Ginseng-Induced Changes to Blood Vessel Dilation and the Metabolome of Rats

 , ,
, ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Ginseng Extract
2.2. Animals and Ginseng Administration
2.3. Blood Lipid Characteristics
2.4. Global Metabolomics Analysis
2.5. Steroid Hormone Analysis
2.6. Analysis of Plasma Cytokines and Enzymes
2.7. Blood Vessel Tension Measurement Using an Organ Chamber
2.8. Statistical Analysis
3. Results
3.1. Animal Characteristics
3.2. Metabolomic Analysis
3.3. Blood Vessel Health-Related Factors
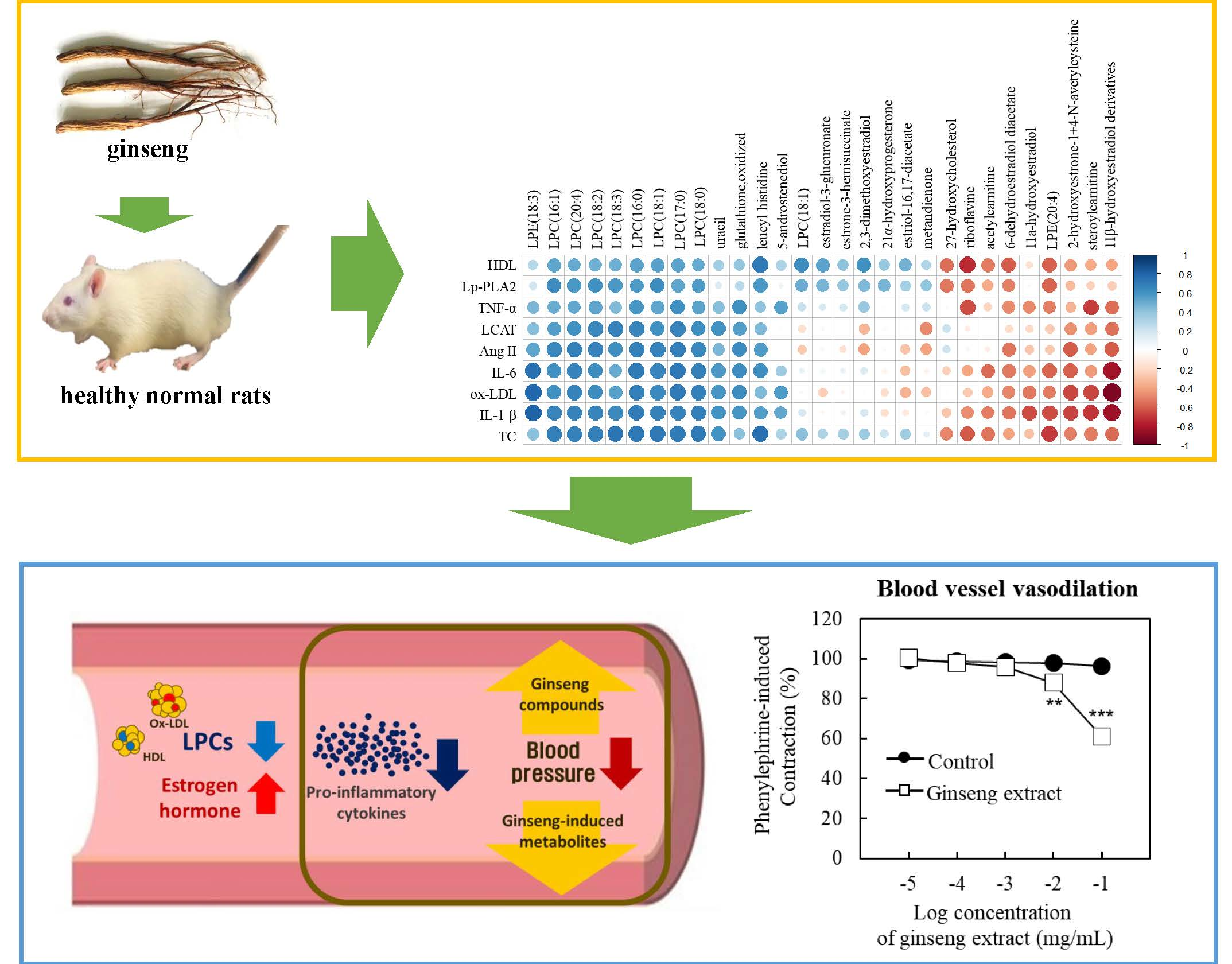
3.4. Correlation Analysis
3.5. Effect of Ginseng Extract on Blood Vessel Tension
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, J.-S. Investigation of phenolic, flavonoid, and vitamin contents in different parts of Korean ginseng (Panax ginseng C.A. Meyer). Prev. Nutr. Food Sci. 2016, 21, 263–270. [Google Scholar] [CrossRef]
- Patel, S.; Rauf, A. Adaptogenic herb ginseng (Panax) as medical food: Status quo and future prospects. Biomed. Pharmacother. 2017, 85, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Jovanovski, E.; Bateman, E.A.; Bhardwaj, J.; Fairgrieve, C.; Mucalo, I.; Jenkins, A.L.; Vuksan, V. Effect of Rg3-enriched Korean red ginseng (Panax ginseng) on arterial stiffness and blood pressure in healthy individuals: A randomized controlled trial. J. Am. Soc. Hypertens. 2014, 8, 537–541. [Google Scholar] [CrossRef]
- Nagar, H.; Choi, S.; Jung, S.; Jeon, B.H.; Kim, C.-S. Rg3-enriched Korean red ginseng enhances blood pressure stability in spontaneously hypertensive rats. Integr. Med. Res. 2016, 5, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Cha, T.W.; Kim, M.; Kim, M.; Chae, J.S.; Lee, J.H. Blood pressure-lowering effect of Korean red ginseng associated with decreased circulating Lp-PLA 2 activity and lysophosphatidylcholines and increased dihydrobiopterin level in prehypertensive subjects. Hypertens. Res. 2016, 39, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Lu, Y.; Shen, T.; Huang, X.; Man, Y.; Wang, S.; Li, J. Panax notogingseng saponins suppress RAGE/MAPK signaling and NF-kappaB activation in mice. Cell. Physiol. Biochem. 2012, 100730, 875–882. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Zhang, H.G.; Zhang, G.Y.; Fan, J.S.; Li, X.H.; Liu, Y.H.; Li, S.H.; Lian, X.M.; Tang, Z. Panax notoginseng saponins attenuate atherosclerosis in rats by regulating the blood lipid profile and an anti-inflammatory action. Clin. Exp. Pharmacol. Physiol. 2008, 35, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- McGarrah, R.W.; Crown, S.B.; Zhang, G.F.; Shah, S.H.; Newgard, C.B. Cardiovascular metabolomics. Circ. Res. 2018, 122, 1238–1258. [Google Scholar] [CrossRef]
- Ho, J.E.; Lyass, A.; Courchesne, P.; Chen, G.; Liu, C.; Yin, X.; Hwang, S.J.; Massaro, J.M.; Larson, M.G.; Levy, D. Protein biomarkers of cardiovascular disease and mortality in the community. J. Am. Heart Assoc. 2018, 7, 1–30. [Google Scholar] [CrossRef]
- Costa, A.; Franco, O.L. Impact and influence of “omics” technology on hyper tension studies. Int. J. Cardiol. 2017, 228, 1022–1034. [Google Scholar] [CrossRef]
- Kim, H.J.; Cho, C.W.; Hwang, J.T.; Son, N.; Choi, J.H.; Shim, G.S.; Han, C.K. LC-MS-based metabolomic analysis of serum and livers from red ginseng-fed rats. J. Ginseng Res. 2013, 37, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Nofer, J.-R. Estrogens and atherosclerosis: Insights from animal models and cell systems. J. Mol. Endocrinol. 2012, 48, R13–R29. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, N.W. Ultra performance liquid chromatography-tandem mass spectrometry method for profiling of steroid metabolome in human tissue. Anal. Chem. 2013, 85, 4951–4960. [Google Scholar] [CrossRef] [PubMed]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, J.; Kim, S.; Sohn, J. The effect of sugammadex on the vascular tone of isolated rat aorta. Korean J. Anesth. 2018, 71, 242–243. [Google Scholar] [CrossRef]
- Motamed, M.; Nargesi, A.A.; Heidari, B.; Mirmiranpour, H.; Esteghamati, A.; Nakhjavani, M. Oxidized low-density lipoprotein (ox-LDL) to LDL ratio (ox-LDL/LDL) and ox-LDL to High-Density Lipoprotein Ratio (ox-LDL/HDL). Clin. Lab. 2016, 62, 1609–1617. [Google Scholar] [CrossRef]
- Mensink, R.P.; Zock, P.L.; Kester, A.D.M.; Katan, M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: A meta-analysis of 60 controlled trials. Am. J. Clin. Nutr. 2003, 77, 1146–1155. [Google Scholar] [CrossRef]
- Silva-Neto, M.A.C.; Lopes, A.H.; Atella, G.C. Here, there, and everywhere: The ubiquitous distribution of the immunosignaling molecule lysophosphatidylcholine and its role on chagas disease. Front. Immunol. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Schmitz, G.; Ruebsaamen, K. Metabolism and atherogenic disease association of lysophosphatidylcholine. Atherosclerosis 2010, 208, 10–18. [Google Scholar] [CrossRef]
- Choi, J.; Zhang, W.; Gu, X.; Chen, X.; Hong, L.; Laird, J.M.; Salomon, R.G. Lysophosphatidylcholine is generated by spontaneous deacylation of oxidized phospholipids. Chem. Res. Toxicol. 2011, 24, 111–118. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kobayashi, T.; Kamata, K. Role of lysophosphatidylcholine (LPC) in atherosclerosis. Curr. Med. Chem. 2007, 14, 3209–3220. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Kim, J.H. A review on the medicinal potentials of ginseng and ginsenosides on cardiovascular diseases. J. Ginseng Res. 2014, 38, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Buettner, C.; Yeh, G.Y.; Phillips, R.S.; Mittleman, M.A.; Kaptchuk, T.J. Systematic review of the effects of ginseng on cardiovascular risk factors. Ann. Pharmacother. 2006, 40, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Kwon, S.K.; Nagar, H.; Jung, S.B.; Jeon, B.H.; Kim, C.S.; Oh, J.H.; Song, H.J.; Kim, C.S. Rg3-enriched Korean red ginseng improves vascular function in spontaneously hypertensive rats. J. Ginseng Res. 2014, 38, 244–250. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Park, J.; Song, H.; Kim, S.K.; Lee, M.S.; Rhee, D.K.; Lee, Y.J. Effects of ginseng on two main sex steroid hormone receptors: Estrogen and androgen receptors. J. Ginseng Res. 2017, 41, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Raghvendra, K.; Jackson, E.K. Cardiovascular protective effects of 17β-estradiol metabolites. J. Appl. Physiol. 2001, 91, 1868–1883. [Google Scholar]
- Cho, K.S.; Park, C.W.; Kim, C.; Jeon, H.Y.; Kim, W.G.; Lee, S.J.; Kim, Y.; Lee, J.Y.; Choi, Y.D. Effects of Korean ginseng berry extract (GB0710) on penile erection: Evidence from in vitro and in vivo studies. Asian J. Androl. 2013, 15, 503–507. [Google Scholar] [CrossRef]
- Hong, S.Y.; Kim, J.Y.; Ahn, H.Y.; Shin, J.; Kwon, O. Panax ginseng extract rich in ginsenoside protopanaxatriol attenuates blood pressure elevation in spontaneously hypertensive rats by affecting the Akt-dependent phosphorylation of endothelial nitric oxide synthase. J. Agric. Food Chem. 2012, 60, 3086–3091. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, J.; Liu, P.; Lau, C.W.; Gao, Z.; Zhou, D.; Tang, J.; Ng, C.F.; Huang, Y. Ginsenoside Rb3 attenuates oxidative stress and preserves endothelial function in renal arteries from hypertensive rats. Br. J. Pharmacol. 2014, 171, 3171–3181. [Google Scholar] [CrossRef]
- Lee, K.H.; Bae, I.Y.; Park, S.I.; Park, J.; Lee, H.G. Antihypertensive effect of Korean Red Ginseng by enrichment of ginsenoside Rg3 and arginine e fructose. J. Ginseng Res. 2016, 40, 237–244. [Google Scholar] [CrossRef]
- Drzazga, A.; Sowińska, A.; Koziołkiewicz, M. Lysophosphatidylcholine and lysophosphatidylinosiol novel promissing signaling molecules and their possible therapeutic activity. Acta Pol. Pharm. Drug Res. 2014, 71, 887–899. [Google Scholar]
- Chow, R.W.Y.; Handelsman, D.J.; Ng, M.K.C. Minireview: Rapid actions of sex steroids in the endothelium. Endocrinology 2010, 151, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Guo, R.; Wu, S.L.; Sun, M.; Li, M. Protective effects of ginsenoside Rb1 on human umbilical vein endothelial cells in vitro. J. Cardiovasc. Pharmacol. 2007, 50, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-M.; Namkoong, S.; Yun, Y.-G.; Hong, H.-D.; Lee, Y.-C.; Ha, K.-S.; Lee, H.; Kwon, H.J.; Kwon, Y.-G.; Kim, Y.-M. Water extract of Korean red ginseng stimulates angiogenesis by activating the PI3K/Akt-dependent ERK1/2 and eNOS pathways in human umbilical vein endothelial cells. Biol. Pharm. Bull. 2007, 30, 1674–1679. [Google Scholar] [CrossRef]
- Ceballos-picot, I.; Witko-sarsat, V.; Merad-boudia, M.; Nguyen, A.T.; Venin, M.T.H.; Jaudon, M.C.; Zingraff, J.; Verger, C.; Jungers, P.; Descamps-latscha, B. Glutathione antioxidant system as a marker of oxidative stress in chronic renal failure. Free Radic. Biol. Med. 1996, 21, 845–853. [Google Scholar] [CrossRef]
- Ramesh, T.; Kim, S.; Hwang, S.; Sohn, S.; Yoo, S.; Kim, S. Panax ginseng reduces oxidative stress and restores antioxidant capacity in aged rats. Nutr. Res. 2012, 32, 718–726. [Google Scholar] [CrossRef]
- López, M.V.N.; Cuadrado, M.P.G.; Ruiz-poveda, O.M.P.; Villar, A.M.; Fresno, D.; Accame, M.E.C. Neuroprotective effect of individual ginsenosides on astrocytes primary culture. (BBA)-Gen. Subj. 2007, 1770, 1308–1316. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.; Cheng, X.; Wei, F.; Bai, X.; Lin, R. Metabolomics analysis reveals the association between lipid abnormalities and oxidative stress, inflammation, fibrosis, and Nrf2 dysfunction in aristolochic acid-induced nephropathy. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef]
- Kalim, S.; Clish, C.B.; Wenger, J.; Elmariah, S.; Yeh, R.W.; Deferio, J.J.; Pierce, K.; Deik, A.; Gerszten, R.E.; Thadhani, R.; et al. A plasma long-chain acylcarnitine predicts cardiovascular mortality in incident dialysis patients. J. Am. Heart Assoc. 2013, 2, 1–11. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Ginseng | ||
|---|---|---|---|
| GL100 mg/kg/day | GH200 mg/kg/day | ||
| Body weight gain (g) | 129.70 ± 15.74 | 123.90 ± 9.60 | 136.60±19.72 |
| Food intake (g/day) | 23.26 ± 0.66 | 23.98 ± 1.09 | 24.65 ± 1.15 |
| Adipose tissue (g) | 3.53 ± 0.40 | 3.43 ± 0.43 | 3.73 ± 0.59 |
| Kidney (g) | 1.11 ± 0.10 | 1.18 ± 0.09 | 1.14 ± 0.13 |
| Liver (g) | 10.53 ± 1.18 | 10.76 ± 0.98 | 11.06 ± 1.63 |
| TG (mg/dL) | 27.71 ± 1.70 | 24.00 ± 3.81 | 24.43 ± 4.47 |
| TC (mg/dL) | 122.17 ± 6.97 b | 116.8 ± 4.15 b | 101.17 ± 2.32 a |
| HDL (mg/dL) | 53.50 ± 3.02 b | 53.6 ± 2.30 b | 38.17 ± 3.43 a |
| LDL (mg/dL) | 64.37 ± 9.96 b | 58.4 ± 3.71 ab | 54.63 ± 3.90 a |
| oxLDL (ng/dL) | 17.48 ± 3.12 b | 10.35 ± 0.38 a | 10.56 ± 0.34 a |
| oxLDL/HDL | 0.33 ± 0.07 b | 0.19 ± 0.01 a | 0.28 ± 0.03 b |
| oxLDL/LDL | 0.28 ± 0.05 b | 0.18 ± 0.01 a | 0.19 ± 0.01 a |
| oxLDL/TC | 0.14 ± 0.02 b | 0.09 ± 0.00 a | 0.10 ± 0.01 a |
| TC/HDL | 2.29 ± 0.24 | 2.18 ± 0.10 | 2.67 ± 0.24 |
| Metabolite | p-Value a | VIP b | Fold Change (vs. Control) | ||
|---|---|---|---|---|---|
| GL | GH | ||||
| Kidney | uracil | 1.83 × 102 | 0.94 | −1.14 | −1.19 |
| glutathione, oxidized | 5.64 × 104 | 1.40 | −2.78 | −2.87 | |
| succinyladenosine | 1.23 × 102 | 0.95 | −1.50 | −1.00 | |
| riboflavin | 2.00 × 103 | 1.21 | −1.01 | 1.35 | |
| leucyl histidine | 2.51 × 105 | 1.45 | −1.02 | −1.50 | |
| stearoylcarnitine | 2.70 × 102 | 0.96 | 1.47 | 1.60 | |
| Liver | carnitine | 3.30 × 102 | 1.10 | 1.06 | 1.46 |
| LPE(C20:4) | 2.40 × 103 | 2.54 | 1.16 | 1.57 | |
| LPC(C18:1) | 2.24 × 102 | 1.58 | 1.21 | −1.33 | |
| Plasma | LPE(C18:3) | 7.05 × 104 | 0.69 | −2.18 | −2.66 |
| LPC(C16:1) | 1.04 × 104 | 0.55 | −2.19 | −3.97 | |
| LPC(C20:4) | 7.56 × 105 | 1.00 | −2.20 | −4.50 | |
| LPC(C18:2) | 1.46 × 104 | 2.48 | −2.16 | −4.62 | |
| LPC(C18:3) | 1.23 × 103 | 0.17 | −2.32 | −3.39 | |
| LPC(C16:0) | 4.12 × 105 | 3.28 | −2.15 | −3.93 | |
| LPC(C18:1) | 2.12 × 104 | 1.88 | −2.76 | −6.58 | |
| LPC(C17:0) | 3.55 × 105 | 0.49 | −3.20 | −7.34 | |
| LPC(C18:0) | 5.20 × 105 | 2.41 | −3.03 | −5.90 | |
| Estradiol-3-glucuronate | 1.08 × 102 | 0.96 | 1.10 | −1.03 | |
| estrone-3-hemisuccinate | 9.92 × 103 | 1.00 | 1.47 | −1.30 | |
| 11α-hydroxyestradiol | 2.62 × 1023 | 0.79 | 1.13 | 1.10 | |
| 21α-hydroxyprogesterone | 3.87 × 103 | 1.02 | 1.53 | −1.08 | |
| Urine | 27-hydroxycholesterol | 1.78 × 102 | 0.94 | −1.25 | 1.19 |
| metandienone | 3.76 × 102 | 0.78 | 1.26 | 1.00 | |
| 2-hydroxyestrone-1+4-N-acetylcysteine | 3.16 × 102 | 0.90 | 1.98 | 2.13 | |
| 6-dehydroestradiol diacetate | 2.71 × 102 | 0.95 | 1.13 | 1.33 | |
| estriol-16,17-diacetate | 2.60 × 102 | 0.82 | 2.14 | −1.01 | |
| 2,3-dimethoxyestradiol | 3.57 × 102 | 0.85 | 1.48 | −1.62 | |
| 5-androstenediol | 4.17 × 102 | 0.86 | −1.42 | −1.43 | |
| 11β-hydroxyestradiol derivatives | 7.45 × 102 | 1.86 | 6.75 | 6.67 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-J.; Kim, B.-M.; Lee, S.H.; Sohn, J.-T.; Choi, J.W.; Cho, C.-W.; Hong, H.-D.; Rhee, Y.K.; Kim, H.-J. Ginseng-Induced Changes to Blood Vessel Dilation and the Metabolome of Rats. Nutrients 2020, 12, 2238. https://doi.org/10.3390/nu12082238
Lee H-J, Kim B-M, Lee SH, Sohn J-T, Choi JW, Cho C-W, Hong H-D, Rhee YK, Kim H-J. Ginseng-Induced Changes to Blood Vessel Dilation and the Metabolome of Rats. Nutrients. 2020; 12(8):2238. https://doi.org/10.3390/nu12082238
Chicago/Turabian StyleLee, Hyeon-Jeong, Bo-Min Kim, Soo Hee Lee, Ju-Tae Sohn, Jae Woong Choi, Chang-Won Cho, Hee-Do Hong, Young Kyoung Rhee, and Hyun-Jin Kim. 2020. "Ginseng-Induced Changes to Blood Vessel Dilation and the Metabolome of Rats" Nutrients 12, no. 8: 2238. https://doi.org/10.3390/nu12082238
APA StyleLee, H.-J., Kim, B.-M., Lee, S. H., Sohn, J.-T., Choi, J. W., Cho, C.-W., Hong, H.-D., Rhee, Y. K., & Kim, H.-J. (2020). Ginseng-Induced Changes to Blood Vessel Dilation and the Metabolome of Rats. Nutrients, 12(8), 2238. https://doi.org/10.3390/nu12082238