
Dysregulated Arginine Metabolism in Young Patients with Chronic Persistent Asthma and in Human Bronchial Epithelial Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Participants and Laboratory Tests
2.2. Serum Sample Preparation and Metabolism Profile Determination
2.3. Cell Culture
2.4. NOx (Nitrites + Nitrates) Determination
2.5. Statistical Analysis
3. Results
3.1. Clinical Characteristics of the Subjects Enrolled
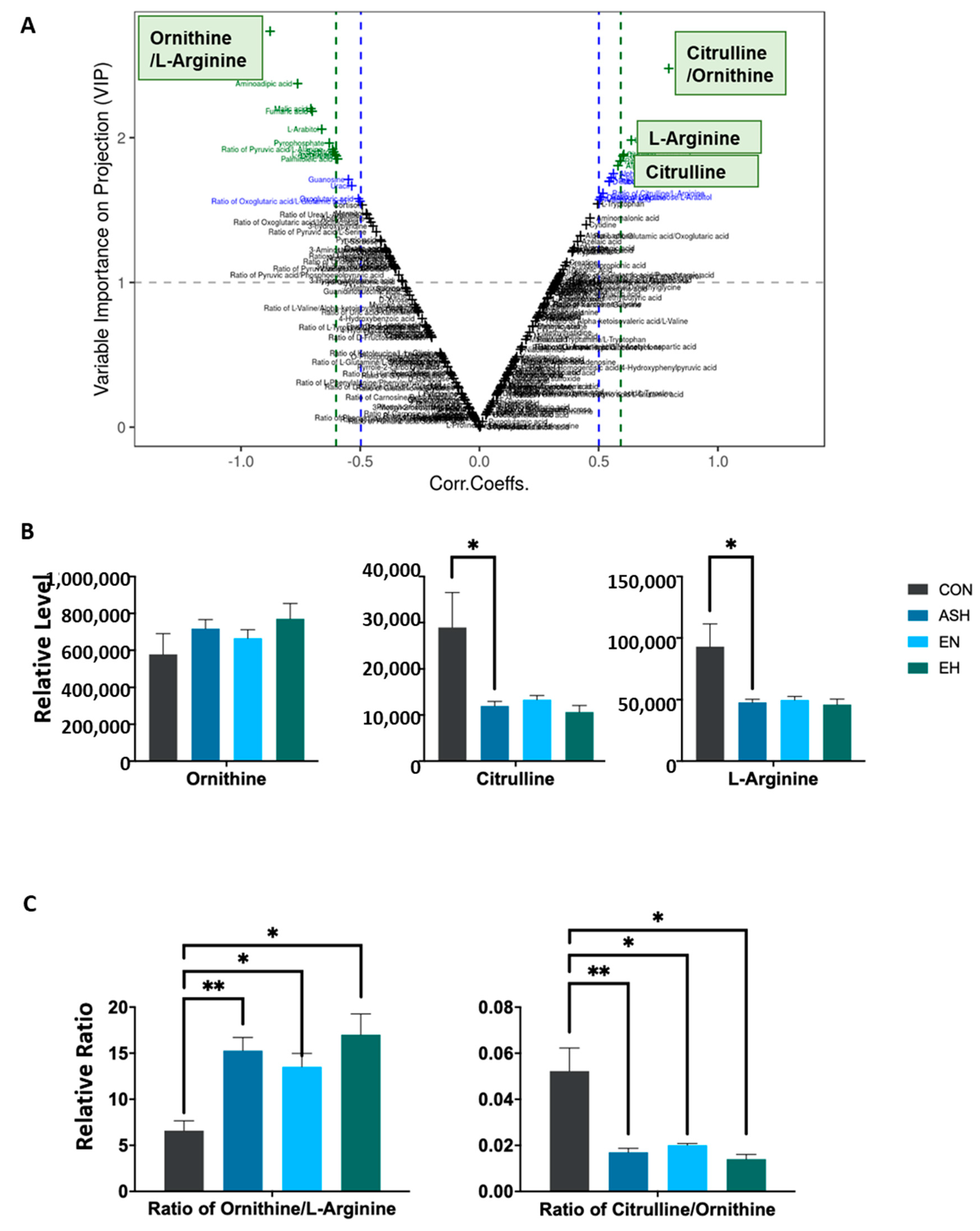
3.2. Comprehensive Metabolomics Profile and Composition in Asthma Patients and Control
3.3. Significantly Altered Pathways during Chronic Persistent Asthma
3.4. Significantly Dysregulated Amino Acids and Their Ratios in Asthma and Control Cohort
3.5. Coefficients between Significantly Altered Metabolites and Serum Eosinophilia Counts
3.6. Addition of L-Citrulline Prevented Asymmetric Dimethylarginine (ADMA)-Mediated Reduction of NO Metabolites in HBECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Approval and Consent to Participate
Abbreviations
| ACT | Asthma Control Test |
| ADMA | Asymmetric Dimethylarginine |
| AHR | Airway Hyperresponsiveness |
| BALF | Bronchoalveolar Lavage Fluid |
| BMI | Body Mass Index |
| HBECs | Human Bronchial Epithelial Cells |
| NO | Nitric Oxide |
References
- Akbari, O.; Stock, P.; Meyer, E.; Kronenberg, M.; Sidobre, S.; Nakayama, T.; Taniguchi, M.; Grusby, M.J.; DeKruyff, R.H.; Umetsu, D.T. Essential role of NKT cells producing IL-4 and IL-13 in the development of allergen-induced airway hyperreactivity. Nat. Med. 2003, 9, 582–588. [Google Scholar] [CrossRef]
- Finkelman, F.D.; Boyce, J.A.; Vercelli, D.; Rothenberg, M.E. Key advances in mechanisms of asthma, allergy, and immunology in 2009. J. Allergy Clin. Immunol. 2010, 125, 312–318. [Google Scholar] [CrossRef]
- Kraj, L.; Krawiec, M.; Koter, M.; Grabon, W.; Kraj, G.; Cholojczyk, M.; Kulus, M.; Baranczyk-Kuzma, A. Altered L-arginine metabolism in children with controlled asthma. Allergy Asthma Proc. 2014, 35, 80–83. [Google Scholar] [CrossRef]
- Quan-Jun, Y.; Jian-Ping, Z.; Jian-Hua, Z.; Yong-Long, H.; Bo, X.; Jing-Xian, Z.; Bona, D.; Yuan, Z.; Cheng, G. Distinct Metabolic Profile of Inhaled Budesonide and Salbutamol in Asthmatic Children during Acute Exacerbation. Basic Clin. Pharmacol. Toxicol. 2017, 120, 303–311. [Google Scholar] [CrossRef]
- Scott, J.A.; Grasemann, H. Arginine metabolism in asthma. Immunol. Allergy Clin. N. Am. 2014, 34, 767–775. [Google Scholar] [CrossRef]
- Zhang, P.; Guo, X.; Xiao, Y.; Zhang, Q.; Ding, C. Twisted intramolecular charge transfer (TICT) based fluorescent probe for lighting up serum albumin with high sensitivity in physiological conditions. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 223, 117318. [Google Scholar] [CrossRef]
- She, J.; Guo, M.; Li, H.; Liu, J.; Liang, X.; Liu, P.; Zhou, B.; Liu, S.; Deng, Y.; Lou, B.; et al. Targeting amino acids metabolic profile to identify novel metabolic characteristics in atrial fibrillation. Clin. Sci. 2018, 132, 2135–2146. [Google Scholar] [CrossRef] [Green Version]
- She, J.; Feng, J.; Deng, Y.; Sun, L.; Wu, Y.; Guo, M.; Liang, X.; Li, J.; Xia, Y.; Yuan, Z. Correlation of Triiodothyronine Level with In-Hospital Cardiac Function and Long-Term Prognosis in Patients with Acute Myocardial Infarction. Dis. Markers 2018, 2018, 5236267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- She, J.; Deng, Y.; Wu, Y.; Xia, Y.; Li, H.; Liang, X.; Shi, R.; Yuan, Z. Hemoglobin A1c is associated with severity of coronary artery stenosis but not with long term clinical outcomes in diabetic and nondiabetic patients with acute myocardial infarction undergoing primary angioplasty. Cardiovasc. Diabetol. 2017, 16, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Byrne, P.M.; Reddel, H.K.; Eriksson, G.; Ostlund, O.; Peterson, S.; Sears, M.R.; Jenkins, C.; Humbert, M.; Buhl, R.; Harrison, T.W.; et al. Measuring asthma control: A comparison of three classification systems. Eur. Respir. J. 2010, 36, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Cai, G.; Su, M.; Chen, T.; Zheng, X.; Xu, Y.; Ni, Y.; Zhao, A.; Xu, L.X.; Cai, S.; et al. Serum metabolite profiling of human colorectal cancer using GC-TOFMS and UPLC-QTOFMS. J. Proteome Res. 2009, 8, 4844–4850. [Google Scholar] [CrossRef]
- Wang, J.H.; Chen, W.L.; Li, J.M.; Wu, S.F.; Chen, T.L.; Zhu, Y.M.; Zhang, W.N.; Li, Y.; Qiu, Y.P.; Zhao, A.H.; et al. Prognostic significance of 2-hydroxyglutarate levels in acute myeloid leukemia in China. Proc. Natl. Acad. Sci. USA 2013, 110, 17017–17022. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Qiu, Y.; Jiang, W.; Suttlemyre, K.; Su, M.; Zhang, W.; Jia, W.; Du, X. ADAP-GC 2.0: Deconvolution of coeluting metabolites from GC/TOF-MS data for metabolomics studies. Anal. Chem. 2012, 84, 6619–6629. [Google Scholar] [CrossRef]
- Frolkis, A.; Knox, C.; Lim, E.; Jewison, T.; Law, V.; Hau, D.D.; Liu, P.; Gautam, B.; Ly, S.; Guo, A.C.; et al. SMPDB: The Small Molecule Pathway Database. Nucleic Acids Res. 2010, 38, D480–D487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewison, T.; Su, Y.; Disfany, F.M.; Liang, Y.; Knox, C.; Maciejewski, A.; Poelzer, J.; Huynh, J.; Zhou, Y.; Arndt, D.; et al. SMPDB 2.0: Big improvements to the Small Molecule Pathway Database. Nucleic Acids Res. 2014, 42, D478–D484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnica, D.; Que, L.G.; Baffi, C.; Grasemann, H.; Fiedler, K.; Yang, Z.; Etling, E.; Wasil, K.; Wenzel, S.E.; Freeman, B.; et al. l-citrulline prevents asymmetric dimethylarginine-mediated reductions in nitric oxide and nitrosative stress in primary human airway epithelial cells. Clin. Exp. Allergy 2017, 47, 190–199. [Google Scholar] [CrossRef]
- Holguin, F.; Grasemann, H.; Sharma, S.; Winnica, D.; Wasil, K.; Smith, V.; Cruse, M.H.; Perez, N.; Coleman, E.; Scialla, T.J.; et al. L-Citrulline increases nitric oxide and improves control in obese asthmatics. JCI Insight 2019, 4, e131733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, M.F.; DiNicolantonio, J.J.; Lerner, A. Review—Nutraceuticals Can Target Asthmatic Bronchoconstriction: NADPH Oxidase-Dependent Oxidative Stress, RhoA and Calcium Dynamics. J. Asthma Allergy 2021, 14, 685–701. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.A.; Maarsingh, H.; Holguin, F.; Grasemann, H. Arginine Therapy for Lung Diseases. Front. Pharmacol. 2021, 12, 627503. [Google Scholar] [CrossRef]
- Mabalirajan, U.; Ahmad, T.; Leishangthem, G.D.; Joseph, D.A.; Dinda, A.K.; Agrawal, A.; Ghosh, B. Beneficial effects of high dose of L-arginine on airway hyperresponsiveness and airway inflammation in a murine model of asthma. J. Allergy Clin. Immunol. 2010, 125, 626–635. [Google Scholar] [CrossRef]
- North, M.L.; Khanna, N.; Marsden, P.A.; Grasemann, H.; Scott, J.A. Functionally important role for arginase 1 in the airway hyperresponsiveness of asthma. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 296, L911–L920. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.A.; North, M.L.; Rafii, M.; Huang, H.; Pencharz, P.; Subbarao, P.; Belik, J.; Grasemann, H. Asymmetric dimethylarginine is increased in asthma. Am. J. Respir. Crit. Care Med. 2011, 184, 779–785. [Google Scholar] [CrossRef] [PubMed]
- North, M.L.; Grasemann, H.; Khanna, N.; Inman, M.D.; Gauvreau, G.M.; Scott, J.A. Increased ornithine-derived polyamines cause airway hyperresponsiveness in a mouse model of asthma. Am. J. Respir. Cell Mol. Biol. 2013, 48, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Lane, C.; Knight, D.; Burgess, S.; Franklin, P.; Horak, F.; Legg, J.; Moeller, A.; Stick, S. Epithelial inducible nitric oxide synthase activity is the major determinant of nitric oxide concentration in exhaled breath. Thorax 2004, 59, 757–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ckless, K.; van der Vliet, A.; Janssen-Heininger, Y. Oxidative-nitrosative stress and post-translational protein modifications: Implications to lung structure-function relations. Arginase modulates NF-kappaB activity via a nitric oxide-dependent mechanism. Am. J. Respir. Cell Mol. Biol. 2007, 36, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Asthma (n = 30) | Control (n = 30) | p |
|---|---|---|---|
| Age (years) | 25.5 ± 2.3 | 25.0 ± 2.8 | ns |
| Female (%) | 50% | 50% | ns |
| BMI (kg/m2) | 21.4 ± 2.7 | 20.8 ± 2.3 | ns |
| Eosinophil count (*109/L) | 0.35 ± 0.12 | 0.16 ± 0.08 | <0.01 |
| ACT (points) | 22.5 ± 2.1 | - | - |
| Class | Name | Corr. | p |
|---|---|---|---|
| Amino Acid | Ratio of Citrulline/Ornithine | −0.84 | 0.04 |
| Carbohydrates | L-Arabitol | 0.84 | 0.04 |
| Fatty Acids | Palmitoleic acid | −0.80 | 0.06 |
| Amino Acid | Ratio of Ornithine/L-arginine | 0.71 | 0.11 |
| Amino Acid | Citrulline | −0.66 | 0.15 |
| Carbohydrates | Sorbitol | 0.65 | 0.16 |
| Amino Acid | L-arginine | −0.63 | 0.18 |
| Carbohydrates | L-Arabinose | 0.52 | 0.29 |
| Organic Acids | Malic acid | −0.58 | 0.30 |
| Fatty Acids | Tetracosanoic acid | −0.48 | 0.33 |
| Carbohydrates | Ribonolactone | 0.44 | 0.38 |
| Carbohydrates | D-Threitol | 0.39 | 0.44 |
| Organic Acids | Pyrophosphate | −0.35 | 0.49 |
| Amino Acid | Aminoadipic acid | −0.33 | 0.53 |
| Amino Acid | Ratio of Pyruvic acid/L-Alanine | −0.16 | 0.76 |
| Amino Acid | Alpha-ketoisovaleric acid | 0.08 | 0.88 |
| Organic Acids | Fumaric acid | 0.03 | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, B.; Jiang, G.T.; Liu, H.; Guo, M.; Liu, J.; She, J. Dysregulated Arginine Metabolism in Young Patients with Chronic Persistent Asthma and in Human Bronchial Epithelial Cells. Nutrients 2021, 13, 4116. https://doi.org/10.3390/nu13114116
Zhou B, Jiang GT, Liu H, Guo M, Liu J, She J. Dysregulated Arginine Metabolism in Young Patients with Chronic Persistent Asthma and in Human Bronchial Epithelial Cells. Nutrients. 2021; 13(11):4116. https://doi.org/10.3390/nu13114116
Chicago/Turabian StyleZhou, Bo, Gulinigaer Tuerhong Jiang, Hui Liu, Manyun Guo, Junhui Liu, and Jianqing She. 2021. "Dysregulated Arginine Metabolism in Young Patients with Chronic Persistent Asthma and in Human Bronchial Epithelial Cells" Nutrients 13, no. 11: 4116. https://doi.org/10.3390/nu13114116