Frozen Autoclaved Sorghum Enhanced Colonic Fermentation and Lower Visceral Fat Accumulation in Rats


 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of Experimental Diets
2.2. Animal Experimental Design, Care for Laboratory Animals and Post-Mortem Excision of Organs
2.3. DNA Extraction, Next-Generation Sequencing (NGS) and the Analysis of 16S Ribosomal RNA (16S rRNA) Gene Sequences in Cecal Content
2.4. Rat Cecal SCFA Analysis by High Performance Liquid Chromatography (HPLC)
2.5. Fecal and Liver Lipid Analysis
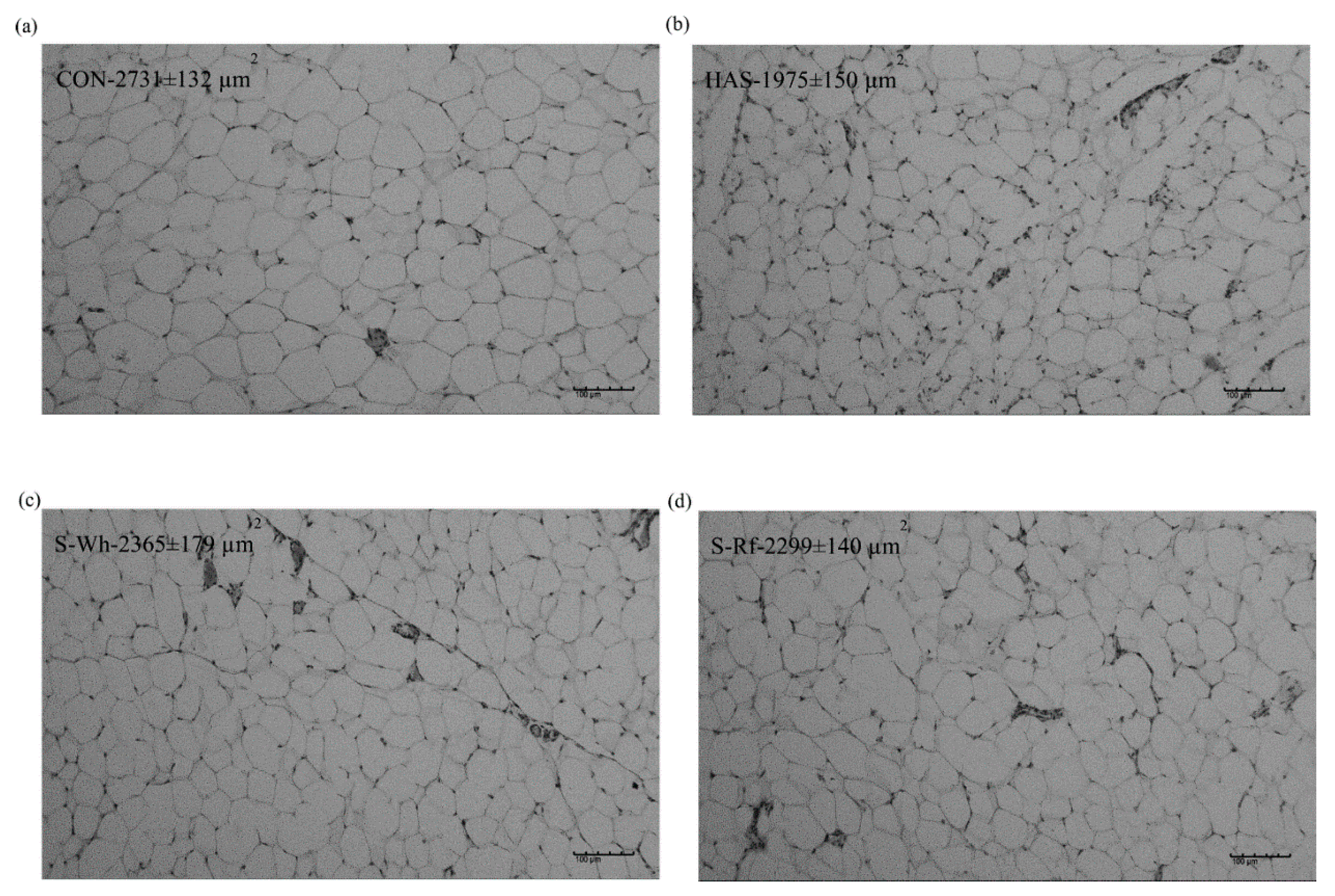
2.6. Mesenteric Adipocyte Staining
2.7. Serum Biochemical Analysis
2.8. Statistical Analysis
3. Results
3.1. Zoometric Parameters, Feed Intake and Organ Weights
3.2. Microbial Community DNA Data
3.2.1. Alpha and Beta Diversities
3.2.2. Taxonomic Diversity
3.3. Cecal Short Chain Fatty Acid Content and pH
3.4. Liver Lipid Profile, Fecal Moisture and Dry Matter Contents, Lipid and Bile Acid Profiles
3.4.1. Liver Lipid Profile
3.4.2. Fecal Moisture, Dry Matter, Lipid and Bile Acid Profiles
3.5. Serum Lipid Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, X.; Shimizu, Y.; Kimura, I. Gut microbial metabolite short-chain fatty acids and obesity. Biosci. Microbiota Food Heal. 2017, 36, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, H.; Maruta, Y.H.; Jozuka, M.; Kimura, R.; Iwabuchi, H.; Yamato, M.; Saito, T.; Fujisawa, K.; Takahashi, Y.; Kimoto, Y.; et al. Effects of acetate on lipid metabolism in muscles and adipose tissues of type 2 diabetic otsuka long-evans tokushima fatty (OLETF) Rats. Biosci. Biotechnol. Biochem. 2009, 73, 570–576. [Google Scholar] [CrossRef] [Green Version]
- Kasubuchi, N.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [Green Version]
- Tsuiki, K.; Fujisawa, H.; Itoh, A.; Sato, M.; Fujita, N. Alterations of starch structure lead to increased resistant starch of steamed rice: Identification of high resistant starch rice lines. J. Cereal Sci. 2016, 68, 88–92. [Google Scholar] [CrossRef]
- Li, X.; Watanabe, K.; Kimura, I. Gut microbiota dysbiosis drives and implies novel therapeutic strategies for diabetes mellitus and related metabolic diseases. Front. Immunol. 2017, 8, 1–7. [Google Scholar] [CrossRef]
- Dugas, L.R.; Lie, L.; Plange-Rhule, J.; Bedu-Addo, K.; Bovet, P.; Lambert, E.V.; Forrester, T.E.; Luke, A.; Gilbert, J.A.; Layden, B.T. Gut microbiota, short chain fatty acids, and obesity across the epidemiologic transition: The METS-Microbiome study protocol. BMC Public Health 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.-J.; Liu, Q.; Hoover, R. Impact of annealing and heat-moisture treatment on rapidly digestible, slowly digestible and resistant starch levels in native and gelatinized corn, pea and lentil starches. Carbohydr. Polym. 2009, 75, 436–447. [Google Scholar] [CrossRef]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant starch: A review. Compr. Rev. Food Sci. Food Saf. 2006, 5, 1–17. [Google Scholar] [CrossRef]
- Tovar, J.; Melito, C. Steam-cooking and dry heating produce resistant starch in legumes. J. Agric. Food Chem. 1996, 44, 2642–2645. [Google Scholar] [CrossRef]
- Eerlingen, R.C.; Crombez, M.; Delcour, J.A. Enzyme-resistant starch. I. Quantitative and qualitative influence of incubation time and temperature of autoclaved starch on resistant starch formation. Cereal Chem. 1993, 70, 339–344. [Google Scholar]
- Haralampu, S.G. Resistant starch—A review of the physical properties and biological impact of RS 3. Carbohydr. Polym. 2000, 41, 285–292. [Google Scholar] [CrossRef]
- Zhu, F. Structure, Physicochemical Properties, Modifications and Uses of Sorghum Starch. Compr. Rev. Food Sci. Food Saf. 2014, 13, 597–610. [Google Scholar] [CrossRef]
- Teixeira, N.D.C.; Queiroz, V.A.V.; Rocha, M.C.; Amorim, A.C.P.; Soares, T.O.; Monteiro, M.A.M.; de Menezes, C.B.; Schaffert, R.E.; Garcia, M.A.V.T.; Junnqueira, R.G. Resistant starch content among several sorghum (Sorghum bicolor) genotypes and the effect of heat treatment on resistant starch retention in two genotypes. Food Chem. 2016, 197, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelpolage, S.W.; Han, K.-H.; Koaze, H.; Hamamoto, T.; Hoshizawa, M.; Fukushima, M. Influence of enzyme-resistant fraction of sorghum (Sorghum bicolor L.) flour on gut microflora composition, short chain fatty acid production and toxic substance metabolism. J. Food Nutr. Res. 2019, 58, 135–145. [Google Scholar]
- Pelpolage, S.W.; Nagata, R.; Fukuma, N.; Shimada, K.; Han, K.-H.; Hamamoto, T.; Hoshizawa, M.; Fukushima, M. In vivo colonic fermentation of sorghum (Sorghum bicolor L.): Important correlations observed among the physiological parameters of cecum, liver, adipose tissue and fasting serum lipid profile. J. Nutr. Sci. Vitaminol. 2019, 65, S222–S227. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.H.; Bean, S.; Hsieh, C.F.; Shi, Y.-C. Changes in protein and starch digestibility in sorghum flour during heat–moisture treatments. J. Sci. Food Agric. 2017, 97, 4770–4779. [Google Scholar] [CrossRef] [PubMed]
- Emmambux, M.N.; Taylor, J.R.N. Properties of heat-treated sorghum and maize meal and their prolamin proteins. J. Agric. Food Chem. 2009, 57, 1045–1050. [Google Scholar] [CrossRef]
- Shen, R.L.; Zhang, W.L.; Dong, J.L.; Ren, G.-X.; Chen, M. Sorghum resistant starch reduces adiposity in high-fat diet-induced overweight and obese rats via mechanisms involving adipokines and intestinal flora. Food Agric. Immunol. 2015, 26, 120–130. [Google Scholar] [CrossRef]
- Yun, S.; Matheson, N. Estimation of amylose content of starches after precipitation of amylopectin by concanavalin-A. Starch-Stärke 1990, 42, 302–305. [Google Scholar] [CrossRef]
- Blevins, J.E.; Overduin, J.; Fuller, J.M.; Cummings, D.E.; Matsumoto, K.; Moralejo, D.H. Normal feeding and body weight in Fishcer 344 rats lacking the cholecystokinin-1 receptor gene. Brain Res. 2009, 1255, 98–112. [Google Scholar] [CrossRef]
- Zhou, A.L.; Hintze, K.J.; Jimenez-Flores, R.; Ward, R.E. Dietary fat composition influences tissue lipid profile and gene expression in fisher-344 rats. Lipids 2012, 47, 1119–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Héliès, J.-M.; Diane, A.; Langlois, A.; Larue-Achagiotis, C.; Fromentin, G.; Tomé, D.; Mormède, P.; Marissal-Arvy, N. Composition of fat storage between fischer 344 and obesity-resistant Lou/C rats fed different diets. Obes. Res. 2005, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.N.; Edmondson, J.; Hildebrandt, P.K.; Bruner, R.H. Influence of dietary protein, fat, and fiber on growth, blood chemistry, and tumor incidences in fisher 344 rats. Nutr. Cancer 1996, 25, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Pelpolage, S.W.; Goto, Y.; Nagata, R.; Fukuma, N.; Furuta, T.; Mizu, M.; Han, K.-H.; Fukushima, M. Colonic fermentation of water soluble fiber fraction extracted from sugarcane (Sacchurum officinarum L.) bagasse in murine models. Food Chem. 2019, 292, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Han, K.-H.; Lee, C.-H.; Kinoshita, M.; Oh, C.-H.; Shimada, K.; Fukushima, M. Spent turmeric reduces fat mass in rats fed a high-fat diet. Food Funct. 2016, 7, 1814–1824. [Google Scholar] [CrossRef]
- Warren, F.J.; Fukuma, N.M.; Mikkelsen, D.; Flanagan, B.M.; Williams, B.A.; Lisle, A.T.; Gidley, M.J. Food starch structure impacts gut microbiome composition. Mol. Biol. Physiol. 2018, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Sloane, S.G. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Han, K.-H.; Tsuchihira, H.; Nakamura, Y.; Simada, K.; Ohba, K.; Aritsuka, T.; Uchino, H.; Kikuchi, H.; Fukushima, M. Inulin-type fructans with different degrees of polymerization improve lipid metabolism but not glucose metabolism in rats fed a high-fat diet under energy restriction. Dig. Dis. Sci. 2013, 58, 2177–2186. [Google Scholar] [CrossRef]
- Matsubara, Y.; Sawabe, A.; Iizuka, Y. Structures of new limonoid glycosides in lemon (Citrus limon burm. f.) peelings. Agric. Biol. Chem. 1990, 54, 1143–1148. [Google Scholar]
- Nagata, R.; Kamibayashi, R.; Bochimoto, H.; Fukuma, N.; Shimada, K.; Tchibe, M.; Takaishi, Y.; Han, K.-H.; Fukushima, M. Chemical modification of cornstarch by hydroxypropylation enhances caecal fermentation-mediated lipid metabolism in rats. Starch-Stärke 2020, 72, 1–7. [Google Scholar] [CrossRef]
- Institute of Medicine. Weight Management: State of the Science and Opportunities for Military Programs; The National Academic Press: Washington, DC, USA, 2003; pp. 57–78. [Google Scholar]
- Lu, Y.; Fan, C.; Li, P.; Lu, Y.; Chang, X.; Qi, K. Short chain fatty acids prevent high-fat-diet-induced obesity in mice by regulating g protein-coupled receptors and gut microbiota. Sci. Rep. 2016, 6, 1–13. [Google Scholar]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Piza, A.; Lee, S.-J. Effects of dietary fibers and prebiotics in adiposity regulation via modulation of gut microbiota. Appl. Biol. Chem. 2020, 63, 1–12. [Google Scholar] [CrossRef]
- Hernández, M.A.G.; Canfora, E.E.; Jocken, J.W.E.; Blaak, E.E. The short-chain fatty acid acetate in body weight control and insulin sensitivity. Nutrients 2019, 11, 1943. [Google Scholar] [CrossRef] [Green Version]
- Schoeler, M.; Caesar, R. Dietary lipids, gut microbiota and lipid metabolism. Rev. Endo. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fechner, A.; Kiehntopf, M.; Jahreis, G. The formation of short-chain fatty acids is positively associated with the blood lipid lowering effect of Lupin kernel fiber in moderately hypercholesterolemic adults. J. Nutr. 2014, 144, 599–607. [Google Scholar] [CrossRef]
- Chen, X.; He, X.; Fu, X.; Huang, Q. In vitro digestion and physicochemical properties of wheat starch/flour modified by heat-moisture treatment. J. Cereal Sci. 2015, 63, 109–115. [Google Scholar] [CrossRef]
- Xiao, J.; Chen, L.; Johnson, S.; Yu, Y.; Zhang, X.; Chenn, J. Predictive modeling of microbiome data using phylogeny-regularized gerneralized linear mixed model. Front. Microbiol. 2018, 9, 1391–1405. [Google Scholar] [CrossRef] [Green Version]
- Castaner, O.; Goday, A.; Park, Y.-M.; Lee, S.-H.; Magkos, F.; Shiow, S.-A.T.E.; Schröder, H. The gut microbiome profile in obesity: A systematic review. Int. J. Endocrin. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Davis, C.D. The gut microbiome and its role in obesity. Nutr. Today 2016, 51, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, C.-H.; Wu, C.-Y. The gut microbiome in obesity. J. Formosan Med. Assoc. 2019, 118, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Bloc’h, J.L.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Grunfeld, C. Introduction to Lipids and Lipoproteins. Available online: https://www.ncbi.nlm.nih.gov/books/NBK305896/ (accessed on 5 June 2020).
- Kvit, K.; Kharchenko, V. Role of gut microbiota in lipid metabolism. Asian J. Pharm. Clin. Res. 2018, 11, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Mardones, P.; Quiñones, V.; Amigo, L.; Moreno, M.; Miquel, J.F.; Schwarz, M.; Miettinen, H.E.; Trigatti, B.; Krieger, M.; VanPatten, S.; et al. Hepatic cholesterol and bile acid metabolism and intestinal cholesterol absorption in scavenger receptor class B type I-deficient mice. J. Lipid Res. 2001, 42, 170–180. [Google Scholar]
- Ooi, L.-G.; Liong, M.-T. Cholesterol-lowering effects of probiotics and prebiotics: A review of in vivo and in vitro findings. Int. J. Mol. Sci. 2010, 11, 2499–2522. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Proia, A.; McNamara, D.J. Origins of neutral steroids in rats. J. Lipid Res. 1981, 22, 485–495. [Google Scholar]
- Huazano-García, A.; López, M.G. Metabolism of short chain fatty acids in the colon and faeces of mice after a supplementation of diets with agave fructans. In Lipid Metabolism; Baez, R.V., Ed.; IntechOpen Publications: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Hara, H. Physiological effects of short-chain fatty acid produced from prebiotics in the colon. Biosci. Microflora 2002, 21, 35–42. [Google Scholar] [CrossRef]
- Ghaffarzadegan, T.; Zhong, Y.; Hållenius, F.F.; Nyman, M. Effects of barley variety, dietary fiber and β-glucan content on bile acid composition in cecum of rats fed low- and high-fat diets. J. Nutr. Biochem. 2018, 53, 104–110. [Google Scholar] [CrossRef]
- Ellegård, L.; Andersson, H. Oat bran rapidly increases bile acid synthesis: An ileostomy study. Eur. J. Clin. Nutr. 2007, 61, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, A.F. The continuing importance of bile acids in liver and intestinal disease. Arch. Intern. Med. 1999, 159, 2647–2658. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dongowski, G.; Jacobasch, G.; Schhmiedl, D. Structural stability and prebiotic properties of resistant starch type 3 increase bile acid turnover and lower secondary bile acid formation. J. Agric. Food Chem. 2005, 53, 9257–9267. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | CON | HAS | S-Wh | S-Rf |
|---|---|---|---|---|
| Final body weight (g) | 250 ± 5 a | 228 ± 5 b | 241 ± 5 ab | 241 ± 3 ab |
| Body weight gain (g/30 days) | 62.0 ± 3.9 a | 40.8 ± 4.4 b | 54.0 ± 2.7 ab | 53.8 ± 2.8 ab |
| Feed intake (g/30 days) | 384 ± 6 a | 357 ± 6 b | 349 ± 7 b | 341 ± 7 b |
| Ep-AT (g/100 g body weight) | 2.57 ± 0.14 a | 1.71 ± 0.06 c | 2.17 ± 0.10 b | 2.12 ± 0.05 b |
| Pe-AT (g/100 g body weight) | 2.29 ± 0.07 a | 1.46 ± 0.09 c | 1.99 ± 0.12 b | 1.94 ± 0.10 b |
| Tv-AT (g/100 g body weight) | 4.86 ± 0.20 a | 3.17 ± 0.13 c | 4.17 ± 0.22 b | 4.06 ± 0.12 b |
| Cecal weight (g) | 2.32 ± 0.22 b | 5.80 ± 1.37 a | 1.99 ± 0.08 b | 2.14 ± 0.13 b |
| Cecal tissue weight (g) | 0.67 ± 0.02 b | 1.27 ± 0.09 a | 0.67± 0.05 b | 0.65 ± 0.02 b |
| Cecal content weight (g) | 1.65 ± 0.21 b | 4.52 ± 1.28 a | 1.31 ± 0.10 b | 1.48 ± 0.12 b |
| Parameter | CON | HAS | S-Wh | S-Rf |
|---|---|---|---|---|
| Acetate (µmol/cecum) | 118 ± 24 b | 271 ± 58 a | 98 ± 12 b | 152 ± 27 ab |
| Propionate (µmol/cecum) | 14.7 ± 2.3 ns | 29.2 ± 9.7 ns | 13.9 ± 2.0 ns | 19.0 ± 3.3 ns |
| n-Butyrate (µmol/cecum) | 2.76 ± 0.70 ab | 5.50 ± 0.67 a | 0.94 ± 0.19 b | 3.85 ± 1.25 ab |
| Total SCFA (µmol/cecum) | 136 ± 27 ab | 306 ± 67 a | 113 ± 14 b | 175 ± 31 ab |
| pH | 7.72 ± 0.10 a | 7.30 ± 0.11 b | 7.71 ± 0.06 a | 7.67 ± 0.11 ab |
| Parameter | CON | HAS | S-Wh | S-Rf |
|---|---|---|---|---|
| Liver lipid parameters | ||||
| Liver weight (g) | 5.91 ± 0.07 ns | 5.85 ± 0.22 ns | 5.80 ± 0.13 ns | 5.83 ± 0.08 ns |
| Liver total lipids (mg) | 305 ± 32 ns | 324 ± 26 ns | 430 ± 49 ns | 305 ± 40 ns |
| Liver cholesterol (mg) | 21.67 ± 1.32 ab | 19.32 ± 0.68 b | 20.91 ± 0.90 ab | 23.90 ± 0.88 a |
| Liver triglyceride (mg) | 56.18 ± 3.92 ns | 46.86 ± 3.26 ns | 56.23 ± 3.30 ns | 55.34 ± 3.42 ns |
| Fecal parameters | ||||
| Fecal dry weight (g) | 0.91 ± 0.03 ab | 1.16 ± 0.18 a | 0.77 ± 0.02 b | 0.76 ± 0.03 b |
| Fecal moisture content (%) | 54.7 ± 3.5 ns | 63.3 ± 4.6 ns | 59.6 ± 2.9 ns | 51.9 ± 2.3 ns |
| Fecal total lipids (mg/day) | 29.8 ± 4.8 ns | 58.6 ± 12.0 ns | 38.1 ± 8.8 ns | 27.7 ± 7.1 ns |
| Fecal neutral sterol (mg/day) | 2.09 ± 0.26 ns | 2.30 ± 0.42 ns | 1.69 ± 0.23 ns | 2.27 ± 0.23 ns |
| Fecal cholesterol (mg/day) | 0.90 ± 0.10 ns | 0.60 ± 0.13 ns | 0.75 ± 0.06 ns | 1.08 ± 0.10 ns |
| Fecal coprostanol (mg/day) | 0.98 ± 0.25 ns | 1.39 ± 0.43 ns | 0.77 ± 0.17 ns | 1.00 ± 0.08 ns |
| Fecal bile acid content (mg/day) | ||||
| Cholic acid | 0.38 ± 0.07 ns | 0.93 ± 0.23 ns | 0.85 ± 0.20 ns | 0.57 ± 0.13 ns |
| Chenodeoxycholic acid | <0.01 ns | 0.07 ± 0.04 ns | 0.01 ± 0.00 ns | <0.01 ns |
| Primary bile acid content | 0.46 ± 0.11 ns | 1.19 ± 0.30 ns | 1.04 ± 0.14 ns | 0.69 ± 0.19 ns |
| Deoxycholic acid | 0.46 ± 0.06 b | 0.34 ± 0.07 b | 0.79 ± 0.09 a | 0.56 ± 0.06 ab |
| Lithocholic acid | 0.61 ± 0.13 b | 0.41 ± 0.10 b | 0.92 ± 0.11 ab | 1.21 ± 0.34 a |
| Secondary bile acid content | 1.15 ± 0.21 ab | 0.81 ± 0.15 b | 1.85 ± 0.20 a | 1.88 ± 0.29 a |
| Total bile acid content | 1.61 ± 0.29 b | 2.01 ± 0.40 ab | 2.89 ± 0.32 a | 2.57 ± 0.31 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelpolage, S.W.; Yoshida, A.; Nagata, R.; Shimada, K.; Fukuma, N.; Bochimoto, H.; Hamamoto, T.; Hoshizawa, M.; Nakano, K.; Han, K.-H.; et al. Frozen Autoclaved Sorghum Enhanced Colonic Fermentation and Lower Visceral Fat Accumulation in Rats. Nutrients 2020, 12, 2412. https://doi.org/10.3390/nu12082412
Pelpolage SW, Yoshida A, Nagata R, Shimada K, Fukuma N, Bochimoto H, Hamamoto T, Hoshizawa M, Nakano K, Han K-H, et al. Frozen Autoclaved Sorghum Enhanced Colonic Fermentation and Lower Visceral Fat Accumulation in Rats. Nutrients. 2020; 12(8):2412. https://doi.org/10.3390/nu12082412
Chicago/Turabian StylePelpolage, Samanthi W., Atsushi Yoshida, Ryuji Nagata, Kenichiro Shimada, Naoki Fukuma, Hiroki Bochimoto, Tetsuo Hamamoto, Michiyo Hoshizawa, Koichi Nakano, Kyu-Ho Han, and et al. 2020. "Frozen Autoclaved Sorghum Enhanced Colonic Fermentation and Lower Visceral Fat Accumulation in Rats" Nutrients 12, no. 8: 2412. https://doi.org/10.3390/nu12082412