
Impact of Oropharyngeal Administration of Colostrum in Preterm Newborns’ Oral Microbiome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Study Design
2.2. Group Definition and Oropharyngeal Administration of Colostrum Protocol
2.3. Sample Collection and DNA Isolation
2.4. 16S rRNA Gene Sequence Processing
2.5. Data Analysis
2.6. Statistical Analyses
2.7. Data Deposition
3. Results
3.1. Descriptive and Clinical Data
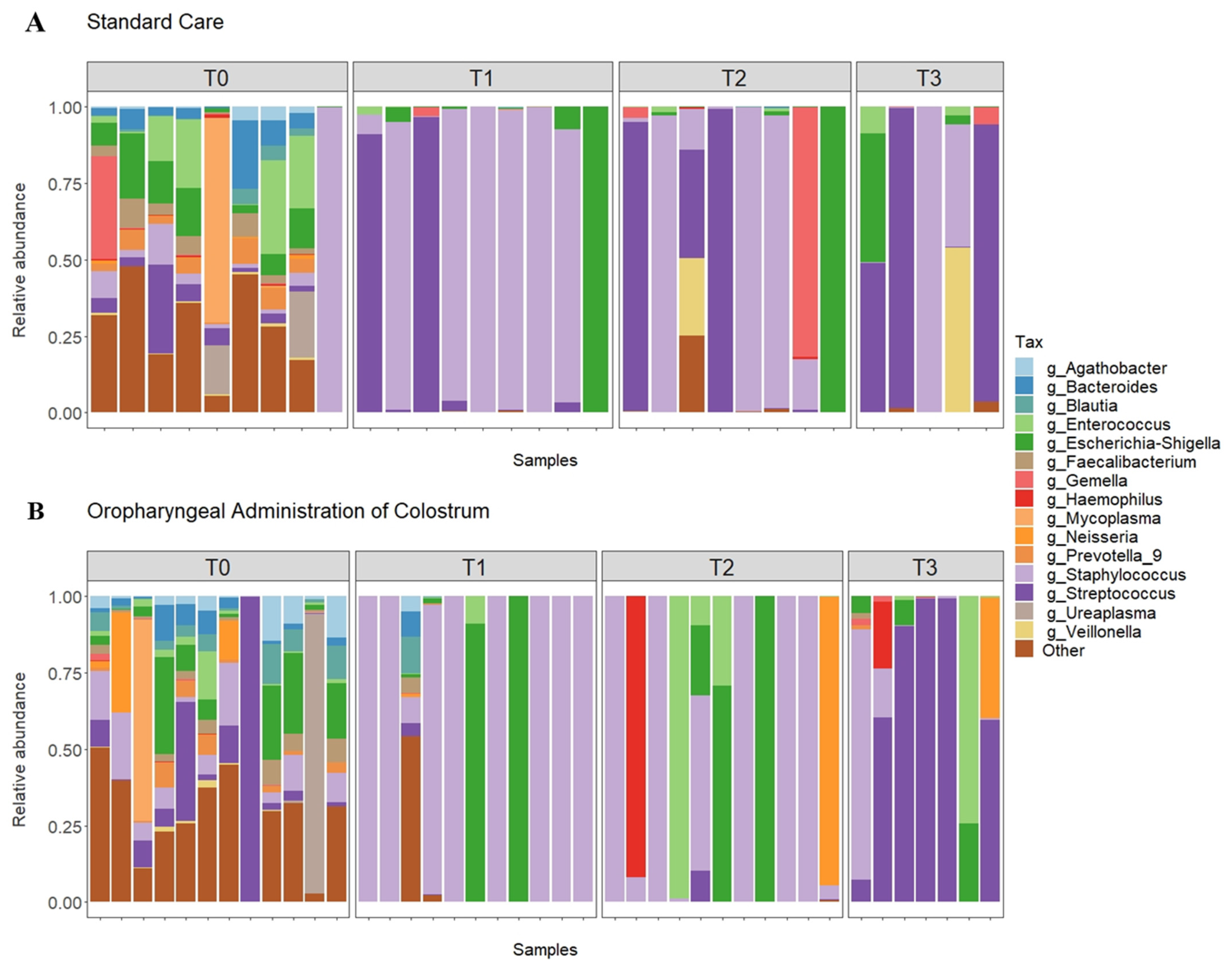
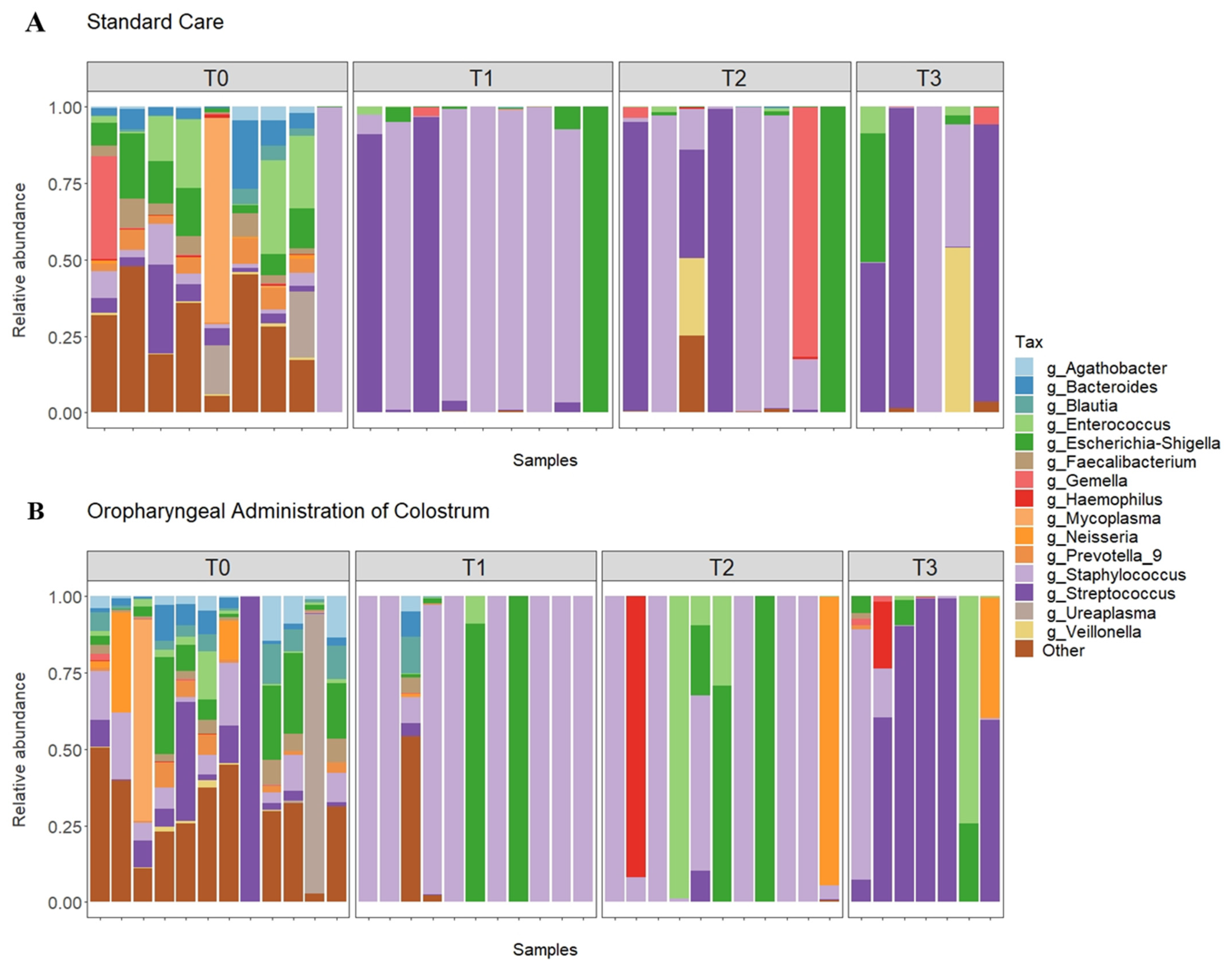
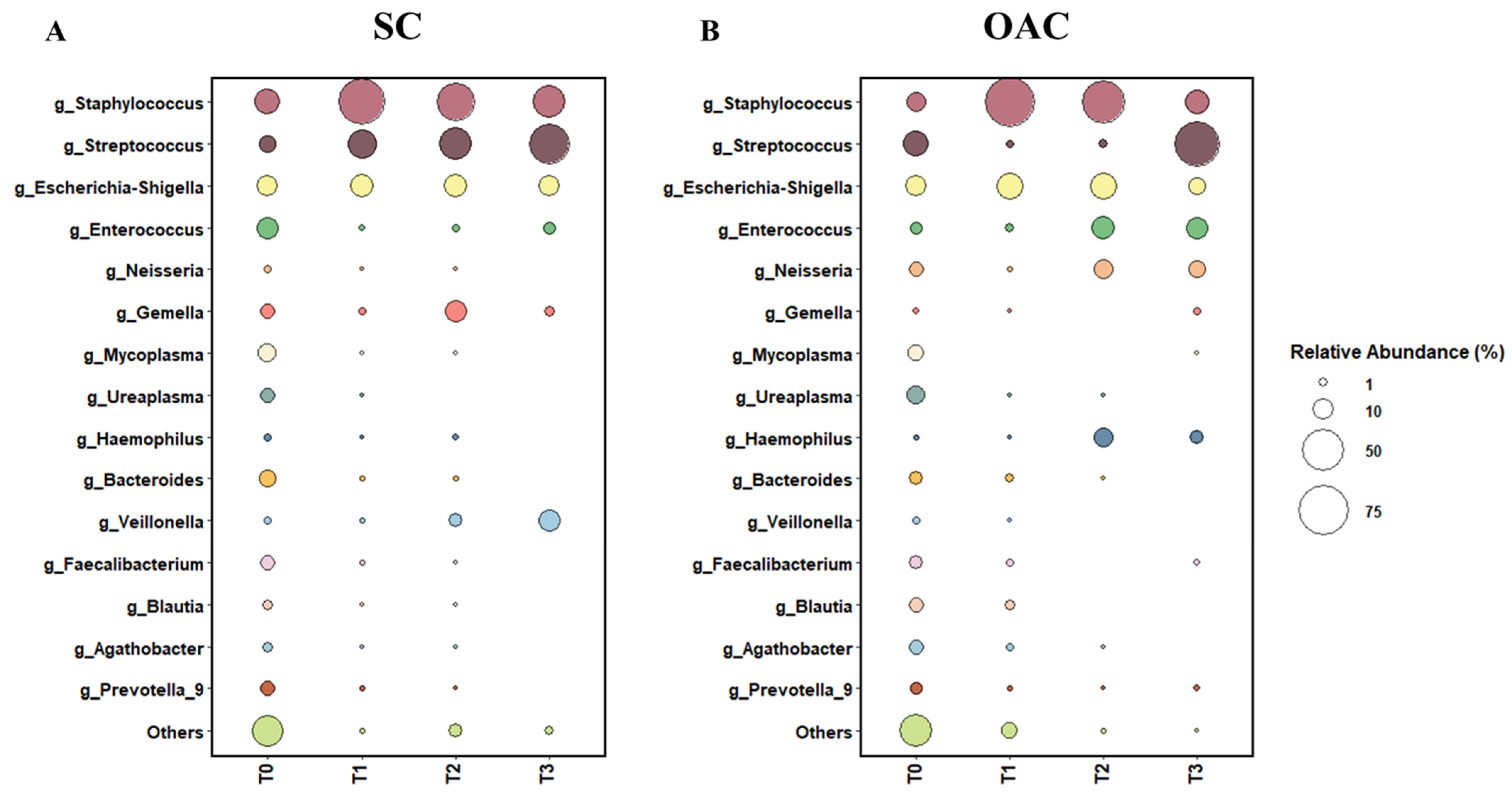
3.2. Relative Abundance
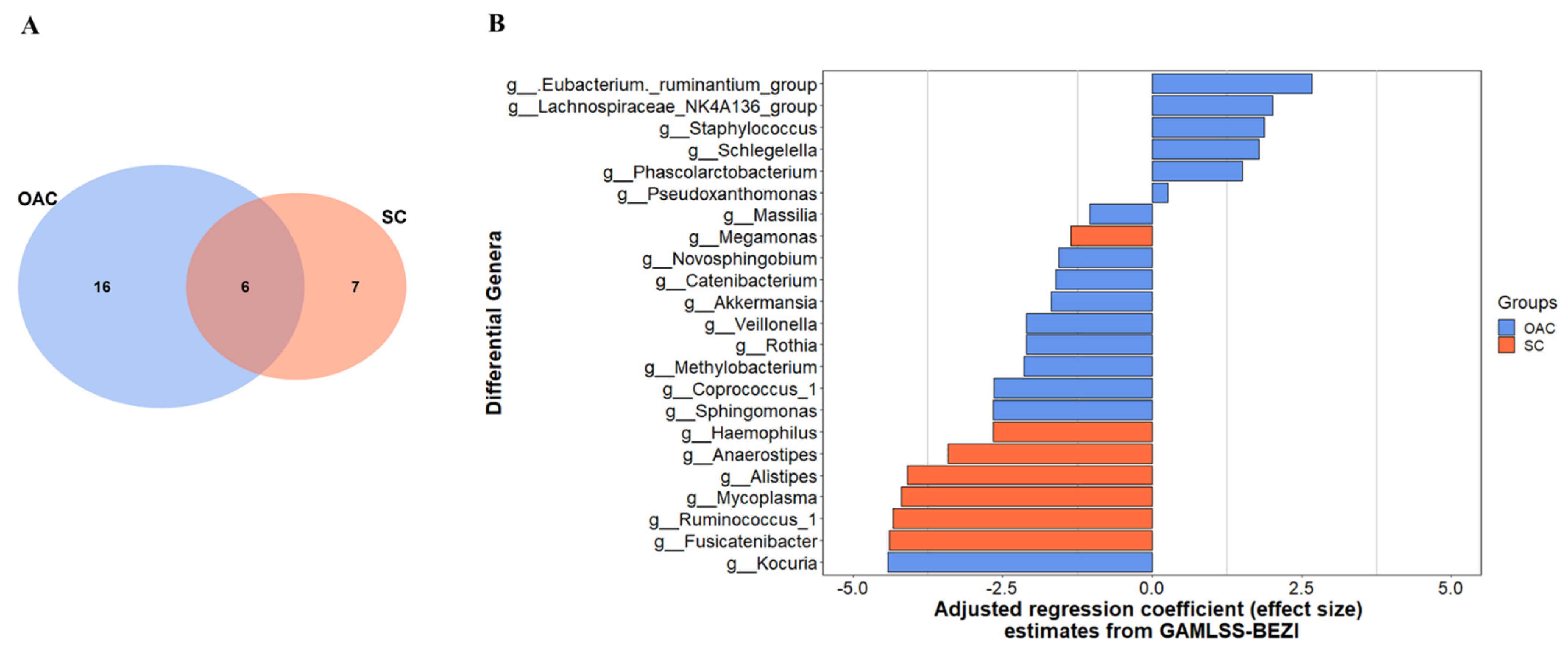
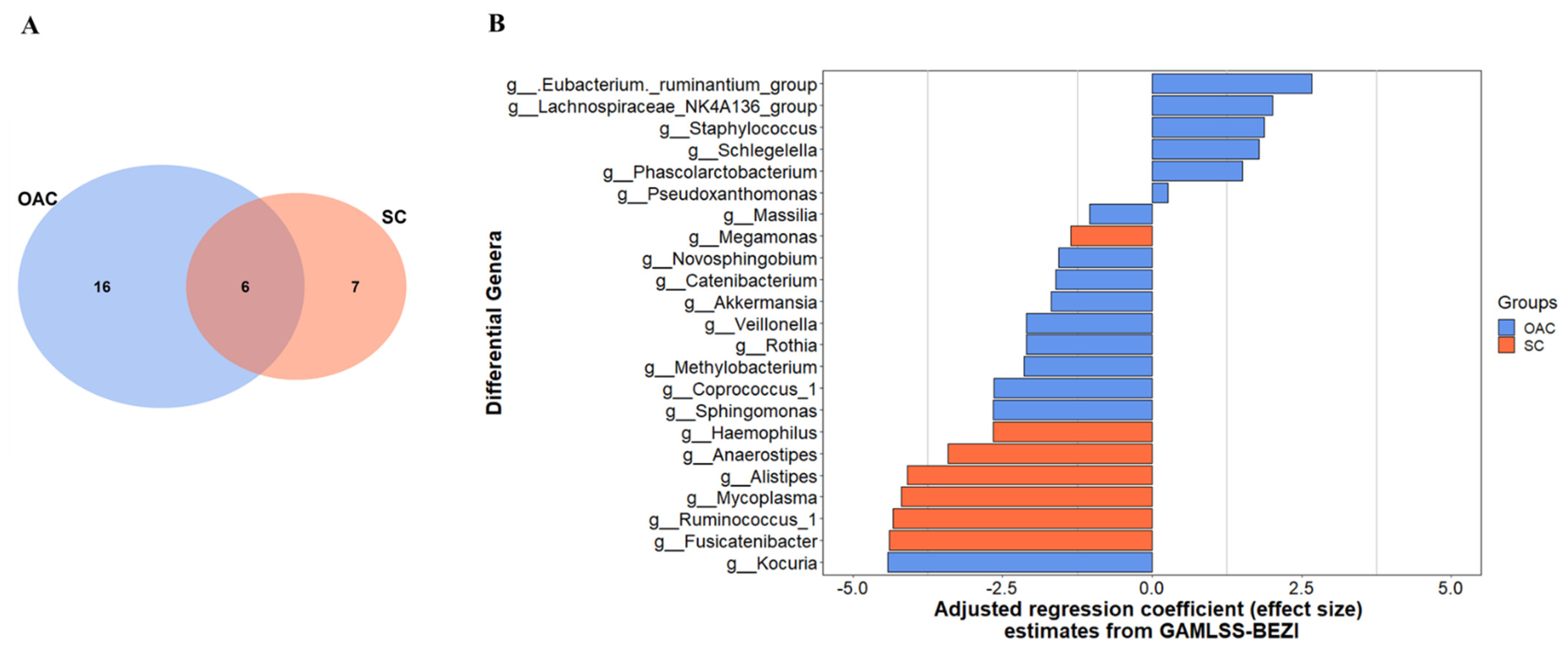
3.3. Comparison between Groups after Administration of Colostrum
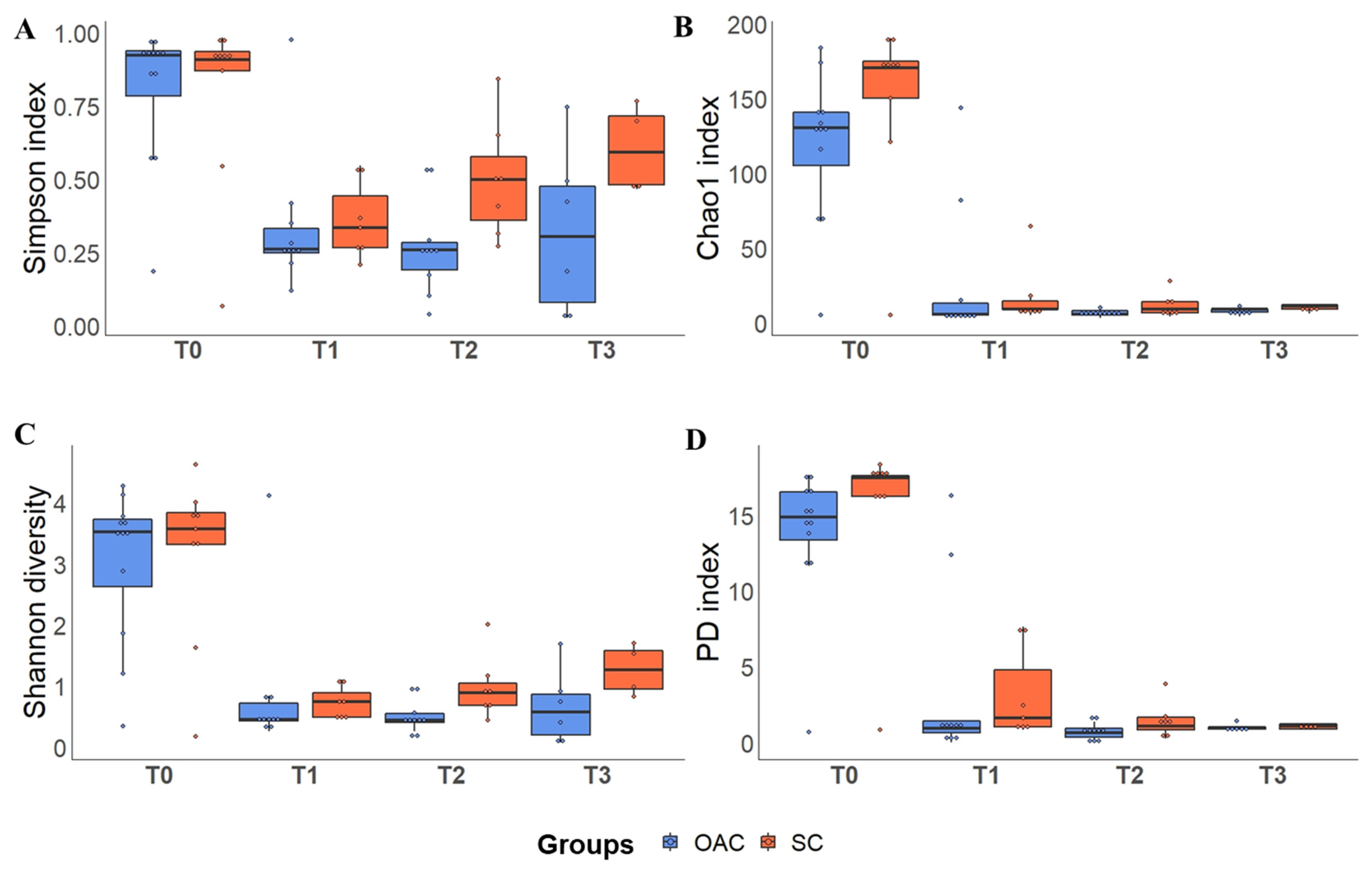
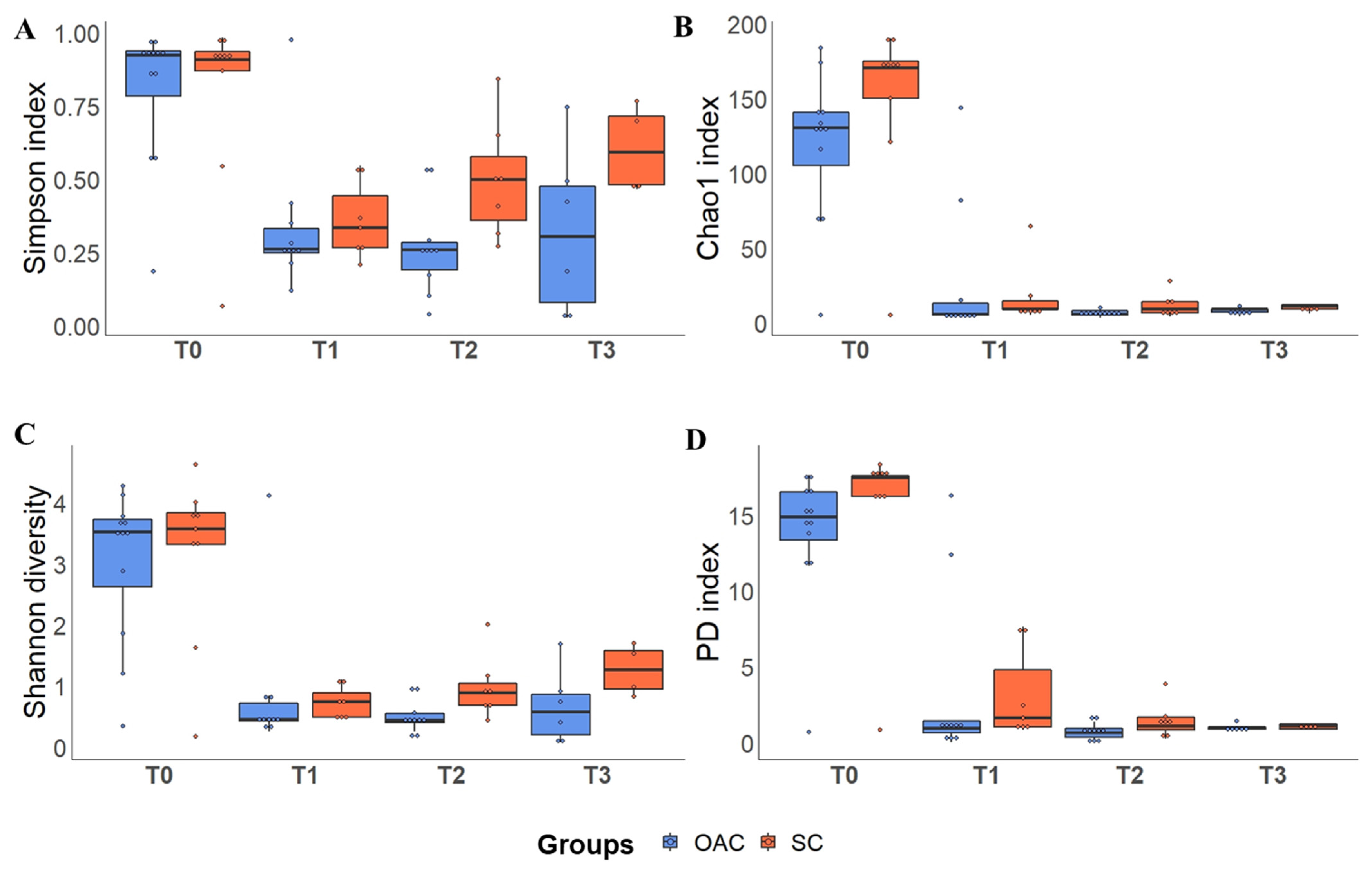
3.4. Alpha Diversity
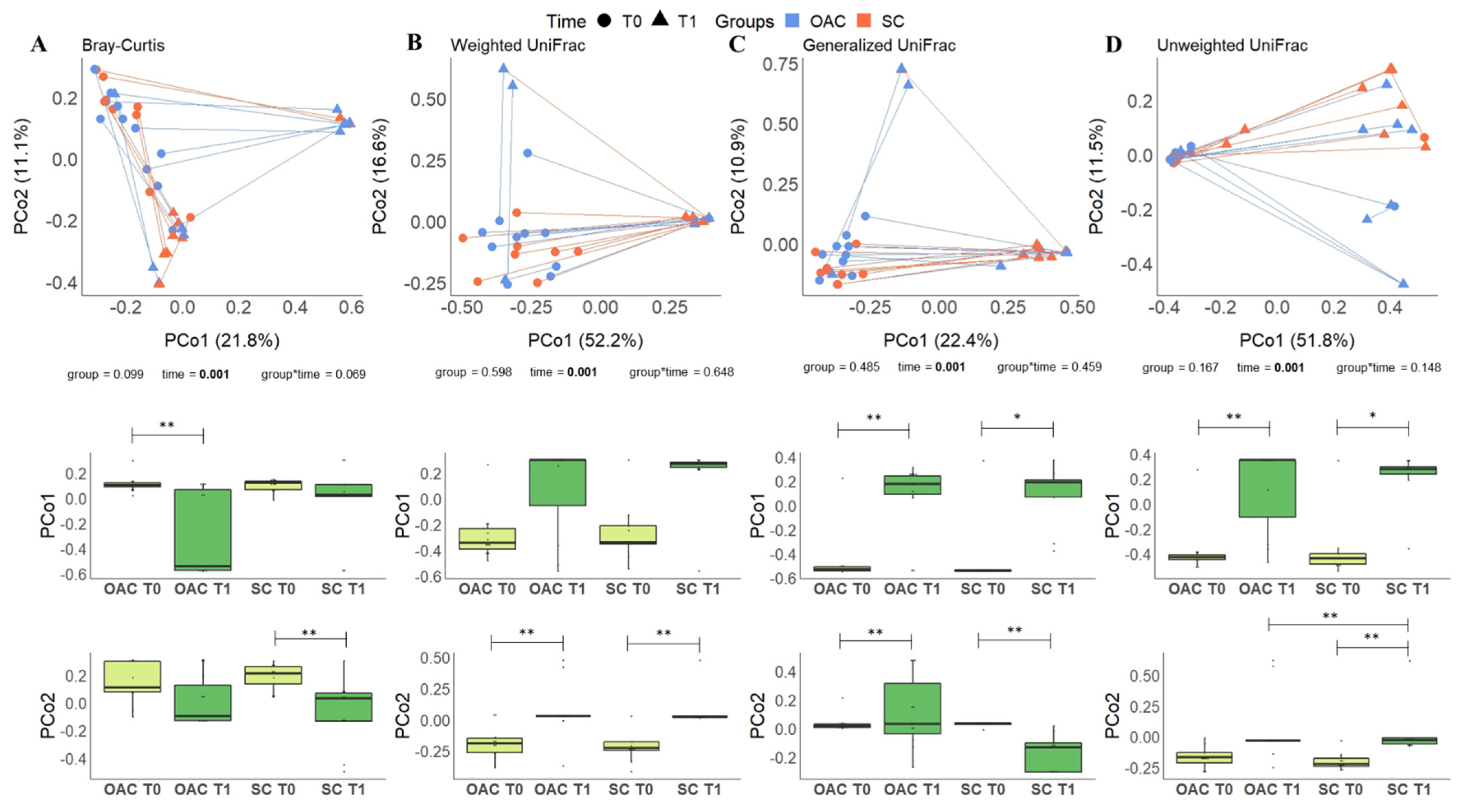
3.5. Beta Diversity between Groups and after the Intervention
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gosalbes, M.J.; Llop, S.; Vallès, Y.; Moya, A.; Ballester, F.; Francino, M.P. Meconium microbiota types dominated by lactic acid or enteric bacteria are differentially associated with maternal eczema and respiratory problems in infants. Clin. Exp. Allergy 2013, 43, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Makino, H.; Cetinyurek Yavuz, A.; Ben-Amor, K.; Roelofs, M.; Ishikawa, E.; Kubota, H.; Swinkels, S.; Sakai, T.; Oishi, K.; et al. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS ONE 2016, 11, e0158498. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, E.L.; Al-Shehri, S.S.; Cowley, D.M.; Liley, H.G.; Bansal, N.; Charles, B.G.; Shaw, P.N.; Duley, J.A.; Knox, C.L. The effect of breastmilk and saliva combinations on the in vitro growth of oral pathogenic and commensal microorganisms. Sci. Rep. 2018, 8, 15112. [Google Scholar] [CrossRef] [Green Version]
- Neu, J. The microbiome during pregnancy and early postnatal life. Semin. Fetal Neonatal Med. 2016, 21, 373–379. [Google Scholar] [CrossRef]
- Rautava, S. Early microbial contact, the breast milk microbiome and child health. J. Dev. Orig. Health Dis. 2016, 7, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shehri, S.S.; Sweeney, E.L.; Cowley, D.M.; Liley, H.G.; Ranasinghe, P.D.; Charles, B.G.; Shaw, P.N.; Vagenas, D.; Duley, J.A.; Knox, C.L. Deep sequencing of the 16S ribosomal RNA of the neonatal oral microbiome: A comparison of breast-fed and formula-fed infants. Sci. Rep. 2016, 6, 38309. [Google Scholar] [CrossRef] [Green Version]
- Böttcher, M.F.; Jenmalm, M.C.; Björkstén, B. Cytokine, chemokine and secretory IgA levels in human milk in relation to atopic disease and IgA production in infants. Pediatric Allergy Immunol. Off. Publ. Eur. Soc. Pediatric Allergy Immunol. 2003, 14, 35–41. [Google Scholar] [CrossRef]
- Claud, E.C.; Savidge, T.; Walker, W.A. Modulation of human intestinal epithelial cell IL-8 secretion by human milk factors. Pediatric Res. 2003, 53, 419–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, B.L.; Jilling, T.; Lapin, B.; Maheshwari, A.; Caplan, M.S. Maternal breast milk transforming growth factor-beta and feeding intolerance in preterm infants. Pediatric Res. 2014, 76, 386–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lönnerdal, B. Bioactive proteins in breast milk. J. Paediatr. Child. Health 2013, 49, 1–7. [Google Scholar] [CrossRef]
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. J. Pediatric Gastroenterol. Nutr. 2015, 60, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Albenzio, M.; Santillo, A.; Stolfi, I.; Manzoni, P.; Iliceto, A.; Rinaldi, M.; Magaldi, R. Lactoferrin levels in human milk after preterm and term delivery. Am. J. Perinatol. 2016, 33, 1085–1089. [Google Scholar] [CrossRef]
- Castellote, C.; Casillas, R.; Ramírez-Santana, C.; Pérez-Cano, F.J.; Castell, M.; Moretones, M.G.; López-Sabater, M.C.; Franch, À. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J. Nutr. 2011, 141, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Gidrewicz, D.A.; Fenton, T.R. A systematic review and meta-analysis of the nutrient content of preterm and term breast milk. BMC Pediatrics 2014, 14, 216. [Google Scholar] [CrossRef] [Green Version]
- W.H.O. Preterm Birth. Available online: https://www.who.int/en/news-room/fact-sheets/detail/preterm-birth (accessed on 11 October 2019).
- Ewaschuk, J.B.; Unger, S.; O’Connor, D.L.; Stone, D.; Harvey, S.; Clandinin, M.T.; Field, C.J. Effect of pasteurization on selected immune components of donated human breast milk. J. Perinatol. 2011, 31, 593–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertino, E.; Coppa, G.V.; Giuliani, F.; Coscia, A.; Gabrielli, O.; Sabatino, G.; Sgarrella, M.; Testa, T.; Zampini, L.; Fabris, C. Effects of holder pasteurization on human milk oligosaccharides. Int. J. Immunopathol. Pharmacol. 2008, 21, 381–385. [Google Scholar] [CrossRef] [Green Version]
- German, J.B.; Freeman, S.L.; Lebrilla, C.B.; Mills, D.A. Human milk oligosaccharides: Evolution, structures and bioselectivity as substrates for intestinal bacteria. In Personalized Nutrition for the Diverse Needs of Infants and Children; KARGER: Basel, Switzerland, 2008; Volume 62, pp. 205–222. [Google Scholar]
- Gephart, S.M.; Weller, M. Colostrum as oral immune therapy to promote neonatal health. Adv. Neonatal Care 2014, 14, 44–51. [Google Scholar] [CrossRef]
- Rodriguez, N.A.; Meier, P.P.; Groer, M.W.; Zeller, J.M.; Engstrom, J.L.; Fogg, L. A pilot study to determine the safety and feasibility of oropharyngeal administration of own mother’s colostrum to extremely low-birth-weight infants. Adv. Neonatal Care 2010, 10, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, H.-S.; Jung, Y.H.; Choi, K.Y.; Shin, S.H.; Kim, E.-K.; Choi, J.-H. Oropharyngeal colostrum administration in extremely premature infants: An RCT. Pediatrics 2015, 135, e357–e366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Fernandez, J.; Sánchez-Martínez, B.; Serrano-López, L.; Martín-Álvarez, E.; Diaz-Castro, J.; Peña-Caballero, M.; Martín-Peregrina, F.; Alonso-Moya, M.; Maldonado-Lozano, J.; Ochoa, J.J.; et al. Enhancement of immune response mediated by oropharyngeal colostrum administration in preterm neonates. Pediatric Allergy Immunol. 2018, 30, 13008. [Google Scholar] [CrossRef]
- Almeida, J.A.G. Capítulo 17—Pasteurização. In Banco de Leite Humano: Funcionamento, Prevenção e Controle de Riscos; Anvisa: Brasília, Brazil, 2007; p. 156. ISBN 978-85-88233-28-7. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package: Ordination methods, diversity analysis and other functions for community and vegetation ecologists. R Package Ver 2016. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 23 November 2021).
- Lahti, L.; Shetty, S. Introduction to the Microbiome R Package. Available online: http://bioconductor.statistik.tu-dortmund.de/packages/3.6/bioc/vignettes/microbiome/inst/doc/vignette.html (accessed on 4 July 2021).
- Hadley, W. ggplot2 Elegant Graphics for Data Analysis, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Pan, W. Akaike’s information criterion in generalized estimating equations. Biometrics 2001, 57, 120–125. [Google Scholar] [CrossRef]
- Oh, S.; Carriere, K.C.; Park, T. Model diagnostic plots for repeated measures data using the generalized estimating equations approach. Comput. Stat. Data Anal. 2008, 53, 222–232. [Google Scholar] [CrossRef]
- Ho, N.T.; Li, F.; Wang, S.; Kuhn, L. metamicrobiomeR: An R package for analysis of microbiome relative abundance data using zero-inflated beta GAMLSS and meta-analysis across studies using random effects models. BMC Bioinform. 2019, 20, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, S.W.; Lee, J.E.; Ha, E.K.; Han, M.Y.; Lee, E.; Kim, C.; Lee, J.H.; Lee, S.W.; Ha, J.E.; et al. Breastmilk feeding during the first 4 to 6 months of age and childhood disease burden until 10 years of age. Nutrients 2021, 13, 2825. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.; Herdt, A.; Mejias-Cepeda, N.; Ladino, J.; Crowley, K.; Levy, P. Early provision of oropharyngeal colostrum leads to sustained breast milk feedings in preterm infants. Pediatrics Neonatol. 2017, 58, 534–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano-Keeler, J.; Azcarate-Peril, M.A.; Weitkamp, J.-H.; Slaughter, J.C.; McDonald, W.H.; Meng, S.; Latuga, M.S.; Wynn, J.L. Oral colostrum priming shortens hospitalization without changing the immunomicrobial milieu. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2017, 37, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, N.A.; Caplan, M.S. Oropharyngeal mother’s milk. Clin. Perinatol. 2019, 46, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Slomka, V.; Hernandez-Sanabria, E.; Herrero, E.R.; Zaidel, L.; Bernaerts, K.; Boon, N.; Quirynen, M.; Teughels, W. Nutritional stimulation of commensal oral bacteria suppresses pathogens: The prebiotic concept. J. Clin. Periodontol. 2017, 44, 344–352. [Google Scholar] [CrossRef]
- Fehr, K.; Moossavi, S.; Sbihi, H.; Boutin, R.C.T.; Bode, L.; Robertson, B.; Yonemitsu, C.; Field, C.J.; Becker, A.B.; Mandhane, P.J.; et al. Breastmilk feeding practices are associated with the co-occurrence of bacteria in mothers’ milk and the infant gut: The CHILD cohort study. Cell Host Microbe 2020, 28, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mushajiang, S.; Luo, B.; Tian, F.; Ni, Y.; Yan, W. The composition and concordance of lactobacillus populations of infant gut and the corresponding breast-milk and maternal gut. Front. Microbiol. 2020, 11, 2985. [Google Scholar] [CrossRef]
- Biagi, E.; Aceti, A.; Quercia, S.; Beghetti, I.; Rampelli, S.; Turroni, S.; Soverini, M.; Zambrini, A.V.; Faldella, G.; Candela, M.; et al. Microbial community dynamics in mother’s milk and infant’s mouth and gut in moderately preterm infants. Front. Microbiol. 2018, 9, 2512. [Google Scholar] [CrossRef] [Green Version]
- Sampaio-Maia, B.; Monteiro-Silva, F. Acquisition and maturation of oral microbiome throughout childhood: An update. Dent. Res. J. 2014, 11, 291–301. [Google Scholar]
- Kolenbrander, P.E.; Palmer, R.J.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell–cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef]
- Costello, E.K.; Carlisle, E.M.; Bik, E.M.; Morowitz, M.J.; Relman, D.A. Microbiome assembly across multiple body sites in low-birthweight infants. mBio 2013, 4, e00782-13. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Schloss, P.D. Dynamics and associations of microbial community types across the human body. Nature 2014, 509, 357–360. [Google Scholar] [CrossRef]
- Hendricks-Muñoz, K.; Xu, J.; Parikh, H.; Xu, P.; Fettweis, J.; Kim, Y.; Louie, M.; Buck, G.; Thacker, L.; Sheth, N. Skin-to-skin care and the development of the preterm infant oral microbiome. Am. J. Perinatol. 2015, 32, 1205–1216. [Google Scholar]
- Sohn, K.; Kalanetra, K.M.; Mills, D.A.; Underwood, M.A. Buccal administration of human colostrum: Impact on the oral microbiota of premature infants. J. Perinatol. 2016, 36, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Bacigalupe, R.; García-Carral, C.; Boix-Amoros, A.; Argüello, H.; Silva, C.B.; de los Angeles Checa, M.; Mira, A.; Rodríguez, J.M. Microbiota of human precolostrum and its potential role as a source of bacteria to the infant mouth. Sci. Rep. 2019, 9, 8435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.E.; Niñonuevo, M.; Mills, D.A.; Lebrilla, C.B.; German, J.B. In vitro fermentation of breast milk oligosaccharides by Bifidobacterium infantis and Lactobacillus gasseri. Appl. Environ. Microbiol. 2006, 72, 4497–4499. [Google Scholar] [CrossRef] [Green Version]
- Gregory, K.E.; LaPlante, R.D.; Shan, G.; Kumar, D.V.; Gregas, M. Mode of birth influences preterm infant intestinal colonization with bacteroides over the early neonatal period. Adv. Neonatal Care Off. J. Natl. Assoc. Neonatal Nurses 2015, 15, 393. [Google Scholar] [CrossRef] [Green Version]
- Young, G.R.; Van Der Gast, C.J.; Smith, D.L.; Berrington, J.E.; Embleton, N.D.; Lanyon, C. Acquisition and development of the extremely preterm infant microbiota across multiple anatomical sites. J. Pediatric Gastroenterol. 2020, 70, 12–19. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | OAC Group (n = 11) | SC Group (n = 9) | Wald Chi-Square | p Value |
|---|---|---|---|---|
| Maternal age (years) # | 29.9 ± 4.8 | 27.1 ± 7.3 | 1.178 | 0.27 |
| Gestational age (weeks) # | 30.6 ± 2.3 | 31.4 ± 1.7 | 0.736 | 0.39 |
| Obstetric history § | 1.546 | 0.21 | ||
| Multiparous | 9 (81.8) | 5 (55.6) | ||
| Primiparous | 2 (18.2) | 4 (44.4) | ||
| Sex § | 0.884 | 0.34 | ||
| Female | 5 (45.5) | 6 (66.7) | ||
| Male | 6 (54.5) | 3 (33.3) | ||
| Type of delivery § | 0.020 | 0.88 | ||
| C-section | 7 (63.6) | 6 (66.7) | ||
| Vaginal | 4 (36.4) | 3 (33.3) | ||
| Sepsis § | 1.135 | 0.28 | ||
| Yes | 5 (45.5) | 2 (22.2) | ||
| No | 6 (54.5) | 7 (77.8) | ||
| Use of antibiotics on NICU £ | 0.050 | 0.82 | ||
| Yes | 9 (81.8)/16.2 (2–30) | 7 (77.8)/9.8 (2–27) | ||
| No | 2 (18.2) | 2 (22.2) | ||
| Parenteral nutrition time (days) | 16.1 ± 10.8 | 10.7 ± 11.4 | 1.309 | 0.25 |
| Birth weight (grams) | 1180.8 ± 249.2 | 1493.3 ± 344.4 | 6.159 | 0.013 * |
| Weight on the 7th day of life (grams) | 1140.1 ± 250.5 | 1416.0 ± 297.2 | 5.649 | 0.017 * |
| Weight on the 14th day of life (grams) | 1278.8 ± 319.5 | 1529.5 ± 281.6 | 3.760 | 0.052 |
| Weight on the 21st day of life (grams) ¥ | 1372.2 ± 298.3 | 1570.1 ± 307.8 | 2.069 | 0.15 |
| Weight at hospital discharge (grams) | 1963.0 ± 237.8 | 2055.0 ± 153.9 | 1.067 | 0.30 |
| Days in the NICU | 44.1 ± 32.2 | 23.4 ± 23.2 | 2.868 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortez, R.V.; Fernandes, A.; Sparvoli, L.G.; Padilha, M.; Feferbaum, R.; Neto, C.M.; Taddei, C.R. Impact of Oropharyngeal Administration of Colostrum in Preterm Newborns’ Oral Microbiome. Nutrients 2021, 13, 4224. https://doi.org/10.3390/nu13124224
Cortez RV, Fernandes A, Sparvoli LG, Padilha M, Feferbaum R, Neto CM, Taddei CR. Impact of Oropharyngeal Administration of Colostrum in Preterm Newborns’ Oral Microbiome. Nutrients. 2021; 13(12):4224. https://doi.org/10.3390/nu13124224
Chicago/Turabian StyleCortez, Ramon V., Andrea Fernandes, Luiz Gustavo Sparvoli, Marina Padilha, Rubens Feferbaum, Corintio Mariani Neto, and Carla R. Taddei. 2021. "Impact of Oropharyngeal Administration of Colostrum in Preterm Newborns’ Oral Microbiome" Nutrients 13, no. 12: 4224. https://doi.org/10.3390/nu13124224
APA StyleCortez, R. V., Fernandes, A., Sparvoli, L. G., Padilha, M., Feferbaum, R., Neto, C. M., & Taddei, C. R. (2021). Impact of Oropharyngeal Administration of Colostrum in Preterm Newborns’ Oral Microbiome. Nutrients, 13(12), 4224. https://doi.org/10.3390/nu13124224