A Novel Dietary Intervention Reduces Circulatory Branched-Chain Amino Acids by 50%: A Pilot Study of Relevance for Obesity and Diabetes



 ,
, 
Abstract
1. Introduction
2. Methods
2.1. Study Ethics and Participants
2.2. Study Protocol
2.3. BCAA Restriction and Control Diet Plan Construction
2.4. Amino Acid Quantification
2.5. Plasma Glucose and Insulin
2.6. Statistical Analysis
3. Results
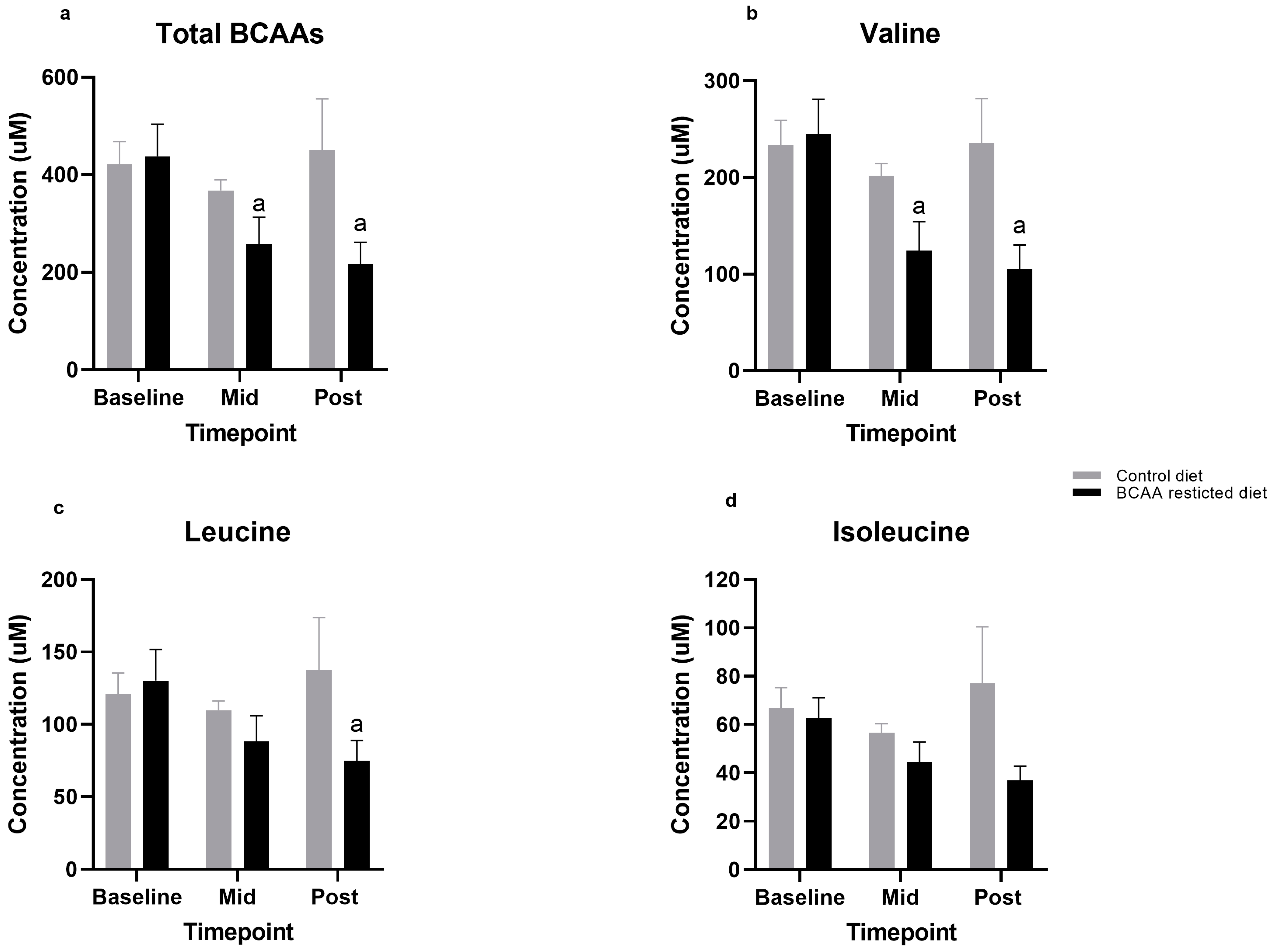
3.1. BCAA Levels
3.2. Individual Amino Acids
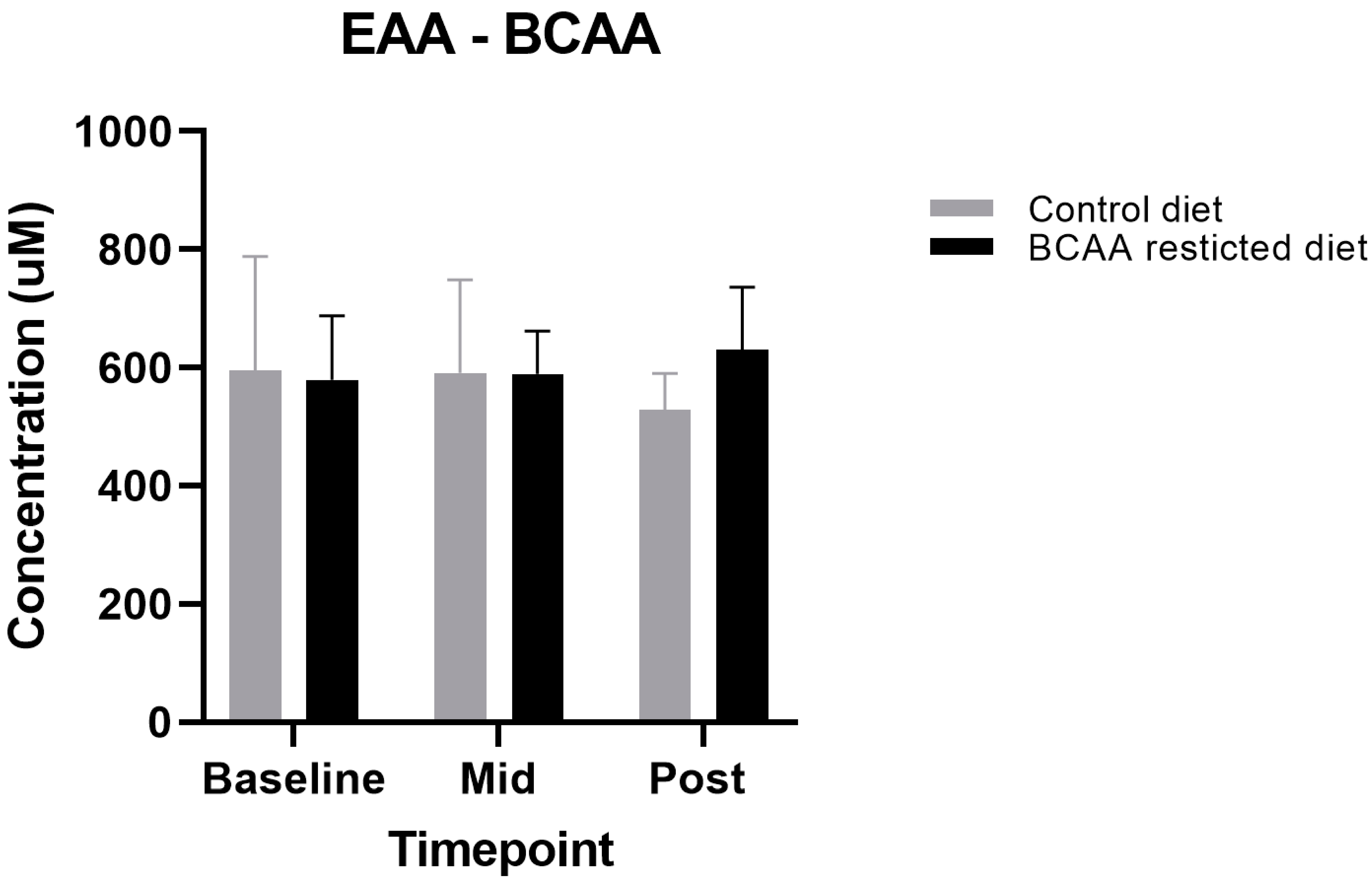
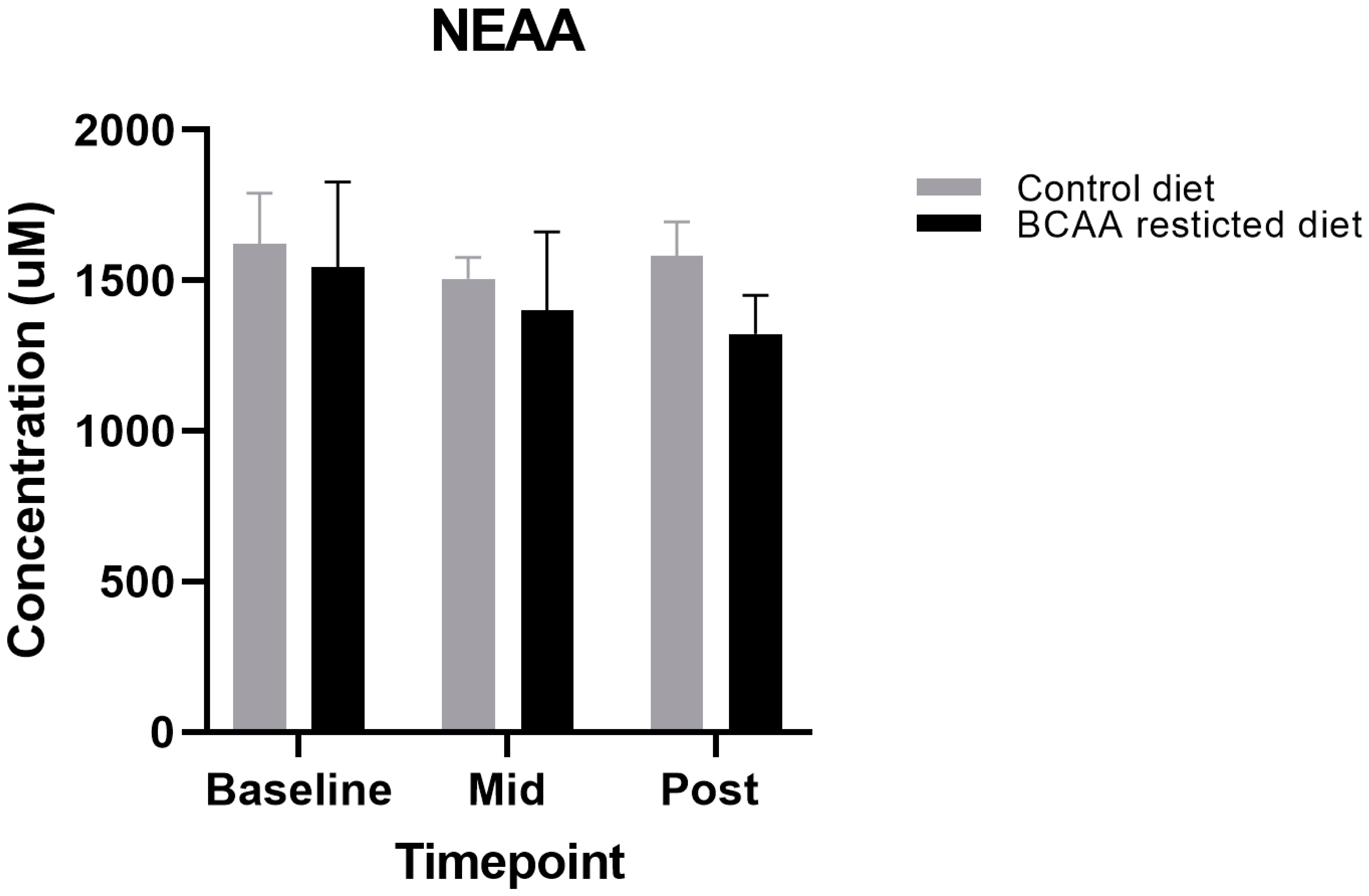
3.3. Other Amino Acids
3.4. Glucose, Insulin, and HOMA-IR Levels
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| T2D | Type 2 diabetes |
| IR | Insulin resistance |
| BCAA | Branch chain amino acid |
| HOMA-IR | Homeostatic Model Assessment of Insulin Resistance |
| EAA | Essential amino acids |
| NEAA | Non-essential amino acids |
| BMI | Body mass index |
| RMR | Resting metabolic rate |
| PAL | Physical activity levels |
| IPAQ | International Physical Activity Questionnaire |
| BP | Blood pressure |
References
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Asghari, G.; Farhadnejad, H.; Teymoori, F.; Mirmiran, P.; Tohidi, M.; Azizi, F. High dietary intake of branched-chain amino acids is associated with an increased risk of insulin resistance in adults. J. Diabetes 2018, 10, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Geidenstam, N.; Al-Majdoub, M.; Ekman, M.; Spégel, P.; Ridderstråle, M. Metabolite profiling of obese individuals before and after a one year weight loss program. Int. J. Obes. 2017, 41, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.-S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405. [Google Scholar] [CrossRef]
- Newgard, C.B. Interplay between Lipids and Branched-Chain Amino Acids in Development of Insulin Resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef]
- Shou, J.; Chen, P.J.; Xiao, W.H. The effects of BCAAs on insulin resistance in athletes. J. Nutr. Sci. Vitaminol. 2019, 65, 383–389. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 723–736. [Google Scholar] [CrossRef]
- White, P.J.; Lapworth, A.L.; An, J.; Wang, L.; McGarrah, R.W.; Stevens, R.D.; Ilkayeva, O.; George, T.; Muehlbauer, M.J.; Bain, J.R.; et al. Branched-chain amino acid restriction in Zucker-fatty rats improves muscle insulin sensitivity by enhancing efficiency of fatty acid oxidation and acyl-glycine export. Mol. Metab. 2016, 5, 538–551. [Google Scholar] [CrossRef]
- Karusheva, Y.; Koessler, T.; Strassburger, K.; Markgraf, D.; Mastrototaro, L.; Jelenik, T.; Simon, M.C.; Pesta, D.; Zaharia, O.P.; Bódis, K.; et al. Short-term dietary reduction of branched-chain amino acids reduces meal-induced insulin secretion and modifies microbiome composition in type 2 diabetes: A randomized controlled crossover trial. Am. J. Clin. Nutr. 2019, 110, 1098–1107. [Google Scholar] [CrossRef]
- Thalacker-Mercer, A.E.; Fleet, J.C.; Craig, B.A.; Carnell, N.S.; Campbell, W.W. Inadequate protein intake affects skeletal muscle transcript profiles in older humans. Am. J. Clin. Nutr. 2007, 85, 1344–1352. [Google Scholar] [CrossRef]
- Frazier, D.M.; Allgeier, C.; Homer, C.; Marriage, B.J.; Ogata, B.; Rohr, F.; Splett, P.L.; Stembridge, A.; Singh, R.H. Nutrition management guideline for maple syrup urine disease: An evidence- and consensus-based approach. Mol. Genet. Metab. 2014, 112, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Henry, C. Basal metabolic rate studies in humans: Measurement and development of new equations. Public Health Nutr. 2005, 8, 1133–1152. [Google Scholar] [CrossRef] [PubMed]
- Type 2 Diabetes in Adults: Management NICE Guideline. 2015. Available online: www.nice.org.uk/guidance/ng28 (accessed on 30 December 2020).
- Schmidt, J.; Rinaldi, S.; Ferrari, P.; Carayol, M.; Achaintre, D.; Scalbert, A.; Cross, A.J.; Gunter, M.J.; Fensom, G.K.; Appleby, P.N.; et al. Metabolic profiles of male meat eaters, fish eaters, vegetarians, and vegans from the EPIC-Oxford cohort. Am. J. Clin. Nutr. 2015, 102, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.A.; Southgate, D.A.T. McCance and Widdowson’s The Composition of Foods; Springer: Berlin, Heidelberg, 1987. [Google Scholar]
- Mitchell, W.K.; Phillips, B.E.; Hill, I.; Greenhaff, P.L.; Lund, J.N.; Williams, J.P.; Rankin, D.; Wilkinson, D.J.; Smith, K.; Atherton, P.J. Human skeletal muscle is refractory to the anabolic effects of leucine during the postprandial muscle-full period in older men. Clin. Sci. 2017, 131, 2643–2653. [Google Scholar] [CrossRef] [PubMed]
- Gutch, M.; Kumar, S.; Razi, S.M.; Gupta, K.; Gupta, A. Assessment of insulin sensitivity/resistance. Indian J. Endocrinol. Metab. 2015, 19, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2019, 81, 139–164. [Google Scholar] [CrossRef]
- Cummings, N.E.; Williams, E.M.; Kasza, I.; Konon, E.N.; Schaid, M.D.; Schmidt, B.A.; Poudel, C.; Sherman, D.S.; Yu, D.; Apelo, S.I.A.; et al. Restoration of metabolic health by decreased consumption of branched-chain amino acids. J. Physiol. 2018, 596, 623–645. [Google Scholar] [CrossRef]
- Cavallaro, N.L.; Garry, J.; Shi, X.; Gerszten, R.E.; Anderson, E.J.; Walford, G.A. A pilot, short-term dietary manipulation of branched chain amino acids has modest influence on fasting levels of branched chain amino acids. Food Nutr. Res. 2016, 60. [Google Scholar] [CrossRef]
- Poortmans, J.R.; Carpentier, A. Protein metabolism and physical training: Any need for amino acid supplementation? Nutrire 2016, 41, 1–17. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Bukhari, S.S.; E Phillips, B.; Limb, M.C.; Cegielski, J.; Brook, M.S.; Rankin, D.; Mitchell, W.K.; Kobayashi, H.; Williams, J.P.; et al. Effects of leucine-enriched essential amino acid and whey protein bolus dosing upon skeletal muscle protein synthesis at rest and after exercise in older women. Clin. Nutr. 2018, 37, 2011–2021. [Google Scholar] [CrossRef]
- Blackburn, P.R.; Gass, J.M.; E Vairo, F.P.; Farnham, K.M.; Atwal, H.K.; Mantia, S.K.M.-; Klee, E.W.; Atwal, P.S. Maple syrup urine disease: Mechanisms and management. Appl. Clin. Genet. 2017, 10, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, Y.; Qi, Q.; Hruby, A.; E Manson, J.; Willett, W.C.; Wolpin, B.M.; Hu, F.B.; Qi, L. Cumulative consumption of branched-chain amino acids and incidence of type 2 diabetes. Int. J. Epidemiol. 2016, 45, 1482–1492. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, Y.; Liang, X.; Zou, L.; Ong, C.N.; Yuan, J.M. Serum amino acids in association with prevalent and incident type 2 diabetes in a Chinese population. Metabolites 2019, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Liu, B.; Qu, H.; Zhang, L.; Lu, Y.; Xu, Y.; Lyu, Z.; Zheng, H. A High Level of Circulating Valine Is a Biomarker for Type 2 Diabetes and Associated with the Hypoglycemic Effect of Sitagliptin. Mediat. Inflamm. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.H.; Crosslin, D.R.; Haynes, C.S.; Nelson, S.; Turer, C.B.; Stevens, R.D.; Muehlbauer, M.J.; Wenner, B.R.; Bain, J.R.; Laferrère, B.; et al. Branched-chain amino acid levels are associated with improvement in insulin resistance with weight loss. Diabetologia 2012, 55, 321–330. [Google Scholar] [CrossRef]
- Smith, G.I.; Yoshino, J.; Stromsdorfer, K.L.; Klein, S.J.; Magkos, F.; Reeds, M.N.; Klein, S.; Mittendorfer, B. Protein Ingestion Induces Muscle Insulin Resistance Independent of Leucine-Mediated mTOR Activation. Diabetes 2015, 64, 1555–1563. [Google Scholar] [CrossRef]
- Fontana, L.; Cummings, N.E.; Apelo, S.I.A.; Neuman, J.C.; Kasza, I.; Schmidt, B.A.; Cava, E.; Spelta, F.; Tosti, V.; Syed, F.A.; et al. Decreased Consumption of Branched-Chain Amino Acids Improves Metabolic Health. Cell Rep. 2016, 16, 520–530. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Diet | Control Diet | BCAA-Restricted Diet | ||
|---|---|---|---|---|
| Menu | Food | Supplement | Food | Supplement |
| Breakfast | Loprofin sliced loaf with peanut butter. | 34% of daily amount to be consumed. Each sachet contains Asn, Glu and Gly. | Loprofin sliced loaf with peanut butter. | 34% of daily amount to be consumed. Each sachet contains Asn, Glu, Gly, Iso, Leu and Val. |
| Snack 1 | Apple and cinnamon muffin made with Loprofin mix. | No supplement taken. | Apple and cinnamon muffin made with Loprofin mix. | No supplement taken. |
| Lunch | Carrot and coriander soup made with Snowpro with a cheese sandwich. | 21% of daily amount to be consumed. Each sachet contains Asn, Glu and Gly. | Carrot and coriander soup made with Snowpro with a cheese sandwich. | 21% of daily amount to be consumed. Each sachet contains Asn, Glu, Gly, Iso, Leu and Val. |
| Snack 2 | Apple and cinnamon muffin made with Loprofin mix. | No supplement taken. | Apple and cinnamon muffin made with Loprofin mix. | No supplement taken. |
| Dinner | Vegetable curry with either Loprofin rice (or pasta). | 26% of daily amount to be consumed. Each sachet contains Asn, Glu and Gly. | Vegetable curry with either Loprofin rice (or pasta). | 26% of daily amount to be consumed. Each sachet contains Asn, Glu, Gly, Iso, Leu and Val. |
| Snack 3 | Mixed chocolate and almond snack. | 19% of daily amount to be consumed. Each sachet contains Asn, Glu and Gly. | Mixed chocolate and almond snack. | 19% of daily amount to be consumed. Each sachet contains Asn, Glu, Gly, Iso, Leu and Val. |
| Control | BCAA-Restricted | |||
|---|---|---|---|---|
| Baseline | Day 7 | Baseline | Day 7 | |
| Age (Years) | 27 ± 7 | * | 24 ± 3 | * |
| Height (m) | 161.4 ± 12 | * | 173.4 ± 7 | * |
| Weight (kg) | 62.2 ± 5 | 62.2 ± 5 | 70.2 ± 9 | 70.2 ± 9 |
| BMI (Kg·m−2) | 22.8 ± 6 | 22.8 ± 6 | 22.2 ± 5 | 22.2 ± 5 |
| BP | 120/82 | 118/69 | 116/83 | 110/79 |
| Control | BCAA-Restricted | |||||
|---|---|---|---|---|---|---|
| Baseline | Day 7 | p Value | Baseline | Day 7 | p Value | |
| Glucose (mmol/L) | 5.5 ± 0.2 | 5.2 ± 0.3 | 0.41 | 5.3 ± 0.2 | 4.9 ± 0.3 | 0.07 |
| Insulin (miliU/L) | 3.8 ± 0.5 | 5.5 ± 0.9 | 0.26 | 6.3 ± 1.0 | 4.9 ± 0.6 | 0.17 |
| HOMA-IR | 1.0 ± 0.2 | 1.3 ± 0.1 | 0.24 | 1.5 ± 0.2 | 1.0 ± 0.1 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramzan, I.; Taylor, M.; Phillips, B.; Wilkinson, D.; Smith, K.; Hession, K.; Idris, I.; Atherton, P. A Novel Dietary Intervention Reduces Circulatory Branched-Chain Amino Acids by 50%: A Pilot Study of Relevance for Obesity and Diabetes. Nutrients 2021, 13, 95. https://doi.org/10.3390/nu13010095
Ramzan I, Taylor M, Phillips B, Wilkinson D, Smith K, Hession K, Idris I, Atherton P. A Novel Dietary Intervention Reduces Circulatory Branched-Chain Amino Acids by 50%: A Pilot Study of Relevance for Obesity and Diabetes. Nutrients. 2021; 13(1):95. https://doi.org/10.3390/nu13010095
Chicago/Turabian StyleRamzan, Imran, Moira Taylor, Beth Phillips, Daniel Wilkinson, Kenneth Smith, Kate Hession, Iskandar Idris, and Philip Atherton. 2021. "A Novel Dietary Intervention Reduces Circulatory Branched-Chain Amino Acids by 50%: A Pilot Study of Relevance for Obesity and Diabetes" Nutrients 13, no. 1: 95. https://doi.org/10.3390/nu13010095
APA StyleRamzan, I., Taylor, M., Phillips, B., Wilkinson, D., Smith, K., Hession, K., Idris, I., & Atherton, P. (2021). A Novel Dietary Intervention Reduces Circulatory Branched-Chain Amino Acids by 50%: A Pilot Study of Relevance for Obesity and Diabetes. Nutrients, 13(1), 95. https://doi.org/10.3390/nu13010095