Effect of a Nutritional Intervention on the Intestinal Microbiota of Vertically HIV-Infected Children: The Pediabiota Study

 , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Methods
2.1. Study Design
2.2. Nutritional Intervention
2.3. Randomization
2.4. Nucleic Acid Purification, Amplification of the 16S rRNA Gene, Sequencing and Bioinformatics Analysis
2.4.1. Nucleic Acid Purification
2.4.2. Amplification of the 16S rRNA Gene
2.4.3. 16S RNA Gene Analysis. Biodiversity and Clustering
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
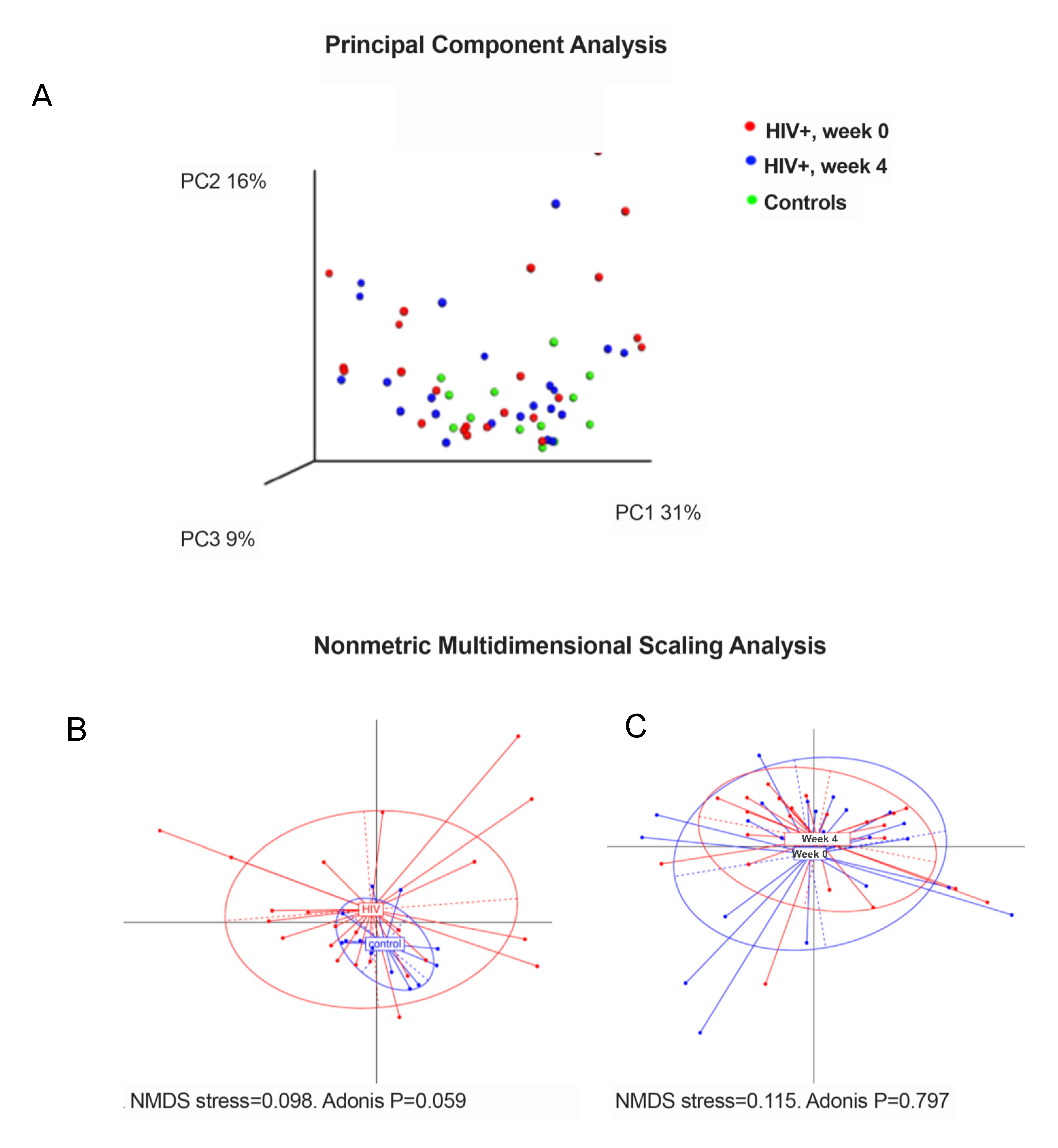
3.2. Analysis of Fecal Microbiota Structure
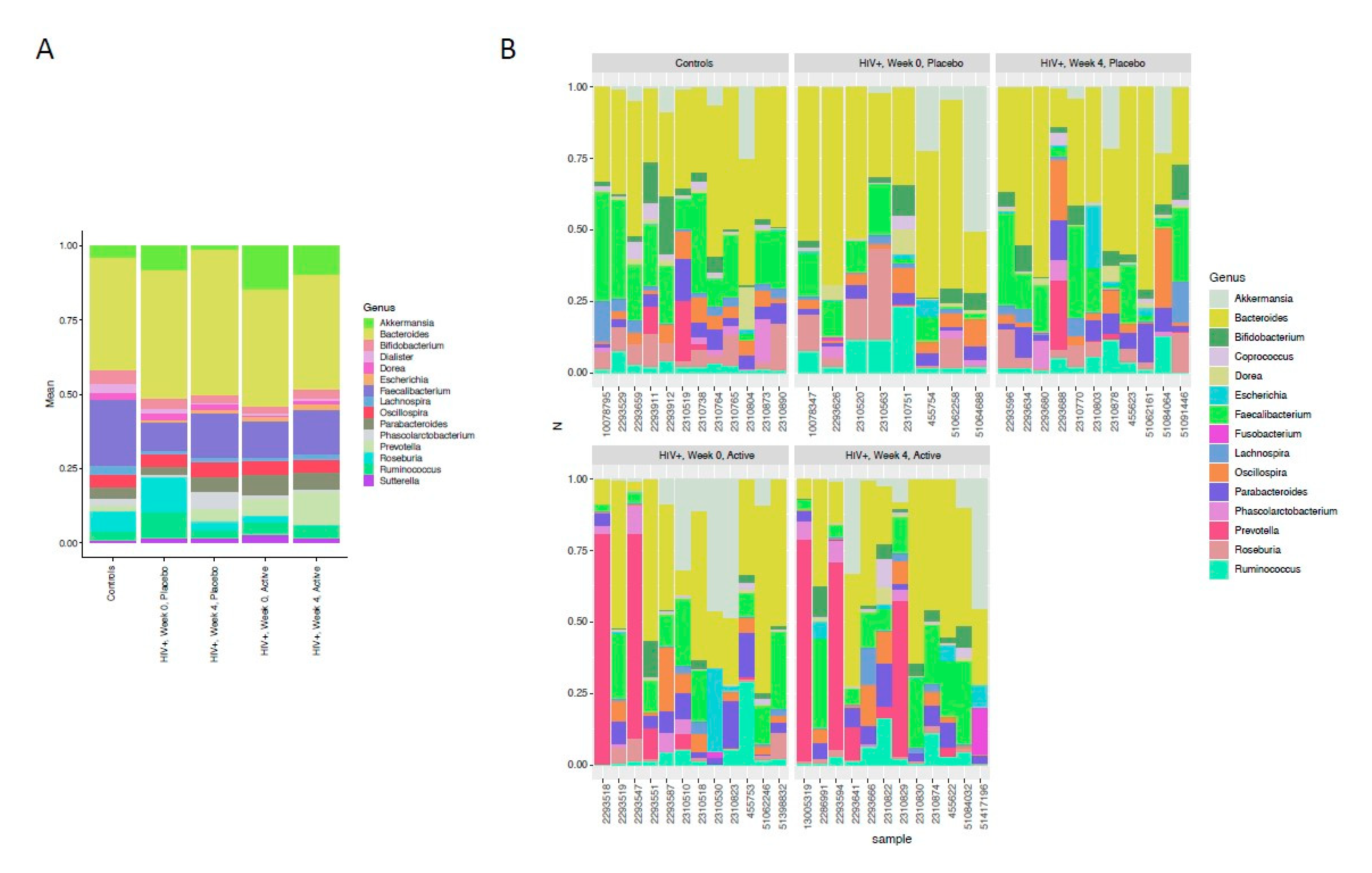
3.3. Bacterial Biomarkers of HIV Infection and Taxa Significantly Affected by the Intervention
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Serrano-Villar, S.; Rojo, D.; Martínez-Martínez, M.; Deusch, S.; Vázquez-Castellanos, J.F.; Sainz, T.; Vera, M.; Moreno, S.; Estrada, V.; Gosalbes, M.J.; et al. HIV infection results in metabolic alterations in the gut microbiota different from those induced by other diseases. Sci. Rep. 2016, 6, 26192. [Google Scholar] [CrossRef]
- Vujkovic-Cvijin, I.; Somsouk, M. HIV and the gut microbiota: Composition, consequences, and avenues for amelioration. Curr. HIV/AIDS Rep. 2019, 16, 201–213. [Google Scholar] [CrossRef]
- Mutlu, E.A.; Keshavarzian, A.; Losurdo, J.; Swanson, G.; Siewe, B.; Forsyth, C.; French, A.; DeMarais, P.; Sun, Y.; Koenig, L.; et al. A compositional look at the human gastrointestinal microbiome and immune activation parameters in HIV infected subjects. PLoS Pathog. 2014, 10, e1003829. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Villar, S.; Rojo, D.; Martínez-Martínez, M.; Deusch, S.; Vázquez-Castellanos, J.F.; Bargiela, R.; Sainz, T.; Vera, M.; Moreno, S.; Estrada, V.; et al. Gut bacteria metabolism impacts immune recovery in HIV-infected individuals. EBioMedicine 2016, 8, 203–216. [Google Scholar] [CrossRef]
- Guillén, Y.; Noguera-Julian, M.; Rivera, J.; Casadellà, M.; Zevin, A.S.; Rocafort, M.; Parera, M.; Rodríguez, C.; Arumí, M.; Carrillo, J.; et al. Low nadir CD4+ T-cell counts predict gut dysbiosis in HIV-1 infection. Mucosal Immunol. 2018, 12, 119–129, in press. [Google Scholar] [CrossRef] [PubMed]
- Mudd, J.C.; Brenchley, J.M. Gut mucosal barrier dysfunction, microbial dysbiosis, and their role in HIV-1 disease progression. J. Infect. Dis. 2016, 214 (Suppl. 2), S58–S66. [Google Scholar] [CrossRef] [PubMed]
- Somsouk, M.; Estes, J.D.; Deleage, C.; Dunham, R.M.; Albright, R.; Inadomi, J.M.; Martin, J.N.; Deeks, S.G.; McCune, J.M.; Peter, W. Hunt gut epithelial barrier and systemic inflammation during chronic HIV infection. AIDS 2015, 29, 43–51. [Google Scholar] [CrossRef]
- Sessa, L.; Reddel, S.; Manno, E.; Quagliariello, A.; Cotugno, N.; del Chierico, F.; Amodio, D.; Capponi, C.; Leone, F.; Bernardi, S.; et al. Distinct gut microbiota profile in ART-treated perinatally HIV-infected patients associated with cardiac and inflammatory biomarkers. AIDS 2019, 1, 1001–1011. [Google Scholar] [CrossRef]
- Kaur, U.S.; Shet, A.; Rajnala, N.; Gopalan, B.P.; Moar, P.; Himanshu, D.; Singh, B.P.; Chaturvedi, R.; Tandon, R. High abundance of genus prevotella in the gut of perinatally HIV-infected children is associated with IP-10 levels despite therapy. Sci. Rep. 2018, 8, 17679. [Google Scholar] [CrossRef]
- Flygel, T.T.; Sovershaeva, E.; Classen-Weitz, S.; Hjerde, E.; Mwaikono, K.S.; Odland, J.Ø.; Ferrand, R.A.; Mchugh, G.; Gutteberg, T.J.; Nicol, M.P.; et al. Composition of gut microbiota of children and adolescents with perinatal HIV infection taking antiretroviral therapy in Zimbabwe. J. Infect. Dis. 2020, 221, 483–492. [Google Scholar] [CrossRef]
- Ruiz-Mateos, E.; Rubio, A.; Vallejo, A.; de la Rosa, R.; Sanchez-Quijano, A.; Lissen, E.; Leal, M. Thymic volume is associated independently with the magnitude of short- and long-term repopulation of CD4+ T cells in HIV-infected adults after highly active antiretroviral therapy (HAART). Clin. Exp. Immunol. 2004, 136, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Dunham, R.M.; Iwai, S.; Maher, M.C.; Albright, R.G.; Broadhurst, M.J.; Hernandez, R.D.; Lederman, M.M.; Huang, Y.; Somsouk, M.; et al. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Sci. Transl. Med. 2013, 5, 193ra91. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.M.; Lee, E.J.; Kotter, C.V.; Austin, G.L.; Dong, Z.; Hecht, D.K.; Gianella, S.; Siewe, B.; Smith, D.M.; Landay, A.L.; et al. An altered intestinal mucosal microbiome in HIV-1 infection is associated with mucosal and systemic immune activation and endotoxemia. Mucosal Immunol. 2014, 7, 983–994. [Google Scholar] [CrossRef]
- Williams, W.B.; Liao, H.X.; Moody, M.A.; TKepler, h.B.; Alam, S.M.; Gao, F.; Wiehe, K.; Trama, A.M.; Jones, K.; Zhang, R.; et al. Diversion of HIV-1 vaccine-induced immunity by gp41-microbiota cross-reactive antibodies. Science 2015, 349, 6249. [Google Scholar] [CrossRef] [PubMed]
- Gori, A.; Rizzardini, G.; Van’t Land, B.; Amor, K.B.; van Schaik, J.; Torti, C.; Quirino, T.; Tincati, C.; Bandera, A.; Knol, J.; et al. Specific prebiotics modulate gut microbiota and immune activation in HAART-naive HIV-infected adults: Results of the “COPA” pilot randomized trial. Mucosal Immunol. 2011, 4, 1–10. [Google Scholar] [CrossRef]
- Vázquez-Castellanos, J.F.; Serrano-Villar, S.; Latorre, A.; Artacho, A.; Ferrús, M.L.; Madrid, N.; Vallejo, A.; Sainz, T.; Martínez-Botas, J.; Ferrando-Martínez, S.; et al. Altered metabolism of gut microbiota contributes to chronic immune activation in HIV-infected individuals. Mucosal Immunol. 2015, 8, 760–772. [Google Scholar] [CrossRef]
- Villar-García, J.; Hernández, J.J.; Güerri-Fernández, R.; González, A.; Lerma, E.; Guelar, A.; Saenz, D.; Sorlí, L.; Montero, M.; Horcajada, J.P.; et al. Effect of probiotics (saccharomyces boulardii) on microbial translocation and inflammation in HIV-treated patients. JAIDS J. Acquir. Immune Defic. Syndr. 2015, 68, 256–263. [Google Scholar] [CrossRef]
- Asmuth, D.M.; Ma, Z.-M.; Albanese, A.; Sandler, N.G.; Devaraj, S.; Knight, T.H.; Flynn, N.M.; Yotter, T.; Garcia, Ju.; Tsuchida, E.; et al. Oral serum-derived bovine immunoglobulin improves duodenal immune reconstitution and absorption function in patients with HIV enteropathy. AIDS 2013, 27, 2207–2217. [Google Scholar] [CrossRef]
- Cahn, P.; Ruxrungtham, K.; Gazzard, B.; Diaz, R.S.; Gori, A.; Kotler, D.P.; Vriesema, A.; Georgiou, N.A.; Garssen, J.; Clerici, M.; et al. The immunomodulatory nutritional intervention NR100157 reduced CD4+ T-cell decline and immune activation: A 1-year multicenter randomized controlled double-blind trial in HIV-infected persons not receiving antiretroviral therapy (The BITE Study). Clin. Infect. Dis. 2013, 57, 139–146. [Google Scholar] [CrossRef]
- Generoso, S.V.; Viana, M.L.; Santos, R.G.; Arantes, R.M.E.; Martins, F.S.; Nicoli, J.R.; Machado, J.A.N.; Correia, M.I.T.D.; Cardoso, V.N. Protection against increased intestinal permeability and bacterial translocation induced by intestinal obstruction in mice treated with viable and heat-killed Saccharomyces boulardii. Eur. J. Nutr. 2011, 50, 261–269. [Google Scholar] [CrossRef]
- De-Souza, D.A.; Greene, L.J. Intestinal permeability and systemic infections in critically ill patients: Effect of glutamine. Crit. Care. Med. 2005, 33, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Viana, M.L.; Santos, R.G.C.; Generoso, S.V.; Arantes, R.M.E.; Correia, M.I.T.D.; Cardoso, V.N. Pretreatment with arginine preserves intestinal barrier integrity and reduces bacterial translocation in mice. Nutrition 2010, 26, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Barham, J.B.; Edens, M.B.; Fonteh, A.N.; Johnson, M.M.; Easter, L.; Chilton, F.H. Addition of eicosapentaenoic acid to gamma-linolenic acid-supplemented diets prevents serum arachidonic acid accumulation in humans. J. Nutr. 2000, 130, 1925–1931. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.; Livak, B.; Burke-Miller, J.; French, A.L.; Glesby, M.J.; Sharma, A.; Young, M.; Villacres, M.C.; Tien, P.C.; Golub, E.T.; et al. Vitamin D insufficiency may impair CD4 recovery among women’s interagency HIV study participants with advanced disease on HAART. AIDS 2013, 27, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Majano, P.; Alonso-Lebrero, J.L.; Janczyk, A.; Martín-Vichez, S.; Molina-Jiménez, F.; Brieva, A.; Pivel, J.P.; González, S.; López-Cabrera, M. Ricardo Moreno-Otero AM3 inhibits LPS-induced iNOS expression in mice. Int. Immunopharmacol. 2005, 5, 1165–1170. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The ribosomal database project: Improved alignments and new tools for rRNA analysis. Nucleic. Acids. Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Sarrabayrouse, G.; Bossard, C.; Chauvin, J.M.; Jarry, A.; Meurette, G.; Quévrain, E.; Bridonneau, C.; Preisser, L.; Asehnoune, K.; Labarrière, N.; et al. CD4CD8αα lymphocytes, a novel human regulatory T cell subset induced by colonic bacteria and deficient in patients with inflammatory bowel disease. PLoS Biol. 2014, 12, e1001833. [Google Scholar] [CrossRef]
- Laval, L.; Martin, R.; Natividad, J.N.; Chain, F.; Miquel, S.; de Maredsous, C.D.; Capronnier, S.; Sokol, H.; Verdu, E.F.; van Vlieg, J.E.T.; et al. Lactobacillus rhamnosus CNCM I-3690 and the commensal bacterium Faecalibacterium prausnitzii A2-165 exhibit similar protective effects to induced barrier hyper-permeability in mice. Gut Microbes 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Machiels, K.; Joossens, M.; Sabino, J.; de Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. GUT 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Seekatz, A.M.; Aas, J.; Gessert, C.E.; Rubin, T.A.; Saman, D.M.; Bakken, J.S.; Young, V.B. Recovery of the gut microbiome following fecal microbiota transplantation. MBIO 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Neff, C.P.; Krueger, O.; Xiong, K.; Arif, S.; Nusbacher, N.; Schneider, J.M.; Cunningham, A.W.; Armstrong, A.; Li, S.; McCarter, M.D.; et al. Fecal microbiota composition drives immune activation in HIV-infected individuals. EBioMedicine 2018, 30, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Noguera-Julian, M.; Rocafort, M.; Guillén, Y.; Rivera, J.; Casadellà, M.; Nowak, P.; Hildebrand, F.; Zeller, G.; Parera, M.; Bellido, R.; et al. Gut microbiota linked to sexual preference and HIV infection. EBioMedicine 2016, 5, 135–146. [Google Scholar] [CrossRef]
- Moya, A.; Ferrer, M. Functional redundancy-induced stability of gut microbiota subjected to disturbance. Trends Microbiol. 2016, 24, 402–413. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- Serrano-Villar, S.; Vázquez-Castellanos, J.F.; Vallejo, A.; Latorre, A.; Sainz, T.; Ferrando-Martínez, S.; Rojo, D.; Martínez-Botas, J.; del Romero, J.; Madrid, N.; et al. The effects of prebiotics on microbial dysbiosis, butyrate production and immunity in HIV-infected subjects. Mucosal Immunol. 2017, 10, 1279–1293. [Google Scholar] [CrossRef]
- Serrano-Villar, S.; de Lagarde, M.; Vázquez-Castellanos, J.; Vallejo, A.; Bernadino, J.I.; Madrid, N.; Matarranz, M.; Díaz-Santiago, A.; Gutiérrez, C.; Cabello, A.; et al. Effects of immunonutrition in advanced human immunodeficiency virus disease: A randomized placebo-controlled clinical trial (promaltia study). Clin. Infect. Dis. 2019, 68, 120–130. [Google Scholar] [CrossRef]
- Stiksrud, B.; Nowak, P.; Nwosu, F.C.; Kvale, D.; Thalme, A.; Sonnerborg, A.; Ueland, P.M.; Holm, K.; Birkeland, S.E.; Dahm, A.E.A.; et al. Reduced levels of D-dimer and changes in gut microbiota composition after probiotic intervention in HIV-Infected individuals on stable ART. J. Acquir. Immune. Defic. Syndr. 2015, 70, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Kostic, A.D.; Howitt, M.R.; Garrett, W.S. Exploring host-microbiota interactions in animal models and humans. Genes Dev. 2013, 27, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Biedermann, L.; Zeitz, J.; Mwinyi, J.; Sutter-Minder, E.; Rehman, A.; Ott, S.J.; Steurer-Stey, C.; Frei, A.; Frei, P.; Scharl, M.; et al. Smoking cessation induces profound changes in the composition of the intestinal microbiota in humans. PLoS ONE 2013, 8, e59260. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Villar, S.; Vásquez-Domínguez, E.; Pérez-Molina, J.A.; Sainz, T.; de Benito, A.; Latorre, A.; Moya, A.; Gosalbes, M.J.; Moreno, S. HIV, HPV, and microbiota: Partners in crime? AIDS 2017, 31, 591–594. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active | Placebo | |
|---|---|---|
| Energy | 65.3 kcal | 74.9 kcal |
| Total lipids | 5.1 g | 0 |
| EPA | 3.2 g | 0 |
| DHA | 0.3 g | 0 |
| GLA | 1.7 g | 0 |
| Total carbohydrates | 10.3 g | 10.9 g |
| Long-chain FOS | 4 g | 0 |
| Short-chain GOS | 3.3 g | 0 |
| Maltodextrin (excipient) | 3 | 0 |
| Total proteins | 3.3 g | 6.9 g |
| L-Glutamine | 2 g | 0 |
| L-Arginine | 1.2 g | 0 |
| Others | 1.2 g | 0 |
| Saccharomyces boulardii | 0.17 g | 0 |
| AM3 | 1 g | 0 |
| Vitamin D | 5.3 × 103 MU | 0 |
| TOTAL | 21.1 | 21.1 |
| Controls N = 11 | Placebo N = 12 | Nutritional Intervention N = 12 | p | |
|---|---|---|---|---|
| Female (n,%) | 4 (36.4) | 7 (58.3) | 8 (66.7) | 1.000 |
| Age (years), mean (SD) | 10 (4.4) | 13.8 (3.6) | 10 (3.4) | 0.064 |
| Caucasian (n,%) | 7 (64) | 6 (50) | 7 (58.3) | 1.000 |
| CD4 count (cells/mm3) | - | 556 (453–754) | 852 (617–1182) | 0.0496 |
| CD4/CD8 ratio | - | 1.1 (0.56–1.67) | 1.4 (1.09–1.94) | 0.106 |
| CD4 Nadir (cells/mm3) | - | 333 (169–382) | 519 (384–979) | 0.006 |
| PI based ART (n,%) | - | 8 (66.7) | 9 (75) | 0.136 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sainz, T.; Gosalbes, M.J.; Talavera, A.; Jimenez-Hernandez, N.; Prieto, L.; Escosa, L.; Guillén, S.; Ramos, J.T.; Muñoz-Fernández, M.Á.; Moya, A.; et al. Effect of a Nutritional Intervention on the Intestinal Microbiota of Vertically HIV-Infected Children: The Pediabiota Study. Nutrients 2020, 12, 2112. https://doi.org/10.3390/nu12072112
Sainz T, Gosalbes MJ, Talavera A, Jimenez-Hernandez N, Prieto L, Escosa L, Guillén S, Ramos JT, Muñoz-Fernández MÁ, Moya A, et al. Effect of a Nutritional Intervention on the Intestinal Microbiota of Vertically HIV-Infected Children: The Pediabiota Study. Nutrients. 2020; 12(7):2112. https://doi.org/10.3390/nu12072112
Chicago/Turabian StyleSainz, Talía, María José Gosalbes, Alba Talavera, Nuria Jimenez-Hernandez, Luis Prieto, Luis Escosa, Sara Guillén, José Tomás Ramos, María Ángeles Muñoz-Fernández, Andrés Moya, and et al. 2020. "Effect of a Nutritional Intervention on the Intestinal Microbiota of Vertically HIV-Infected Children: The Pediabiota Study" Nutrients 12, no. 7: 2112. https://doi.org/10.3390/nu12072112
APA StyleSainz, T., Gosalbes, M. J., Talavera, A., Jimenez-Hernandez, N., Prieto, L., Escosa, L., Guillén, S., Ramos, J. T., Muñoz-Fernández, M. Á., Moya, A., Navarro, M. L., Mellado, M. J., & Serrano-Villar, S. (2020). Effect of a Nutritional Intervention on the Intestinal Microbiota of Vertically HIV-Infected Children: The Pediabiota Study. Nutrients, 12(7), 2112. https://doi.org/10.3390/nu12072112