
Dietary Supplementation with Biobran/MGN-3 Increases Innate Resistance and Reduces the Incidence of Influenza-like Illnesses in Elderly Subjects: A Randomized, Double-Blind, Placebo-Controlled Pilot Clinical Trial



Abstract
:1. Background
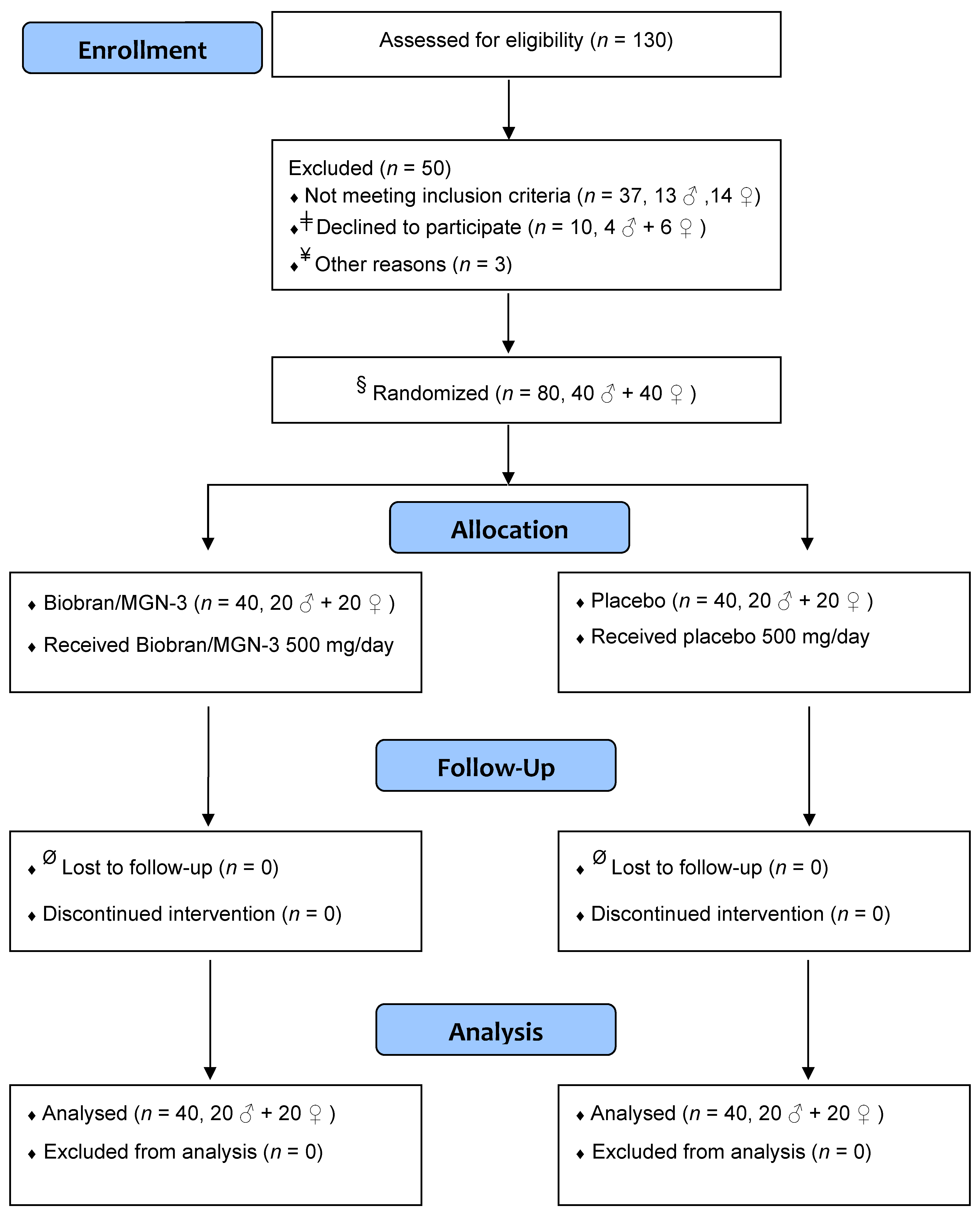
2. Subjects and Methods
2.1. Trial Design
2.2. Participants
2.3. Inclusion and Exclusion Criteria
2.4. Consent
2.5. Randomization and Allocation Concealment
2.6. Intervention
2.7. Ingredients of Biobran/MGN-3 Sachets
2.8. Blinding
2.9. Follow-Up
2.10. Outcome
2.11. Sample Size
2.12. Laboratory Investigations
2.13. Degranulation Assay for NK Cell Activity
2.14. Cell Culture and Flow Cytometry
2.15. Statistical Analysis
3. Results
3.1. Biobran/MGN-3 Did Not Adversely Affect the Hematological, Liver, and Kidney Functions
3.2. Biobran/MGN-3 Supplementation Significantly Reduced the ILI Incidence Rate and Density
3.3. Biobran/MGN-3 Specifically Enhanced NK Cell Activity but Not NKT Activity
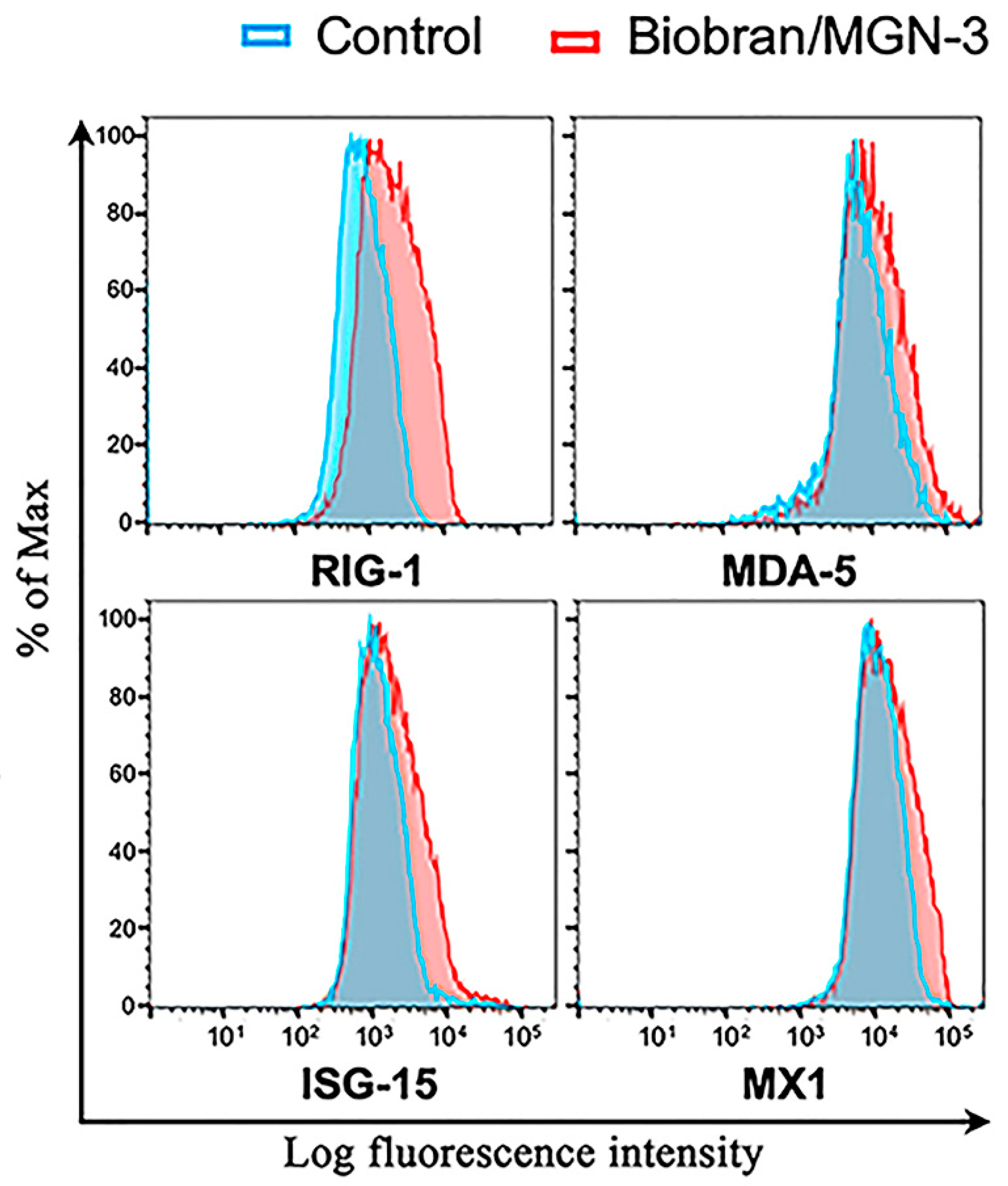
3.4. Biobran/MGN-3 Upregulates the Expression of RIG-1, MDA5, ISG15, and MX1 in Pulmonary Epithelial BEAS-2B Cells
4. Discussion
5. Conclusions and Future Direction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stohr, K. Influenza—WHO cares. Lancet Infect. Dis. 2002, 2, 517. [Google Scholar] [CrossRef]
- Lee, N.; Chan, P.K.; Lui, G.C.; Wong, B.C.; Sin, W.W.; Choi, K.W.; Wong, R.Y.; Lee, E.L.; Yeung, A.C.; Ngai, K.L.; et al. Complications and outcomes of pandemic 2009 Influenza A (H1N1) virus infection in hospitalized adults: How do they differ from those in seasonal influenza? J. Infect. Dis. 2011, 203, 1739–1747. [Google Scholar] [CrossRef]
- Fitzner, J.; Qasmieh, S.; Mounts, A.W.; Alexander, B.; Besselaar, T.; Briand, S.; Brown, C.; Clark, S.; Dueger, E.; Gross, D.; et al. Revision of clinical case definitions: Influenza-like illness and severe acute respiratory infection. Bull. World Health Organ. 2018, 96, 122–128. [Google Scholar] [CrossRef]
- Kammerer, P.E.; Montiel, S.; Kriner, P.; Bojorquez, I.; Ramirez, V.B.; Vazquez-Erlbeck, M. Influenza-like illness surveillance on the California–Mexico border, 2004–2009. Influenza Other Respir. Viruses 2012, 6, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Bollaerts, K.; Antoine, J.; Van Casteren, V.; Ducoffre, G.; Hens, N.; Quoilin, S. Contribution of respiratory pathogens to influenza-like illness consultations. Epidemiol. Infect. 2013, 141, 2196–2204. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Pan, L.; Sun, Q.; Zhu, W.; Zhu, L.; Ye, C.; Xue, C.; Wang, Y.; Liu, Q.; Ma, P.; et al. The clinical and etiological characteristics of influenza-like illness (ILI) in outpatients in Shanghai, China, 2011 to 2013. PLoS ONE 2015, 10, e0119513. [Google Scholar] [CrossRef] [PubMed]
- Souty, C.; Masse, S.; Valette, M.; Behillil, S.; Bonmarin, I.; Pino, C. Baseline characteristics and clinical symptoms related to respiratory viruses identified among patients presenting with influenza-like illness in primary care. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2019, 25, 1147–1153. [Google Scholar] [CrossRef] [Green Version]
- Refaey, S.; Amin, M.; Labib, M.; Kandeel, A. Influenza virus positivity and circulating subtypes among cases of influenza-like illness and severe acute respiratory infection, Egypt, 2012–2015. East Mediterr. Health J. 2016, 22, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Pop-Vicas, A.; Gravenstein, S. Influenza in the elderly: A mini-review. Gerontology 2011, 57, 397–404. [Google Scholar] [CrossRef]
- Talbot, H.K. Influenza in older adults. Infect. Dis. Clin. N. Am. 2017, 31, 757–766. [Google Scholar] [CrossRef]
- Takahasi, K.; Yoneyama, M.; Nishihori, T.; Hirai, R.; Kumeta, H.; Narita, R.; Gale, M., Jr.; Inagaki, F.; Fujita, T. Nonself RNA-sensing mechanism of RIG-I helicase and activation of antiviral immune responses. Mol. Cell. 2008, 29, 428–440. [Google Scholar] [CrossRef]
- Baum, A.; Sachidanandam, R.; Garcia-Sastre, A. Preference of RIG-I for short viral RNA molecules in infected cells revealed by next-generation sequencing. Proc. Natl. Acad. Sci. USA 2010, 108, 3092. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barral, P.M.; Sarkar, D.; Su, Z.Z.; Barber, G.N.; DeSalle, R.; Racaniello, V.R.; Fisher, P.B. Functions of the cytoplasmic RNA sensors RIG-I and MDA-5: Key regulators of innate immunity. Pharmacol. Ther. 2009, 124, 219–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, A.; Medzhitov, R. Regulation of adaptive immunity by the innate immune system. Science 2010, 327, 291–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef]
- Lindenmann, J.; Lane, C.A.; Hobson, D. The resistance of A2G mice to myxoviruses. J. Immunol. 1963, 90, 942–951. [Google Scholar]
- Lenschow, D.J.; Lai, C.; Frias-Staheli, N.; Giannakopoulos, N.V.; Lutz, A.; Wolff, T. IFN-stimulated gene 15 functions as a critical antiviral molecule against influenza, herpes, and Sindbis viruses. Proc. Natl. Acad. Sci. USA 2007, 104, 1371–1376. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.; Struckhoff, J.J.; Schneider, J.; Martinez-Sobrido, L.; Wolff, T.; Garcia-Sastre, A.; Zhang, D.E.; Lenschow, D.J. Mice lacking the ISG15 E1 enzyme UbE1L demonstrate increased susceptibility to both mouse-adapted and non-mouse-adapted influenza B virus infection. J. Virol. 2009, 83, 1147–1151. [Google Scholar] [CrossRef] [Green Version]
- Orange, J.S. Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect. 2002, 4, 1545–1558. [Google Scholar] [CrossRef]
- Lodoen, M.B.; Lanier, L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006, 18, 391–398. [Google Scholar] [CrossRef]
- Vivier, E.; Ugolini, S.; Blaise, D.; Chabannon, C.; Brossay, L. Targeting natural killer cells and natural killer T cells in cancer. Nat. Rev. Immunol. 2012, 12, 239–252. [Google Scholar] [CrossRef]
- Terunuma, H.; Deng, X.; Dewan, Z.; Fujimoto, S.; Yamamoto, N. Potential role of NK cells in the induction of immune responses: Implications for NK cell-based immunotherapy for cancers and viral infections. Int. Rev. Immunol. 2008, 27, 93–110. [Google Scholar] [CrossRef]
- Cooley, S.; Burns, L.J.; Repka, T.; Miller, J.S. Natural killer cell cytotoxicity of breast cancer targets is enhanced by two distinct mechanisms of antibody-dependent cellular cytotoxicity against LFA-3 and HER2/neu. Exp. Hematol. 1999, 27, 1533–1541. [Google Scholar] [CrossRef]
- Hammer, Q.; Romagnani, C. About training and memory: NK-cell adaptation to viral infections. Adv. Immunol. 2017, 133, 171–207. [Google Scholar]
- Peng, H.; Tian, Z. NK cells in liver homeostasis and viral hepatitis. Sci. China Life Sci. 2018, 61, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Mikulak, J.; Oriolo, F.; Zaghi, E.; Di Vito, C.; Mavilio, D. Natural killer cells in HIV-1 infection and therapy. AIDS 2017, 31, 2317–2330. [Google Scholar] [CrossRef]
- Flórez-Álvarez, L.; Hernandez, J.C.; Zapata, W. NK cells in HIV-1 infection: From basic science to vaccine strategies. Front. Immunol. 2018, 9, 2290. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.K.; Kim, T.S.; Hufford, M.M.; Braciale, T.J. Viral infection of the lung: Host response and sequelae. J. Allergy Clin. Immunol. 2013, 132, 1263–1276. [Google Scholar] [CrossRef] [PubMed]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.; Yagita, H.; Takeda, K.; van Dommelen, S.L.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK cell cytotoxicity. Mol. Immunol. 2005, 42, 501–510. [Google Scholar] [CrossRef]
- Miller, J.S. Biology of Natural Killer Cells in Cancer and Infection. Cancer Investig. 2002, 20, 405–419. [Google Scholar] [CrossRef]
- Ghoneum, M.; Suzuki, K.; Gollapudi, S. Phorbol myristate acetate corrects impaired NK function of old mice. Scand. J. Immunol. 1991, 34, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Mariani, E.; Sgobbi, S.; Meneghetti, A.; Tadolini, M.; Tarozzi, A.; Sinoppi, M.; Cattini, L.; Facchini, A. Perforins in human cytolytic cells: The effect of age. Mech. Ageing Dev. 1996, 92, 195–209. [Google Scholar] [CrossRef]
- Rukavina, D.; Laskarin, G.; Rubesa, G.; Strbo, N.; Bedenicki, I.; Manestar, D.; Glavas, M.; Christmas, S.E.; Podack, E.R. Age-related decline of perforin expression in human cytotoxic T lymphocytes and natural killer cells. Blood 1998, 92, 2410–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoneum, M. Enhancement of human natural killer cell activity by modified arabinoxylan from rice bran (MGN-3). Int. J. Immunother. 1998, XIV, 89–99. [Google Scholar]
- Ghoneum, M.; Jewett, A. Production of tumor necrosis factor-alpha and interferon-gamma from human peripheral blood lymphocytes by MGN-3, a modified arabinoxylan from rice bran, and its synergy with interleukin-2 in vitro. Cancer Detect. Prev. 2000, 24, 314–324. [Google Scholar]
- Cholujova, D.; Jakubikova, J.; Czako, B.; Martisova, M.; Hunakova, L.; Duraj, J.; Mistrik, M.; Sedlak, J. MGN-3 arabinoxylan rice bran modulates innate immunity in multiple myeloma patients. Cancer Immunol. Immunother. 2013, 62, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martínez, A.; Valentín, J.; Fernández, L.; Hernández-Jiménez, E.; López-Collazo, E. Arabinoxylan rice bran (MGN-3/Biobran) enhances natural killer cell-mediated cytotoxicity against neuroblastoma in vitro and in vivo. Cytotherapy 2015, 17, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; Abedi, S. Enhancement of natural killer cell activity of aged mice by modified arabinoxylan rice bran (MGN-3/Biobran). J. Pharm. Pharmacol. 2004, 56, 1581–1588. [Google Scholar] [CrossRef]
- Elsaid, A.F.; Shaheen, M.; Ghoneum, M. Biobran/MGN-3, an arabinoxylan rice bran, enhances NK cell activity in geriatric subjects: A randomized, double blind, placebo-controlled clinical trial. Exp. Ther. Med. 2018, 15, 2313–2320. [Google Scholar] [CrossRef] [Green Version]
- Ghoneum, M.; Matsuura, M.; Gollapudi, S. Modified arabinoxylan rice bran (MGN3/Biobran) enhances intracellular killing of microbes by human phagocytic cells in vitro. Int. J. Immunopathol. Pharmacol. 2008, 21, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoneum, M.; Agrawal, S. Activation of human monocyte-derived dendritic cells in vitro by biological response modifier arabinoxylan rice bran (MGN-3/Biobran). Int. J. Immunopathol. Pharmcol. 2011, 24, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; Agrawal, S. MGN-3/Biobran enhances generation of cytotoxic CD8+ T cells via upregulation of DEC-205 expression on dendritic cells. Int. J. Immunopathol. Pharmacol. 2014, 27, 523–530. [Google Scholar] [CrossRef]
- Ghoneum, M. Anti-HIV activity in vitro of MGN-3, an activated arabinoxylane from rice bran. Biochem. Biophys. Res. Commun. 1998, 243, 25–29. [Google Scholar] [CrossRef]
- Tazawa, K.; Ichihashi, K.; Fujii, T.; Omura, K.; Anazawa, M.; Maeda, H. The orally administration of the Hydrolysis Rice Bran prevents a common cold syndrome for the elderly people based on immunomodulatory function. J. Trad. Med. 2003, 20, 132–141. [Google Scholar]
- Salama, H.; Medhat, E.; Shaheen, M.; Zekri, A.N.; Darwish, T.; Ghoneum, M. Arabinoxylan rice bran (Biobran) suppresses the viremia level in patients with chronic HCV infection: A randomized trial. Int. J. Immunopathol. Pharmacol. 2016, 29, 647–653. [Google Scholar] [CrossRef]
- Elsaid, A.F.; Fahmi, R.M.; Shaheen, M.; Ghoneum, M. The enhancing effects of Biobran/MGN-3, an arabinoxylan rice bran, on healthy old adults’ health-related quality of life: A randomized, double-blind, placebo-controlled clinical trial. Qual. Life Res. 2020, 29, 357–367. [Google Scholar] [CrossRef]
- Ghoneum, M.; Brown, J. NK immunorestoration of cancer patients by MGN-3, a modified arabinoxylan rice bran (study of 32 patients followed for up to 4 years). Anti-Aging Med. Ther. 1999, 3, 217–226. [Google Scholar]
- Takahara, K.; Sano, K. The life prolongation and QOL improvement effect of rice bran arabinoxylan derivative (MGN-3. Bio-Bran) for progressive cancer. Clin. Pharmacol. Ther. 2004, 14, 267–271. [Google Scholar]
- World Health Organization. Proposed Working Definition of an Older Person in Africa for the MDS Project. Available online: http://www.who.int/healthinfo/survey/ageingdefnolder/en/index.html (accessed on 6 May 2020).
- Centers for Disease Control and Prevention. Principles of Epidemiology in Public Health Practice, Third Edition. An Introduction to Applied Epidemiology and Biostatistics. Lesson 3: Measures of Risk; Section 2: Morbidity Frequency Measures. Available online: https://www.cdc.gov/csels/dsepd/ss1978/lesson3/section2.html (accessed on 3 September 2020).
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef]
- Dean, A.G.; Sullivan, K.M.; Soe, M.M. OpenEpi: Open Source Epidemiologic Statistics for Public Health, Version 3.01. Available online: www.OpenEpi.com (accessed on 29 January 2014).
- Corp IBM. IBM SPSS Statistics for Windows, Version 20; IBM Corp.: Armonk, NY, USA, 2010. [Google Scholar]
- Tazawa, K. BioBran/MGN-3 (Rice Bran Arabinoxylan Deritive): Basic and Clinical Application to Integrative Medicine; Iyakushuppan Co., Publishers: Tokyo, Japan, 2006; pp. 18–22. [Google Scholar]
- Daiwa Pharmaceutical Co., Ltd. BioBran. Available online: https://www.daiwa-pharm.com/english/product/biobran.html (accessed on 10 November 2020).
- Laue, C.; Stevens, Y.; van Erp, M.; Papazova, E.; Soeth, E.; Pannenbeckers, A.; Stolte, E.; Böhm, R.; Gall, S.L.; Falourd, X.; et al. Adjuvant Effect of Orally Applied Preparations Containing Non-Digestible Polysaccharides on Influenza Vaccination in Healthy Seniors: A Double-Blind, Randomised, Controlled Pilot Trial. Nutrients 2021, 13, 2683. [Google Scholar] [CrossRef]
- Boudreau, J.E.; Hsu, K.C. Natural killer cell education in human health and disease. Curr. Opin. Immunol. 2018, 50, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, H.; Mao, H.; Yu, M.; Yang, F.; Feng, T.; Fan, Y.; Lu, Q.; Shen, C.; Yin, Z.; et al. Impaired NK cell antiviral cytokine response against influenza virus in small-for-gestational-age neonates. Cell. Mol. Immunol. 2013, 10, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Sun, H.Y.; Xiao, W.H.; Zhang, C.; Tian, Z.G. Natural killer cell dysfunction in hepatocellular carcinoma and NK cell-based immunotherapy. Acta Pharmacol. Sin. 2015, 36, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Orange, J.S. Natural killer cell deficiency. J. Allergy Clin. Immunol. 2013, 132, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Loo, Y.M.; Gale, M., Jr. Immune signaling by RIG-I-like receptors. Immunity 2011, 34, 680–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gitlin, L.; Barchet, W.; Gilfillan, S.; Cella, M.; Beutler, B.; Flavell, R.A.; Diamond, M.S.; Colonna, M. Essential role of mda-5 in type I IFN responses to polyriboinosinic:polyribocytidylic acid and encephalomyocarditis picornavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 8459–8464. [Google Scholar] [CrossRef] [Green Version]
- van Kasteren, P.B.; Beugeling, C.; Ninaber, D.K.; Frias-Staheli, N.; van Boheemen, S.; Garcia-Sastre, A.; Snijder, E.J.; Kikkert, M. Arterivirus and nairovirus ovarian tumor domaincontaining deubiquitinases target activated RIG-I to control innate immune signaling. J. Virol. 2012, 86, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ye, X.; Dunker, W.; Song, Y.; Karijolich, J. RIG-I like receptor sensing of host RNAs facilitates the cell-intrinsic immune response to KSHV infection. Nat. Commun. 2018, 9, 4841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Filzmayer, C.; Ni, Y.; Sültmann, H.; Mutz, P.; Hiet, M.-S.; Vondran, F.W.; Bartenschlager, R.; Urban, S. Hepatitis D virus replication is sensed by MDA5 and induces IFN-b/l responses in hepatocytes. J. Hepatol. 2018, 69, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.J.; Lenschow, D.J. The antiviral activities of ISG15. J. Mol. Biol. 2013, 425, 4995–5008. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Sato, S.; Sotoyama, Y.; Orba, Y.; Sawa, H.; Yamauchi, H.; Sasaki, M.; Takaoka, A. RIG-I triggers a signaling-abortive anti-SARS-CoV-2 defense in human lung cells. Nat. Immunol. 2021, 22, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, N.G.; Chauveau, L.; Hertzog, J.; Bridgeman, A.; Fowler, G.; Moonen, J.P.; Dupont, M.; Russell, R.A.; Noerenberg, M.; Rehwinkel, J. The RNA sensor MDA5 detects SARS-CoV-2 infection. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Treatment | Parameter | Baseline | Post-Treatment | p-Value § |
|---|---|---|---|---|
| Placebo ǂ | RBCs (×106/µL) | 4.3 (0.5) | 4.1 (0.5) | 0.58 |
| HB (g/dL) | 11.3 (1.5) | 13.8 (1.4) | 0.40 | |
| HCT % | 35.7 (4.0) | 36.7 (3.5) | 0.51 | |
| MCHC (g/dL) | 27.3 (2.7) | 37.6 (0.7) | 0.80 | |
| WBC (×103/µL) | 5.7 (0.9) | 6.1 (1.2) | 0.76 | |
| Neutrophils % | 56.1 (14.5) | 57.3 (11.6) | 0.32 | |
| Lymphocytes % | 33.3 (10.2) | 39.5 (9.8) | 0.23 | |
| Platelets (×103/µL) | 218.8 (48.4) | 231.5 (48.0) | 0.19 | |
| ALP (U/L) | 185.5 (51.3) | 183.2 (50.1) | 0.66 | |
| ALT (U/L) | 22.0 (9.3) | 20.3 (8.4) | 0.15 | |
| AST (U/L) | 17.7 (4.6) | 18.3 (5.8) | 0.07 | |
| UA (mg/dL) | 7.4 (2.9) | 8.2 (3.8) | 0.51 | |
| Biobran/MGN-3 ǂ (500 mg/day) for 3 months | RBCs (×106/µL) | 4.8 (0.5) | 4.7 (0.6) | 0.18 |
| HB (g/dL) | 12.1 (1.7) | 13.9 (1.6) | 0.93 | |
| HCT % | 37.4 (4.4) | 36.9 (3.1) | 0.6 | |
| MCHC (g/dL) | 32.3 (0.9) | 37.4 (1.5) | 0.46 | |
| WBC (×103/µL) | 6.8 (1.8) | 6.7 (1.4) | 0.68 | |
| Neutrophils % | 57.3 (14.3) | 60.7 (7.3) | 0.37 | |
| Lymphocytes % | 32.5 (7.9) | 38.8 (8.5) | 0.24 | |
| Platelets (×103/µL) | 187.3 (37.1) | 208.8 (16.9) | 0.12 | |
| ALP (U/L) | 213.0 (43.0) | 195.2 (35.1) | 0.58 | |
| ALT (U/L) | 22.0 (7.2) | 19.3 (6.6) | 0.38 | |
| AST (U/L) | 23.8 (11.0) | 20.2 (8.8) | 0.26 | |
| UA (mg/dL) | 5.9 (2.5) | 6.7 (1.8) | 0.33 |
| Treatment | Parameter | Baseline | Post-Treatment | p-Value § |
|---|---|---|---|---|
| Placebo | NK | 5.3 (1.9) ǂ | 6.0 (1.7) ǂ | 0.62 |
| NKT | 4.5 (1.6) ǂ | 5.6 (2.4) ǂ | 0.44 | |
| NK CD-107 a | 45.3 (12.0) ǂ | 50.8 (19.5) ¥ | 0.38 | |
| NKT CD-107 a | 67.9 (15.6) ǂ | 75.5 (22.3) ǂ | 0.23 | |
| Biobran/MGN-3 (500 mg/day) for 3 months | NK | 6.1 (2.6) ǂ | 6.7 (1.7) ǂ | 0.60 |
| NKT | 3.1 (0.6) ǂ | 4.6 (2.5) ǂ | 0.25 | |
| NK CD-107 a | 49.5 (10.4) ǂ | 75.2 (6.6) ¥ | 0.004 * | |
| NKT CD-107 a | 70.6 (10.1) ǂ | 76.9 (9.8) ǂ | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsaid, A.F.; Agrawal, S.; Agrawal, A.; Ghoneum, M. Dietary Supplementation with Biobran/MGN-3 Increases Innate Resistance and Reduces the Incidence of Influenza-like Illnesses in Elderly Subjects: A Randomized, Double-Blind, Placebo-Controlled Pilot Clinical Trial. Nutrients 2021, 13, 4133. https://doi.org/10.3390/nu13114133
Elsaid AF, Agrawal S, Agrawal A, Ghoneum M. Dietary Supplementation with Biobran/MGN-3 Increases Innate Resistance and Reduces the Incidence of Influenza-like Illnesses in Elderly Subjects: A Randomized, Double-Blind, Placebo-Controlled Pilot Clinical Trial. Nutrients. 2021; 13(11):4133. https://doi.org/10.3390/nu13114133
Chicago/Turabian StyleElsaid, Ahmed F., Sudhanshu Agrawal, Anshu Agrawal, and Mamdooh Ghoneum. 2021. "Dietary Supplementation with Biobran/MGN-3 Increases Innate Resistance and Reduces the Incidence of Influenza-like Illnesses in Elderly Subjects: A Randomized, Double-Blind, Placebo-Controlled Pilot Clinical Trial" Nutrients 13, no. 11: 4133. https://doi.org/10.3390/nu13114133
APA StyleElsaid, A. F., Agrawal, S., Agrawal, A., & Ghoneum, M. (2021). Dietary Supplementation with Biobran/MGN-3 Increases Innate Resistance and Reduces the Incidence of Influenza-like Illnesses in Elderly Subjects: A Randomized, Double-Blind, Placebo-Controlled Pilot Clinical Trial. Nutrients, 13(11), 4133. https://doi.org/10.3390/nu13114133