Effects of Ginseng Ingestion on Salivary Testosterone and DHEA Levels in Healthy Females: An Exploratory Study



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Study Design
2.3. Subject Recruitment
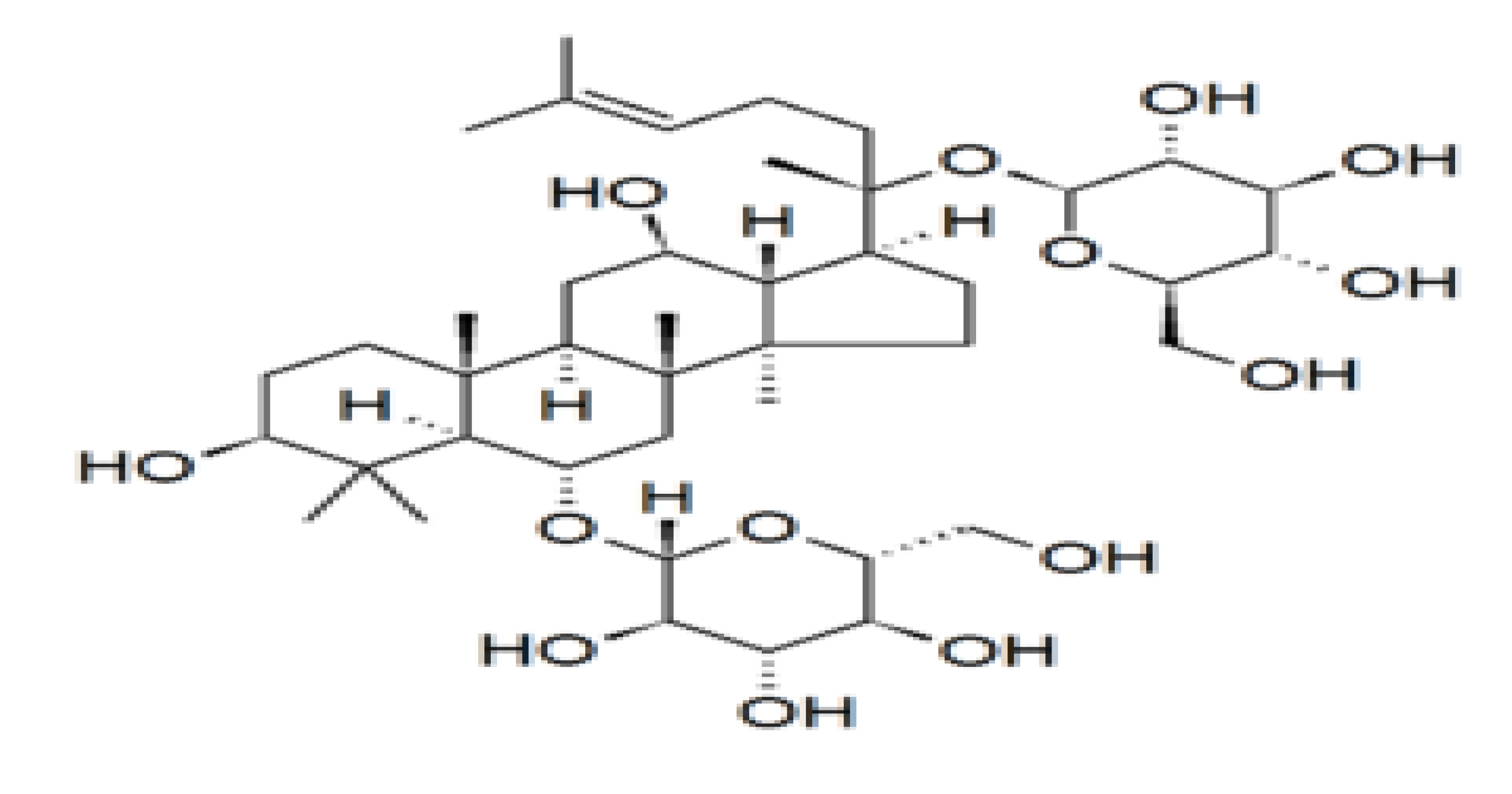
2.4. Ginseng Supplement
2.5. Study Protocol
2.6. Diet Diaries
2.7. Sample Preparation and Hormone Estimation
2.8. Statistical Analysis
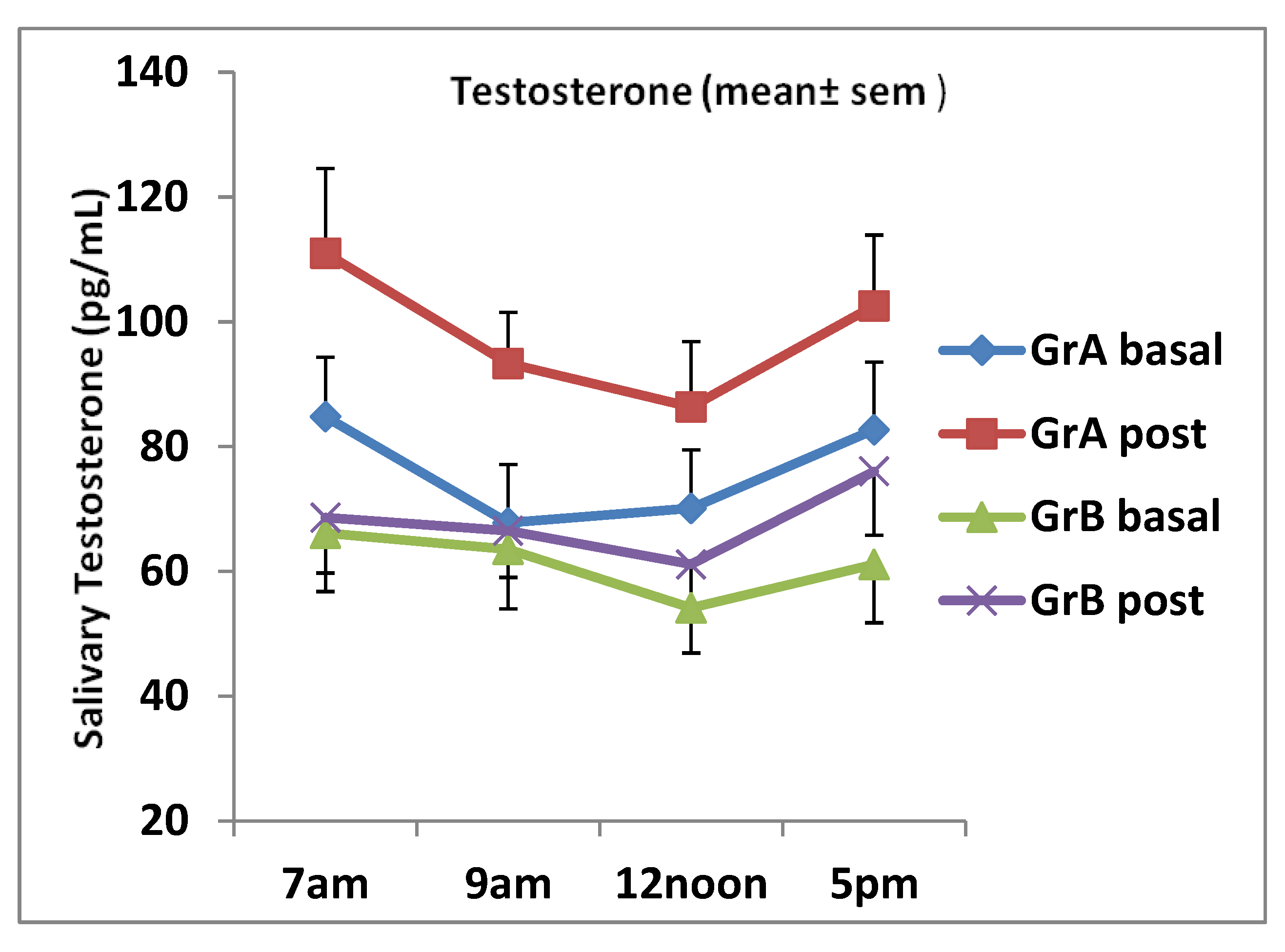
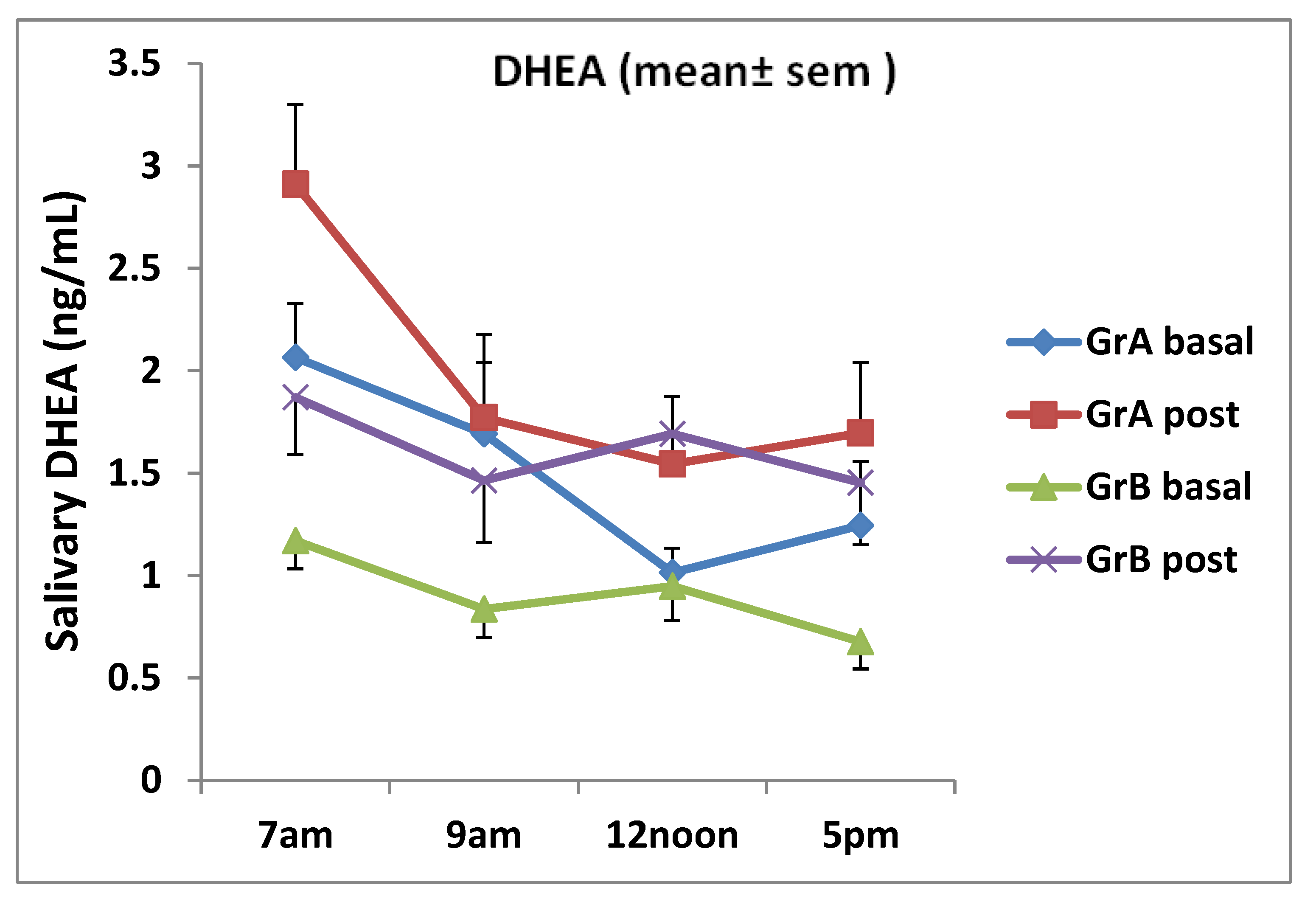
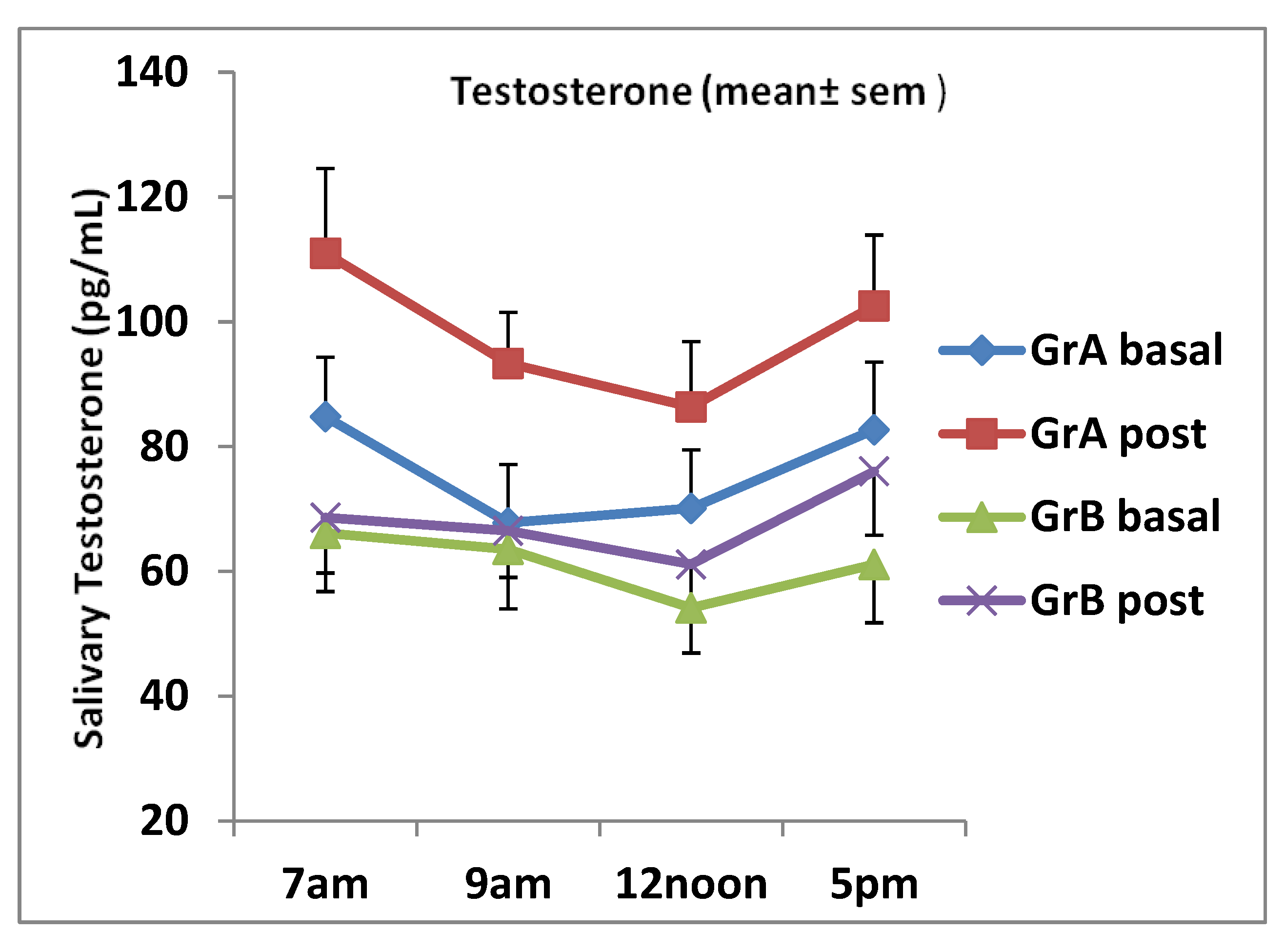
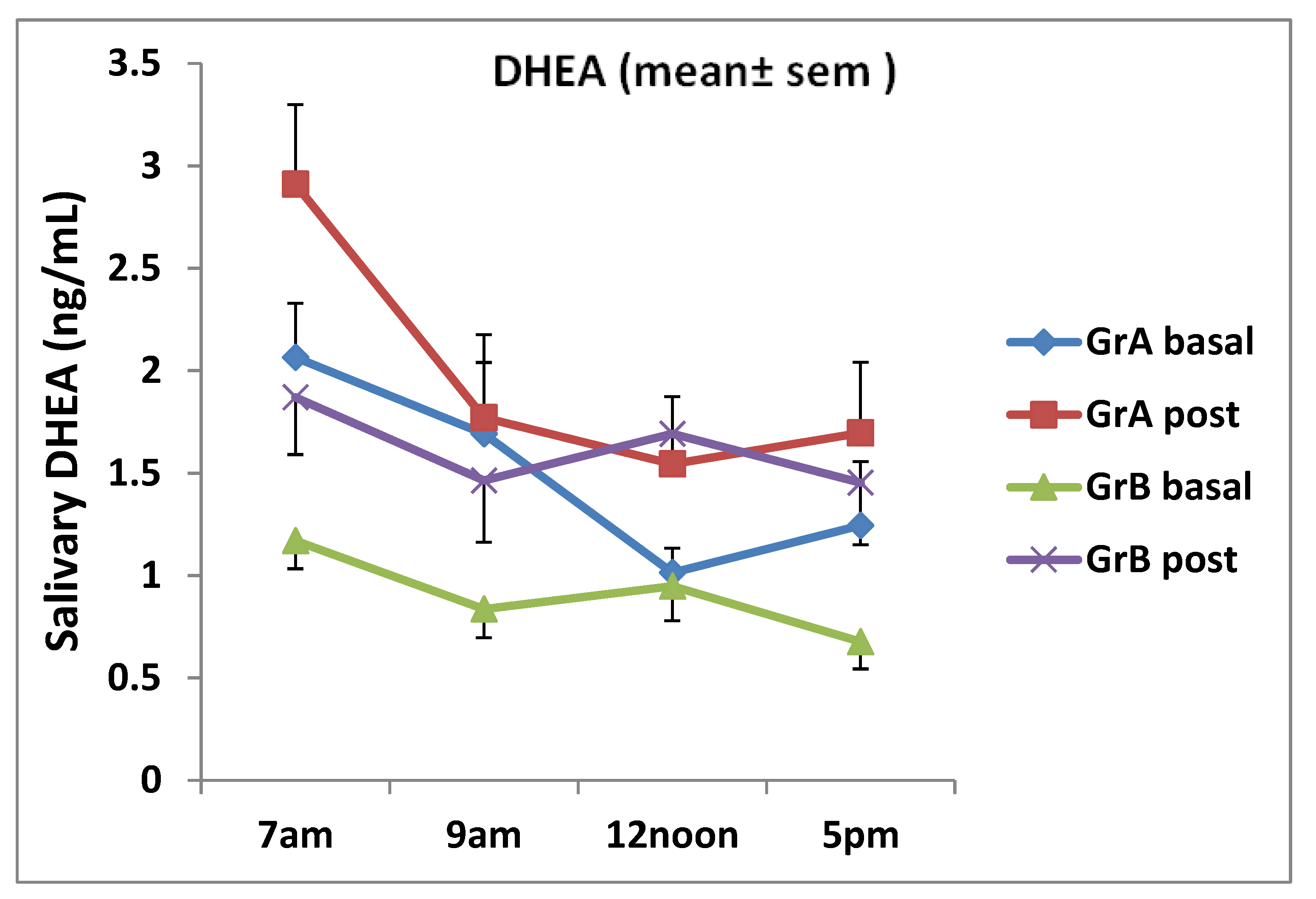
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTH | Adrocorticotrophic hormone |
| BMI | Body mass index |
| DHEA | Dehydroepiandrosterone |
| DHEAS | Dehydroepiandrosterone sulphate |
| ELISA | Enzyme-linked immunosorbent assay |
| SHBG | Sex hormone binding globulin |
| SPSS | Statistical Package for the Social Sciences |
| Testo | Testosterone |
References
- Li, C.P.; Li, R.C. An Introductory Note to Ginseng. Am. J. Chin. Med. 1973, 1, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Agarwal, S.S.; Gupta, B.M. Immunomodulatory activity of Panax ginseng extract. Planta Medica 1984, 50, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Scaglione, F.; Ferrara, F.; Dugnani, S.; Falchi, M.; Santoro, G.; Fraschini, F. Immunomodulatory effects of two extracts of Panax ginseng CA Meyer. Drugs Exp. Clin. Res. 1990, 16, 537–542. [Google Scholar] [PubMed]
- Kim, D.H. Chemical diversity of Panax ginseng, Panax quinquifolium, and Panax notoginseng. J. Ginseng Res. 2012, 36, 1. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Kim, N.H.; Park, S.J.; Kim, J.M.; Ryu, J.H. Ginseng in traditional herbal prescriptions. J. Ginseng Res. 2012, 36, 225. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Song, H.; Kim, S.-K.; Lee, M.S.; Rhee, D.K.; Lee, Y. Effects of ginseng on two main sex steroid hormone receptors: Estrogen and androgen receptors. J. Ginseng Res. 2016, 41, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Min, H. Ginseng, the ‘immunity boost’: The effects of Panax ginseng on immune system. J. Ginseng Res. 2012, 36, 354. [Google Scholar] [CrossRef] [Green Version]
- Shibata, S.; Tanaka, O.; Soma, K.; Aando, T.; Iida, Y.; Nakamura, H. Studies on saponins and sapogenins of ginseng. The structure of panaxatriol. Tetrahedron Lett. 1965, 42, 207–213. [Google Scholar] [CrossRef]
- Attele, A.S.; Wu, J.A.; Yuan, C.S. Ginseng pharmacology: Multiple constituents and multiple actions. Biochem. Pharmacol. 1999, 58, 1685–1693. [Google Scholar] [CrossRef]
- Harkey, M.R.; Henderson, G.L.; Gershwin, M.E.; Stern, J.S.; Hackman, R.M. Variability in commercial ginseng products: An analysis of 25 preparations. Am. J. Clin. Nutr. 2001, 73, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Park, C.S.; Yoo, M.H.; Noh, K.-H.; Oh, D.K. Biotransformation of ginsenosides by hydrolyzing the sugar moieties of ginsenosides using microbial glycosidases. Appl. Microbiol. Biotechnol. 2010, 87, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.; Lee, K.Y.; Lee, Y.; Lee, Y.H.; Lee, S.K. Ginsenoside-Rg1 down-regulates glucocorticoid receptor and displays synergistic effects with cAMP. Steroids 1998, 63, 421–424. [Google Scholar] [CrossRef]
- Wu, J.; Pan, Z.; Wang, Z.; Zhu, W.; Shen, Y.; Cui, R.; Lin, J.; Yu, H.; Wang, Q.; Yu, Y.; et al. Ginsenoside Rg1 protection against β-amyloid peptide-induced neuronal apoptosis via estrogen receptor α and glucocorticoid receptor-dependent anti-protein nitration pathway. Neuropharmacology 2012, 63, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; Leung, S.W.; Ji, L.; Huang, Y.; Hou, M.; Xu, A.; Wang, Z.; Vanhoutte, P.M. Notoginsenoside Ft1 activates both glucocorticoid and estrogen receptors to induce endothelium-dependent, nitric oxide-mediated relaxations in rat mesenteric arteries. Biochem. Pharmacol. 2014, 88, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-S.; Park, H.S.; Park, J.W.; Li, S.H.; Chun, Y.-S. Red ginseng and 20 (S)-Rg3 control testosterone-induced prostate hyperplasia by deregulating androgen receptor signaling. J. Nat. Med. 2011, 66, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, H.; Rayburn, E.R.; Zhao, Y.; Hill, D.L.; Zhang, R. 20-(S)-25-methoxyl-dammarane-3β, 12β, 20-triol, a novel natural product for prostate cancer therapy: Activity in vitro and in vivo and mechanisms of action. Br. J. Cancer 2008, 98, 792–802. [Google Scholar] [CrossRef] [Green Version]
- Park, G.H.; Park, K.Y.; Cho, H.-I.; Lee, S.M.; Han, J.S.; Won, C.H.; Shin, H. Red ginseng extract promotes the hair growth in cultured human hair follicles. J. Med. Food 2015, 18, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.H.; Kim, I.H.; Lee, M.J.; Nguyen, C.T.; Ha, J.A.; Lee, S.C.; Choi, S.; Choi, K.T.; Pyo, S.; Rhee, D.K. Anti-oxidative stress effect of red ginseng in the brain is mediated by peptidyl arginine deiminase type IV (PADI4) repression via estrogen receptor (ER) β up-regulation. J. Ethnopharmacol. 2013, 148, 474–485. [Google Scholar] [CrossRef]
- Ding, J.; Xu, Y.; Ma, P.; Jinna, A.; Yang, X.; Liu, Z.; Lin, N. Estrogenic effect of the extract of Renshen (Radix Ginseng) on reproductive tissues in immature mice. J. Tradit. Chin. Med. 2015, 35, 460–467. [Google Scholar]
- Xu, Y.; Ding, J.; Ma, X.-P.; Ma, Y.-H.; Liu, Z.Q.; Lin, N. Treatment with Panax ginseng antagonizes the estrogen decline in ovariectomized mice. Int. J. Mol. Sci. 2014, 15, 7827–7840. [Google Scholar] [CrossRef] [Green Version]
- Shim, M.K.; Lee, Y. Estrogen Receptor Is Activated by Korean Red Ginseng In Vitro but Not In Vivo. J. Ginseng Res. 2012, 36, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, H.G. Androgen production in women. Fertil. Steril. 2002, 77, 3–5. [Google Scholar] [CrossRef]
- Al-Dujaili, E.A.S.; Kenyon, C.J.; Nicol, M.R.; Mason, J.I. Liquorice and glycyrrhetinic acid increase DHEA and deoxycorticosterone levels in vivo and in vitro by inhibiting adrenal SULT2A1 activity. Mol. Cell. Endocrinol. 2011, 336, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Labrie, F.; Luu-The, V.; Lin, S.-X.; Simard, J.; Pelletier, G.; Labrie, C. Is dehydroepiandrosterone a hormone? J. Endocrinol. 2005, 187, 169–196. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, M.H.; Siddiqi, M.Z.; Ahn, S.; Kang, S.; Kim, Y.J.; Sathishkumar, N.; Yang, N.U.; Yang, D.C. Ginseng saponins and the treatment of osteoporosis: Mini literature review. J. Ginseng Res. 2013, 37, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.H.; Cho, H.J.; Kim, H.H.; Rhee, M.H.; Ryu, J.H.; Park, H.J. Inhibitory effects of total saponin from Korean red ginseng via vasodilator-stimulated phosphoprotein-Ser157 phosphorylation on thrombin-induced platelet aggregation. J. Ginseng Res. 2013, 37, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, J.; Xing, Y.; Gong, L.; Li, H.; Wu, Z.; Li, Y.; Wang, J.; Wang, Y.; Dong, L.; et al. Effects of ginsenoside Rg1 or 17beta-estradiol on a cognitively impaired, ovariectomized rat model of Alzheimer’s disease. Neuroscience 2012, 220, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Luong, T.T.; Kim, G.L.; Pyo, S.; Rhee, D.K. Korean Red Ginseng inhibits apoptosis in neuroblastoma cells via estrogen receptor β-mediated phosphatidylinositol-3 kinase/Akt signaling. J. Ginseng Res. 2014, 39, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.G.; Chan, H.Y.; Man, C.W.-Y.; Wong, M.S. Differential ERα-mediated rapid estrogenic actions of ginsenoside Rg1 and estren in human breast cancer MCF-7 cells. J. Steroid Biochem. Mol. Boil. 2014, 141, 104–112. [Google Scholar] [CrossRef]
- Kopalli, S.R.; Won, Y.J.; Hwang, S.Y.; Cha, K.M.; Kim, S.Y.; Han, C.K.; Lee, S.H.; Hong, J.Y.; Kim, S.-K. Korean red ginseng protects against doxorubicin-induced testicular damage: An experimental study in rats. J. Funct. Foods 2016, 20, 96–107. [Google Scholar] [CrossRef]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Kang, J.; Chen, D.; Han, N.; Ma, H. Ample evidence: Dehydroepiandrosterone (DHEA) conversion into activated steroid hormones occurs in adrenal and ovary in female rat. PLoS ONE 2015, 10, e0124511. [Google Scholar] [CrossRef] [PubMed]
- Gleicher, N.; Barad, D.H. Dehydroepiandrosterone (DHEA) supplementation in diminished ovarian reserve (DOR). Reprod. Boil. Endocrinol. 2011, 9, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orentreich, N.; Brind, J.L.; Rizer, R.L.; Vogelman, J.H. Age changes and sex differences in serum dehydroepiandrosterone sulfate concentrations throughout adulthood. J. Clin. Endocrinol. Metab. 1984, 59, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Vining, R.F.; McGinley, R.A.; Symons, R.G. Hormones in saliva: Mode of entry and consequent implications for clinical interpretation. Clin. Chem. 1983, 29, 1752–1756. [Google Scholar] [CrossRef]
- Association, W.M.; World Medical Association Declaration of Helsinki. Ethical principles for medical research involving human subjects. Bull. World Health Organ. 2001, 79, 373. [Google Scholar]
- Legislation. Data Protection Act. 1998. Available online: http://www.legislation.gov.uk/ukpga/1998/29/contents (accessed on 25 April 2017).
- Lee, S.M.; Bae, B.-S.; Park, H.W.; Ahn, N.G.; Cho, B.G.; Cho, Y.L.; Kwak, Y.S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Altman, D.G.; Schulz, K.F.; Moher, D.; Egger, M.; Davidoff, F.; Elbourne, D.; Gøtzsche, P.C.; Lang, T. The revised consort statement for reporting randomized trials: Explanation and elaboration. Ann. Intern. Med. 2001, 134, 663–694. [Google Scholar] [CrossRef]
- Al-Dujaili, E.A.S. Development and validation of a Simple and Direct ELISA method for the determination of conjugated (glucuronide) and non-conjugated Testosterone excretion in urine. Clin. Chim. Acta 2006, 364, 172–179. [Google Scholar] [CrossRef]
- Bancroft, J.; Sherwin, B.B.; Alexander, G.; Davidson, D.W.; Walker, A. Oral contraceptives, androgens, and the sexuality of young women: II. The role of androgens. Arch. Sex. Behav. 1991, 20, 121–135. [Google Scholar] [CrossRef]
- Coenen, C.M.; Thomas, C.M.; Borm, G.F.; Hollanders, J.M.; Rolland, R. Changes in androgens during treatment with four low-dose contraceptives. Contraception 1996, 53, 171–176. [Google Scholar] [CrossRef]
- Al-Dujaili, E.A.S.; Sharp, M. Establishing Circadian Rhythm Profiles for Salivary Testosterone in Women: Evidence of Decline during Aging. In Proceedings of the 8th European Congress of Endocrinology, Glasgow, UK, 1–5 April 2006; p. 734. [Google Scholar]
- Zumoff, B.; Strain, G.W.; Miller, L.K.; Rosner, W. Twenty-four-hour mean plasma testosterone concentration declines with age in normal premenopausal women. J. Clin. Endocrinol. Metab. 1995, 80, 1429–1430. [Google Scholar] [PubMed]
- Al-Turk, W.; Al-Dujaili, E.A.S. Effect of age, gender and exercise on salivary dehydroepiandrosterone circadian rhythm profile in human volunteers. Steroids 2016, 106, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Kiss, Z.; Ghosh, P.M. Women in cancer thematic review: Circadian rhythmicity and the influence of ‘clock’ genes on prostate cancer. Endocr. Relat. Cancer 2016, 23, T123–T134. [Google Scholar] [CrossRef] [Green Version]
- Tawab, M.A.; Bahr, U.; Karas, M.; Wurglics, M.; Schubert-Zsilavecz, M. Degradation of ginsenosides in humans after oral administration. Drug Metab. Dispos. 2003, 31, 1065–1071. [Google Scholar] [CrossRef] [Green Version]
- Granger, D.A.; Schwartz, E.B.; Booth, A.; Curran, M.; Zakaria, D. Assessing dehydroepiandrosterone in saliva: A simple radioimmunoassay for use in studies of children, adolescents and adults. Psychoneuroendocrinology 1999, 24, 567–579. [Google Scholar] [CrossRef]
- Hofman, L.F. Human saliva as a diagnostic specimen. J. Nutr. 2001, 131, 1621S–1625S. [Google Scholar] [CrossRef]
- Al-Dujaili, E.A.S.; Sharp, M.A. Female Salivary Testosterone: Measurement, Challenges and Applications, Steroids—Physiology to Clinical Medicine, 21th ed.; Ostojic, S.M., Ed.; InTechOpen: London, UK, 2012; ISBN 978-953-51-0857-3. Available online: http://www.intechopen.com/books/steroids-from-physiology-to-clinical-medicine/female-salivary-testosterone-measurement-challenges-and-applications (accessed on 23 March 2020). [CrossRef]
- Vittek, J.; L’Hommedieu, D.G.; Gordon, G.G.; Rappaport, S.C.; Southren, A.L. Direct radioimmunoassay of salivary testosterone: Correlation with free and total serum testosterone. Life Sci. 1985, 37, 711–716. [Google Scholar] [CrossRef]
- Longcope, C.; Hui, S.L.; Johnston, C.C. Free estradiol, free testostereone, and sex hormone-binding globulin in perimenopausal women. J. Clin. Endocrinol. Metab. 1987, 64, 513–518. [Google Scholar] [CrossRef]
- Voegtline, K.M.; Granger, D.A. Dispatches from the interface of salivary bioscience and neonatal research. Front. Endocrinol. 2014, 5, 25. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Demers, L.M. Bioavailable testosterone in salivary glands. Clin. Chem. 1991, 37, 139–140. [Google Scholar] [CrossRef]
- Vermeulen, A.; Verdonck, L.; Kaufman, J.M. A critical evaluation of simple methods for the estimation of free testosterone in serum. J. Clin. Endocrinol. Metab. 1999, 84, 3666–3672. [Google Scholar] [CrossRef]
- Goncharov, N.P.; Katsya, G.V.; Chagina, N.A.; Gooren, L.J. Testosterone and obesity in men under the age of 40 years. Andrologia 2009, 41, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.K.; Rosner, W.; Lee, H.; Hier, J.; Sesmilo, G.; Schoenfeld, D.; Neubauer, G.; Klibanski, A. Measurement of free testosterone in normal women and women with androgen deficiency: Comparison of methods. J. Clin. Endocrinol. Metab. 2004, 89, 525–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendel, C.M. The free hormone hypothesis: A physiologically based mathematical model. Endocr. Rev. 1989, 10, 232–274. [Google Scholar] [CrossRef] [PubMed]
- Lood, Y.; Aardal-Eriksson, E.; Webe, C.; Ahlner, J.; Ekman, B.; Wahlberg, J. Relationship between testosterone in serum, saliva and urine during treatment with intramuscular testosterone undecanoate in gender dysphoria and male hypogonadism. Andrology 2017, 6, 86–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadore, E.L.; Lhullier, M.; Brentano, E.; Silva, M.; Ambrosini, M.; Spinelli, R.; Silva, R.; Kruel, L. Correlations between serum and salivary hormonal concentrations in response to resistance exercise. Sports Sci. 2008, 26, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.R.; Hackney, A.C. Relationship between salivary and serum testosterone levels in response to different exercise intensities. Hormones 2014, 14, 258–264. [Google Scholar]
- Black, A.E.; Welch, A.; Bingham, S.A. Validation of dietary intakes measured by diet history against 24 h urinary nitrogen excretion and energy expenditure measured by the doubly-labelled water method in middle-aged women. Br. J. Nutr. 2000, 83, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Al-Dujaili, E.A.S.; Bryant, M.L. Effect of Meal Fat Content on Salivary Testosterone and Cortisol Levels in Healthy Female Volunteers. In Proceedings of the 196th Meeting of the Society for Endocrinology and Society for Endocrinology joint Endocrinology and Diabetes Day, London, UK, 7–9 November 2005; Volume 10, p. 75. [Google Scholar]
- Berrino, F.; Bellati, C.; Secreto, G.; Camerini, E.; Pala, V.; Panico, S.; Allegro, G.; Kaaks, R. Reducing bioavailable sex hormones through a comprehensive change in diet: The diet and androgens (DIANA) randomized trial. Cancer Epidemiol. Biomark. Prev. 2001, 10, 25–33. [Google Scholar]
- Gallagher, P.; Leitch, M.; Massey, A.E.; McAllister-Williams, R.H.; Young, A.H. Assessing cortisol and dehydroepiandrosterone (DHEA) in saliva: Effects of collection method. J. Psychopharmacol. 2006, 20, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Ahn, R.S.; Lee, Y.J.; Choi, J.Y.; Kwon, H.B.; Chun, S.I. Salivary Cortisol and DHEA Levels in the Korean Population: Age-Related Differences, Diurnal Rhythm, and Correlations with Serum Levels. Yonsei Med. J. 2007, 48, 379–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aizawa, K.; Iemitsu, M.; Maeda, S.; Otsuki, T.; Sato, K.; Ushida, T.; Mesaki, N.; Akimoto, T. Acute exercise activates local bioactive androgen metabolism in skeletal muscle. Steroids 2010, 75, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Heaney, J.L.; Carroll, U.; Phillips, A.C. Physical activity, life events stress, cortisol, and DHEA: Preliminary findings that physical activity may buffer against the negative effects of stress. J. Aging Phys. Act. 2014, 22, 465–473. [Google Scholar] [CrossRef]
- Flynn, M.A.; Weaver-Osterholtz, D.; Sharpe-Timms, K.; Allen, S.; Krause, G. Dehydroepiandrosterone replacement in aging humans. J. Clin. Endocrinol. Metab. 1999, 84, 1527–1533. [Google Scholar] [CrossRef]
- Jenkinson, C.; Petroczi, A.; Naughton, D.P. Effects of dietary components on testosterone metabolism via UDP-glucuronosyltransferase. Front. Endocrinol. 2013, 4, 80. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Average Daily Value | Pre Ginseng | ± SD | Post Ginseng | ± SD | p-value |
|---|---|---|---|---|---|
| Group A Testo (pg/mL) | 76.3 | 16.6 | 98.4 | 21.1 | < 0.01 |
| Group B Testo (pg/mL) | 61.6 | 16.9 | 68.4 | 11.5 | 0.132 |
| Group A DHEA (ng/mL) | 1.53 | 0.63 | 1.98 | 0.89 | 0.02 |
| Group B DHEA (ng/mL) | 0.91 | 0.32 | 1.62 | 0.49 | 0.014 |
| Nutrient | Mean | ± SD | Paired t-test | |
|---|---|---|---|---|
| Group A | ||||
| Energy (kcal) | Pre | 1610.46 | 336.701 | 0.376 |
| Post | 1446.11 | 384.541 | ||
| Protein (g) | Pre | 64.44 | 16.704 | 0.435 |
| Post | 58.22 | 23.419 | ||
| Carbohydrate (g) | Pre | 216.11 | 74.824 | 0.323 |
| Post | 187.00 | 36.166 | ||
| Fat (g) | Pre | 59.44 | 18.290 | 0.500 |
| Post | 54.44 | 24.501 | ||
| Group B | ||||
| Energy (kcal) | Pre | 1696.25 | 258.578 | 0.050 |
| Post | 2030.38 | 382.317 | ||
| Protein (g) | Pre | 72.25 | 16.968 | 0.147 |
| Post | 83.38 | 17.541 | ||
| Carbohydrate (g) | Pre | 229.00 | 42.007 | 0.281 |
| Post | 247.88 | 38.698 | ||
| Fat (g) | Pre | 56.58 | 17.883 | 0.041 |
| Post | 81.25 | 33.856 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Dujaili, E.A.S.; Hajleh, M.N.A.; Chalmers, R. Effects of Ginseng Ingestion on Salivary Testosterone and DHEA Levels in Healthy Females: An Exploratory Study. Nutrients 2020, 12, 1582. https://doi.org/10.3390/nu12061582
Al-Dujaili EAS, Hajleh MNA, Chalmers R. Effects of Ginseng Ingestion on Salivary Testosterone and DHEA Levels in Healthy Females: An Exploratory Study. Nutrients. 2020; 12(6):1582. https://doi.org/10.3390/nu12061582
Chicago/Turabian StyleAl-Dujaili, Emad A. S., Maha N. Abu Hajleh, and Ruth Chalmers. 2020. "Effects of Ginseng Ingestion on Salivary Testosterone and DHEA Levels in Healthy Females: An Exploratory Study" Nutrients 12, no. 6: 1582. https://doi.org/10.3390/nu12061582
APA StyleAl-Dujaili, E. A. S., Hajleh, M. N. A., & Chalmers, R. (2020). Effects of Ginseng Ingestion on Salivary Testosterone and DHEA Levels in Healthy Females: An Exploratory Study. Nutrients, 12(6), 1582. https://doi.org/10.3390/nu12061582