 2004, 10, 26–27. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P. The nuclear receptor superfamily: The second decade. Cell 1995, 8, 835–839. [Google Scholar] [CrossRef] [Green Version]
- Tsui, K.H.; Wu, L.L.; Chang, P.L.; Hsieh, M.L.; Juang, H.H. Identifying the combination of the transcriptional regulatory sequences on prostate specific antigen and human glandular kallikrein genes. J. Urol. 2004, 172, 2029–2034. [Google Scholar] [CrossRef]
- Vernon, S.E.; Williams, W.D. Pre-treatment and post-treatment evaluation of prostatic adenocarcinoma for prostatic specific acid phosphatase and prostatic specific antigen by immunohistochemistry. J. Urol. 1983, 130, 95–98. [Google Scholar] [CrossRef]
- Hong, S.J.; Kim, S.I.; Kwon, S.M.; Lee, J.R.; Chung, B.C. Comparative study of concentration of isoflavones and lignans in plasma and prostatic tissues of normal control and benign prostatic hyperplasia. Yonsei Med. J. 2002, 43, 236–241. [Google Scholar] [CrossRef] [Green Version]
- Adhami, V.M.; Siddiqui, I.A.; Sarfaraz, S.; Khwaja, S.I.; Hafeez, B.B.; Ahmad, N.; Mukhtar, H. Effective prostate cancer chemopreventive intervention with green tea polyphenols in the TRAMP model depends on the stage of the disease. Clin. Cancer Res. 2009, 15, 1947–1953. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TP | ALB | BUN | CREA | AST | ALT | ALP | Bil | TG | TCHO | |
|---|---|---|---|---|---|---|---|---|---|---|
| Sham | 5.4 ± 0.1 | 3.3 ± 0.1 | 20.4 ± 0.5 | 0.50 ± 0.05 | 123.1 ± 10.4 | 34.3 ± 5.8 | 485.5 ± 68.9 | 0.16 ± 0.03 | 114.7 ± 33.1 | 73.7 ± 4.4 |
| VC | 6.3 ± 0.9 | 4.1 ± 0.5 | 25.2 ± 4.3 | 0.56 ± 0.10 | 134.2 ± 22.5 | 33.7 ± 7.9 | 530.8 ± 110.3 | 0.17 ± 0.03 | 99.9 ± 33.6 | 86.6 ± 18.4 |
| PC | 6.5 ± 0.6 | 4.1 ± 0.3 | 25.0 ± 2.0 | 0.51 ± 0.05 | 132.4 ± 11.5 | 36.5 ± 5.5 | 595.8 ± 128.4 | 0.14 ± 0.04 | 130.7 ± 51.2 | 87.8 ± 21.8 |
| T1 | 5.7 ± 0.1 | 3.8 ± 0.1 | 23.4 ± 2.6 | 0.52 ± 0.03 | 94.1 ± 8.7 * | 31.9 ± 2.8 | 517.4 ± 103.3 | 0.17 ± 0.01 | 79.0 ± 26.9 | 78.4 ± 6.6 |
| T2 | 5.7 ± 0.1 | 3.7 ± 0.1 | 21.4 ± 0.8 | 0.50 ± 0.09 | 100.6 ± 8.1 * | 34.0 ± 3.3 | 435.3 ± 91.8 | 0.16 ± 0.03 | 113.3 ± 57.3 | 83.0 ± 9.4 |
| T3 | 5.8 ± 0.2 | 3.8 ± 0.1 | 22.7 ± 2.2 | 0.52 ± 0.06 | 96.3 ± 8.0 * | 31.6 ± 3.6 | 469.2 ± 91.9 | 0.16 ± 0.02 | 123.9 ± 59.3 | 74.8 ± 12.7 |
| T4 | 5.6 ± 0.2 | 3.7 ± 0.2 | 20.1 ± 1.3 | 0.44 ± 0.03 | 82.5 ± 19.4 * | 32.6 ± 2.1 | 358.8 ± 37.1* | 0.18 ± 0.02 | 72.3 ± 19.9 | 73.8 ± 6.8 |
| Cyn-Hex | Cyn-Ara | Cyn-Pent | Cyn | CQ1 | CQ2 | QV | |
|---|---|---|---|---|---|---|---|
| (× 103) | |||||||
| T1 | 586 | 158 | 2.09 | 47.7 | 3.07 | 5.73 | 13.2 |
| T2 | 268 | 68 | 1.08 | 20.9 | 2.21 | 3.55 | 10 |
| T3 | 310 | 80.9 | 1.13 | 25.7 | 2.76 | 4.24 | 10.9 |
| T4 | 230 | 56.8 | 0.69 | 18.4 | 2.45 | 3.86 | 0.88 |
| Sample | Contents (mg/mL) | |||
|---|---|---|---|---|
| 1-(3, 4-Dihydroxycinnamoyl Cyclopenta-2, 3-dilo) | Methyl 3-O-Caffeoylquinic Acid | Cyanidin-3-Glucoside | Cyanidin-3-Xyloside | |
| T1 | 0.5349 | ND | 0.7642 | 2.6843 |
| T2 | 0.5461 | ND | 0.5107 | 2.3228 |
| T3 | 0.5015 | ND | 0.4559 | 2.1206 |
| T4 | 0.5583 | ND | 0.6002 | 2.4279 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.-H.; Jegal, J.; Kim, Y.N.; Heo, J.-D.; Rho, J.-R.; Yang, M.H.; Jeong, E.J. The Effects of Aronia melanocarpa Extract on Testosterone-Induced Benign Prostatic Hyperplasia in Rats, and Quantitative Analysis of Major Constituents Depending on Extract Conditions. Nutrients 2020, 12, 1575. https://doi.org/10.3390/nu12061575
Kim N-H, Jegal J, Kim YN, Heo J-D, Rho J-R, Yang MH, Jeong EJ. The Effects of Aronia melanocarpa Extract on Testosterone-Induced Benign Prostatic Hyperplasia in Rats, and Quantitative Analysis of Major Constituents Depending on Extract Conditions. Nutrients. 2020; 12(6):1575. https://doi.org/10.3390/nu12061575
Chicago/Turabian StyleKim, Na-Hyun, Jonghwan Jegal, Yun Na Kim, Jeong-Doo Heo, Jung-Rae Rho, Min Hye Yang, and Eun Ju Jeong. 2020. "The Effects of Aronia melanocarpa Extract on Testosterone-Induced Benign Prostatic Hyperplasia in Rats, and Quantitative Analysis of Major Constituents Depending on Extract Conditions" Nutrients 12, no. 6: 1575. https://doi.org/10.3390/nu12061575
APA StyleKim, N.-H., Jegal, J., Kim, Y. N., Heo, J.-D., Rho, J.-R., Yang, M. H., & Jeong, E. J. (2020). The Effects of Aronia melanocarpa Extract on Testosterone-Induced Benign Prostatic Hyperplasia in Rats, and Quantitative Analysis of Major Constituents Depending on Extract Conditions. Nutrients, 12(6), 1575. https://doi.org/10.3390/nu12061575