Separating the Wheat from the Chaff: Nutritional Value of Plant Proteins and Their Potential Contribution to Human Health



Abstract
:1. Introduction
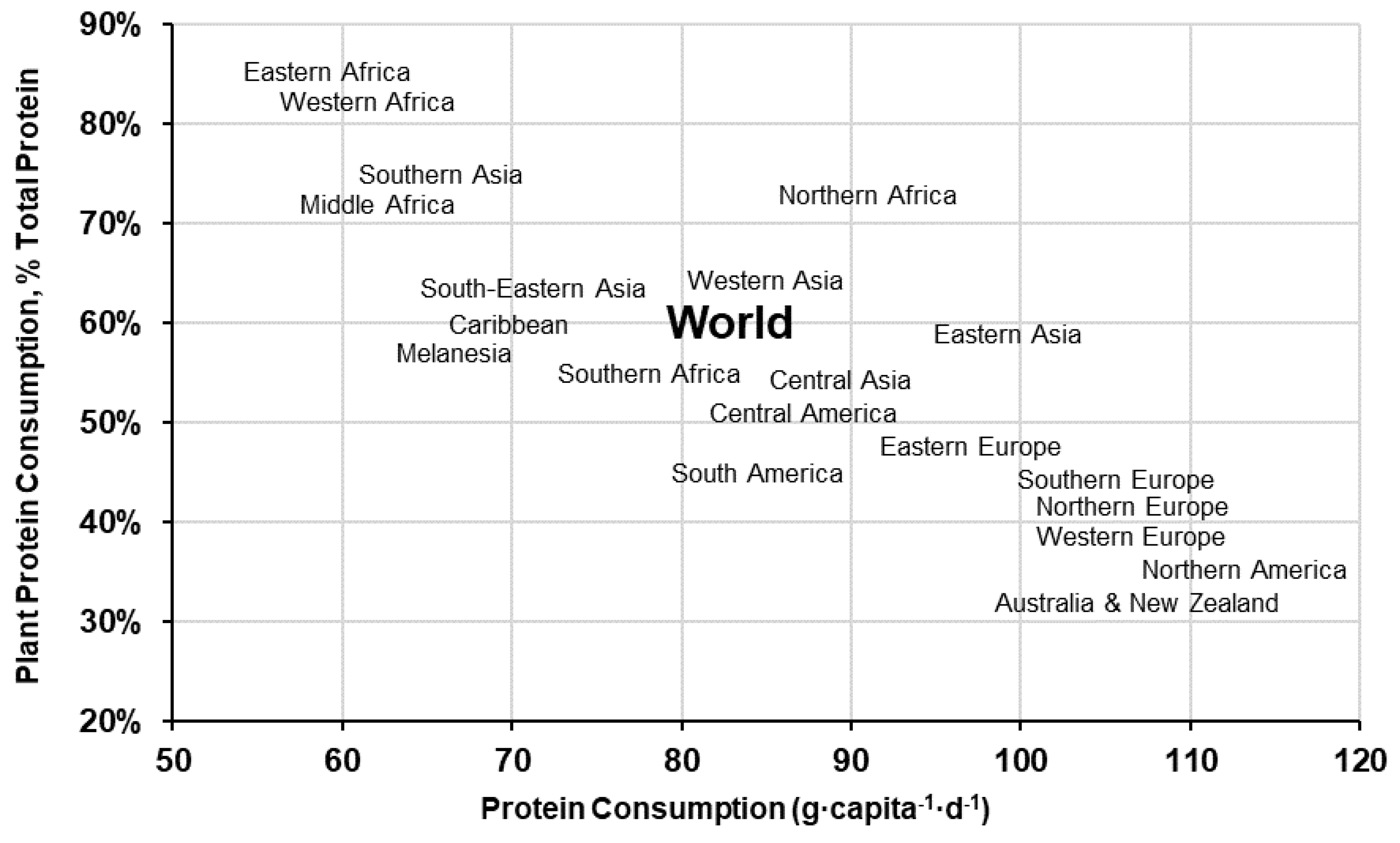
2. Plant Protein Consumption: Trends and Patterns
3. Human Protein and Amino Acid Requirements
4. Protein Quality
5. Protein Complementation
6. Timing and Distribution
7. Digestibility and Bioavailability
8. Genetic Engineering and Selective Breeding
9. Sustainability and Nutrition
10. Summary and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Millward, D.J. Metabolic Demands for Amino Acids and the Human Dietary Requirement: Millward and Rivers (1988) Revisited. J. Nutr. 1998, 128, 2563–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomé, D.; Bos, C. Dietary protein and nitrogen utilization. J. Nutr. 2000, 130, 1868–1873. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R.R.; Baum, J.I.; Starck, C.; Moughan, P.J. Factors contributing to the selection of dietary protein food sources. Clin. Nutr. 2018, 37, 130–138. [Google Scholar] [CrossRef]
- World Health Organisation (WHO); Food Agriculture Organisation (FAO) of the United Nations. Protein and Amino Acid Requirements in Human Nutrition—Report of A Joint FAO/WHO/UNU Expert Consultation; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2017: Building Resilience for Peace and Food Security; FAO: Rome, Italy, 2017; Available online: http://www.fao.org/3/a-i7695e.pdf (accessed on 4 May 2020).
- Ghosh, S.; Suri, D.; Uauy, R. Assessment of protein adequacy in developing countries: Quality matters. Br. J. Nutr. 2012, 108, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. The impact of dietary protein intake on longevity and metabolic health. EBioMedicine 2019, 43, 632–640. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT: Food Balance Sheets. Available online: http://www.fao.org/faostat/en/#data/FBS (accessed on 4 May 2020).
- Food and Agriculture Organization of the UN; International Fund for Agricultural Development; UNICEF; World Food Programme; WHO. The State of Food Security and Nutrition in the World; Food and Agriculture Organization of the UN: Rome, Italy, 2018. [Google Scholar]
- Pimentel, D.; Pimentel, M. Sustainability of meat-based and plant-based diets and the environment. Am. J. Clin. Nutr. 2003, 78, 660–663. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; Declerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- IPCC. Climate Change and Land; The Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2019. [Google Scholar]
- World Health Organization (WHO). Sustainable Healthy Diets: Guiding Principles; Food & Agriculture Org: Rome, Italy, 2019. [Google Scholar]
- Hirvonen, K.; Bai, Y.; Headey, D.; Masters, W.A. Affordability of the EAT–Lancet reference diet: A global analysis. Lancet Glob. Health 2020, 8, e59–e66. [Google Scholar] [CrossRef] [Green Version]
- Ridoutt, B.; Hendrie, G.A.; Noakes, M. Dietary Strategies to Reduce Environmental Impact: A Critical Review of the Evidence Base. Adv. Nutr. 2017, 8, 933–946. [Google Scholar] [CrossRef] [Green Version]
- Adesogan, A.T.; Havelaar, A.H.; McKune, S.L.; Eilittä, M.; Dahl, G.E. Animal source foods: Sustainability problem or malnutrition and sustainability solution? Perspective matters. Glob. Food Secur. 2020, 25. [Google Scholar] [CrossRef]
- Young, V.R.; Pellett, P.L. Plant proteins in relation to human protein and amino acid nutrition. Am. J. Clin. Nutr. 1994, 59, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rand, W.M.; Pellett, P.L.; Young, V.R. Meta-analysis of nitrogen balance studies for estimating protein requirements in healthy adults. Am. J. Clin. Nutr. 2003, 77, 109–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pencharz, P.B.; Elango, R.; Wolfe, R.R. Recent developments in understanding protein needs—How much and what kind should we eat? Appl. Physiol. Nutr. Metab. 2016, 41, 577–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traylor, D.A.; Gorissen, S.H.; Phillips, S.M. Perspective: Protein Requirements and Optimal Intakes in Aging: Are We Ready to Recommend More Than the Recommended Daily Allowance? Adv. Nutr. 2018, 9, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layman, D.K.; Anthony, T.G.; Rasmussen, B.B.; Adams, S.H.; Lynch, C.J.; Brinkworth, G.D.; Davis, T.A. Defining meal requirements for protein to optimize metabolic roles of amino acids. Am. J. Clin. Nutr. 2015, 101, 1330–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition: Report of an FAO Expert Consultation; FAO Food Nutr Pap.: Rome, Italy, 2013. [Google Scholar]
- Wolfe, R.R.; Rutherfurd, S.M.; Kim, I.-Y.; Moughan, P.J. Protein quality as determined by the Digestible Indispensable Amino Acid Score: Evaluation of factors underlying the calculation. Nutr. Rev. 2016, 74, 584–599. [Google Scholar] [CrossRef]
- Van Der Spiegel, M.; Noordam, M.Y.; Van Der Fels-Klerx, H.J. Safety of Novel Protein Sources (Insects, Microalgae, Seaweed, Duckweed, and Rapeseed) and Legislative Aspects for Their Application in Food and Feed Production. Compr. Rev. Food Sci. Food Saf. 2013, 12, 662–678. [Google Scholar] [CrossRef]
- United States Department of Agriculture. FoodData Central. Available online: https://fdc.nal.usda.gov/ (accessed on 4 May 2020).
- Young, V.R.; Scrimshaw, N.S.; Torún, B.; Viteri, F. Soybean protein in human nutrition: An overview. J. Am. Oil Chem. Soc. 1979, 56, 110–120. [Google Scholar] [CrossRef]
- Pellett, P.L.; Ghosh, S. Lysine Fortification: Past, Present, and Future. Food Nutr. Bull. 2004, 25, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Gunaratna, N.S.; De Groote, H.; Nestel, P.; Pixley, K.V.; McCabe, G.P. A meta-analysis of community-based studies on quality protein maize. Food Policy 2010, 35, 202–210. [Google Scholar] [CrossRef]
- Ghosh, S.; Smriga, M.; Vuvor, F.; Suri, D.; Mohammed, H.; Armah, S.M.; Scrimshaw, N.S. Effect of lysine supplementation on health and morbidity in subjects belonging to poor peri-urban households in Accra, Ghana. Am. J. Clin. Nutr. 2010, 92, 928–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervantes-Pahm, S.K.; Liu, Y.; Stein, H.H. Digestible indispensable amino acid score and digestible amino acids in eight cereal grains. Br. J. Nutr. 2014, 111, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, N.; Islam, S.; Munmun, S.; Mohiduzzaman, M.; Longvah, T. Amino acid profiles and digestible indispensable amino acid scores of proteins from the prioritized key foods in Bangladesh. Food Chem. 2016, 213, 83–89. [Google Scholar] [CrossRef]
- Mathai, J.K.; Liu, Y.; Stein, H.H. Values for digestible indispensable amino acid scores (DIAAS) for some dairy and plant proteins may better describe protein quality than values calculated using the concept for protein digestibility-corrected amino acid scores (PDCAAS). Br. J. Nutr. 2017, 117, 490–499. [Google Scholar] [CrossRef]
- Han, F.; Han, F.; Wang, Y.; Fan, L.; Song, G.; Chen, X.; Jiang, P.; Miao, H.; Han, Y. Digestible indispensable amino acid scores of nine cooked cereal grains. Br. J. Nutr. 2019, 121, 30–41. [Google Scholar] [CrossRef]
- Moughan, P.J.; Gilani, S.; Rutherfurd, S.M.; Tomé, D. True Ileal Amino Acid Digestibility Coefficients for Application in the Calculation Digestible Indispensable Amino Acid Score (DIAAS) in Human Nutrition. Available online: http://www.fao.org/ag/humannutrition/36216-04a2f02ec02eafd4f457dd2c9851b4c45.pdf (accessed on 4 May 2020).
- Rutherfurd, S.M.; Fanning, A.C.; Miller, B.J.; Moughan, P.J. Protein Digestibility-Corrected Amino Acid Scores and Digestible Indispensable Amino Acid Scores Differentially Describe Protein Quality in Growing Male Rats. J. Nutr. 2014, 145, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Nosworthy, M.G.; Neufeld, J.; Frohlich, P.; Young, G.; Malcolmson, L.; House, J.D. Determination of the protein quality of cooked Canadian pulses. Food Sci. Nutr. 2017, 5, 896–903. [Google Scholar] [CrossRef]
- Bergström, J.; Fürst, P.; Vinnars, E. Effect of a Test Meal, without and with Protein, on Muscle and Plasma Free Amino Acids. Clin. Sci. 1990, 79, 331–337. [Google Scholar] [CrossRef]
- Mamerow, M.M.; Mettler, J.A.; English, K.L.; Casperson, S.L.; Arentson-Lantz, E.; Sheffield-Moore, M.; Layman, D.K.; Paddon-Jones, D. Dietary protein distribution positively influences 24-h muscle protein synthesis in healthy adults. J. Nutr. 2014, 144, 876–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, S.M.; Chevalier, S.; Leidy, H.J. Protein “requirements” beyond the RDA: Implications for optimizing health. Appl. Physiol. Nutr. Metab. 2016, 41, 565–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorissen, S.; Horstman, A.M.; Franssen, R.; Crombag, J.J.; Langer, H.T.; Bierau, J.; Respondek, F.; Van Loon, L. Ingestion of Wheat Protein Increases In Vivo Muscle Protein Synthesis Rates in Healthy Older Men in a Randomized Trial. J. Nutr. 2016, 146, 1651–1659. [Google Scholar] [CrossRef]
- Monteyne, A.J.; Coelho, M.O.; Porter, C.; Abdelrahman, D.R.; Jameson, T.S.; Jackman, S.R.; Blackwell, J.R.; Finnigan, T.J.; Stephens, F.B.; Dirks, M.L.; et al. Mycoprotein ingestion stimulates protein synthesis rates to a greater extent than milk protein in rested and exercised skeletal muscle of healthy young men: A randomized controlled trial. Am. J. Clin. Nutr. 2020, 112, 318–333. [Google Scholar] [CrossRef] [PubMed]
- Mongeau, R.; Sarwar, G.; Peace, R.W.; Brassard, R. Relationship between dietary fiber levels and protein digestibility in selected foods as determined in rats. Plant Foods Hum. Nutr. 1989, 39, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Gilani, G.S.; Xiao, C.W.; Cockell, K.A. Impact of Antinutritional Factors in Food Proteins on the Digestibility of Protein and the Bioavailability of Amino Acids and on Protein Quality. Br. J. Nutr. 2012, 108, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Sá, A.G.; Moreno, Y.M.; Carciofi, B.A. Food processing for the improvement of plant proteins digestibility. Crit. Rev. Food Sci. Nutr. 2019, 1–20. [Google Scholar] [CrossRef]
- Nagra, S.A.; Bhatty, N. In vivo (rat assay) assessment of nutritional improvement of peas (Pisum sativum L.). East. Mediterr. Health J. 2007, 13, 646–653. [Google Scholar]
- Pasini, G.; Simonato, B.; Giannattasio, M.; Peruffo, A.D.; Curioni, A. Modifications of wheat flour proteins during in vitro digestion of bread dough, crumb, and crust: An electrophoretic and immunological study. J. Agric. Food Chem. 2001, 49, 2254–2261. [Google Scholar] [CrossRef]
- Turgeon, S.L.; Rioux, L.-E. Food matrix impact on macronutrients nutritional properties. Food Hydrocoll. 2011, 25, 1915–1924. [Google Scholar] [CrossRef]
- Mertz, E.T.; Bates, L.S.; Nelson, O.E. Mutant Gene That Changes Protein Composition and Increases Lysine Content of Maize Endosperm. Science 1964, 145, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Amir, R. Fortifying plants with the essential amino acids lysine and methionine to improve nutritional quality. Plant Biotechnol. J. 2013, 11, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Gambhir, G.; Dass, A.; Tripathi, A.K.; Singh, A.; Jha, A.K.; Yadava, P.; Choudhary, M.; Rakshit, S. Genetically modified crops: Current status and future prospects. Planta 2020, 251, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Z.-B.; Xing, A.; Moon, B.P.; Koellhoffer, J.P.; Huang, L.; Ward, R.T.; Clifton, E.; Falco, S.C.; Cigan, A.M. Cas9-Guide RNA Directed Genome Editing in Soybean. Plant Physiol. 2015, 169, 960–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessari, P.; Lante, A.; Mosca, G. Essential amino acids: Master regulators of nutrition and environmental footprint? Sci. Rep. 2016, 6, 26074. [Google Scholar] [CrossRef] [Green Version]
- Ertl, P.; Knaus, W.; Zollitsch, W. An approach to including protein quality when assessing the net contribution of livestock to human food supply. Animal 2016, 10, 1883–1889. [Google Scholar] [CrossRef] [Green Version]
- Clune, S.; Crossin, E.; Verghese, K. Systematic review of greenhouse gas emissions for different fresh food categories. J. Clean. Prod. 2017, 140, 766–783. [Google Scholar] [CrossRef] [Green Version]
- Brundtland Commission. Our Common Future; Oxford University Press: Oxford, UK, 1987. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Plant Source | Global Supply % | Protein % Mass | CP True Ileal Digestibility % | DIAAS a | |||
|---|---|---|---|---|---|---|---|
| Leu | Lys | SAA | Trp | ||||
| Wheat [24,27,32,33,34,35,36] | 32 | 11–17 | 71–94 | 0.82–1.06 | 0.20–0.54 | 0.64–1.51 | 1.15–1.62 |
| Rice [27,32,33,35,36,37] | 21 | 8–9 | 73–90 | 0.84–1.17 | 0.37–0.73 | 0.40–2.11 | 0.84–2.29 |
| Maize [27,32,36,37] | 8 | 7–9 | 70–76 | 1.31–2.01 | 0.48–0.54 | 0.68–1.46 | 0.70–1.04 |
| Pulses [27,33,34,36,37,38] | 5 | 22–30 | 78–90 | 0.97–1.16 | 1.05–1.53 | 0.46–0.85 | 0.78–1.82 |
| Beans [27,36,37,38] | 3 | 23–25 | 58–83 | 0.72–1.09 | 0.93–0.98 | 0.49–0.60 | 0.76–1.86 |
| Potatoes [36,27] | 3 | 2–3 | 52–58 | 0.34–0.39 | 0.42–0.46 | 0.38–0.77 | 0.49–1.42 |
| Soya [27,34,36,37] | 3 | 12–43 | 68–88 | 1.14–1.39 | 1.10–1.25 | 0.93–1.12 | 2.04–2.11 |
| Sorghum [27,33,36] | 2 | 10–11 | 65–83 | 1.58–1.79 | 0.26–0.29 | 0.54–0.97 | 0.57–1.05 |
| Groundnuts [27,36,37] | 2 | 26 | 77–91 | 0.84–0.94 | 0.38–0.52 | 0.58–0.98 | 0.74–1.58 |
| Millet [27,35,36] | 1 | 8–17 | 80–90 | 1.70–1.73 | 0.07–0.10 | 0.62–1.17 | 0.77–1.82 |
| WPC [27,34,36,37] | N/A | 80–85 | 95–98 | 1.91–1.98 | 1.80–2.41 | 1.99–2.00 | 2.99–3.40 |
| WMP [24,27,36] | N/A | 28 | 96 | 1.62 | 1.54 | 1.43 | 1.82 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davies, R.W.; Jakeman, P.M. Separating the Wheat from the Chaff: Nutritional Value of Plant Proteins and Their Potential Contribution to Human Health. Nutrients 2020, 12, 2410. https://doi.org/10.3390/nu12082410
Davies RW, Jakeman PM. Separating the Wheat from the Chaff: Nutritional Value of Plant Proteins and Their Potential Contribution to Human Health. Nutrients. 2020; 12(8):2410. https://doi.org/10.3390/nu12082410
Chicago/Turabian StyleDavies, Robert W., and Philip M. Jakeman. 2020. "Separating the Wheat from the Chaff: Nutritional Value of Plant Proteins and Their Potential Contribution to Human Health" Nutrients 12, no. 8: 2410. https://doi.org/10.3390/nu12082410