
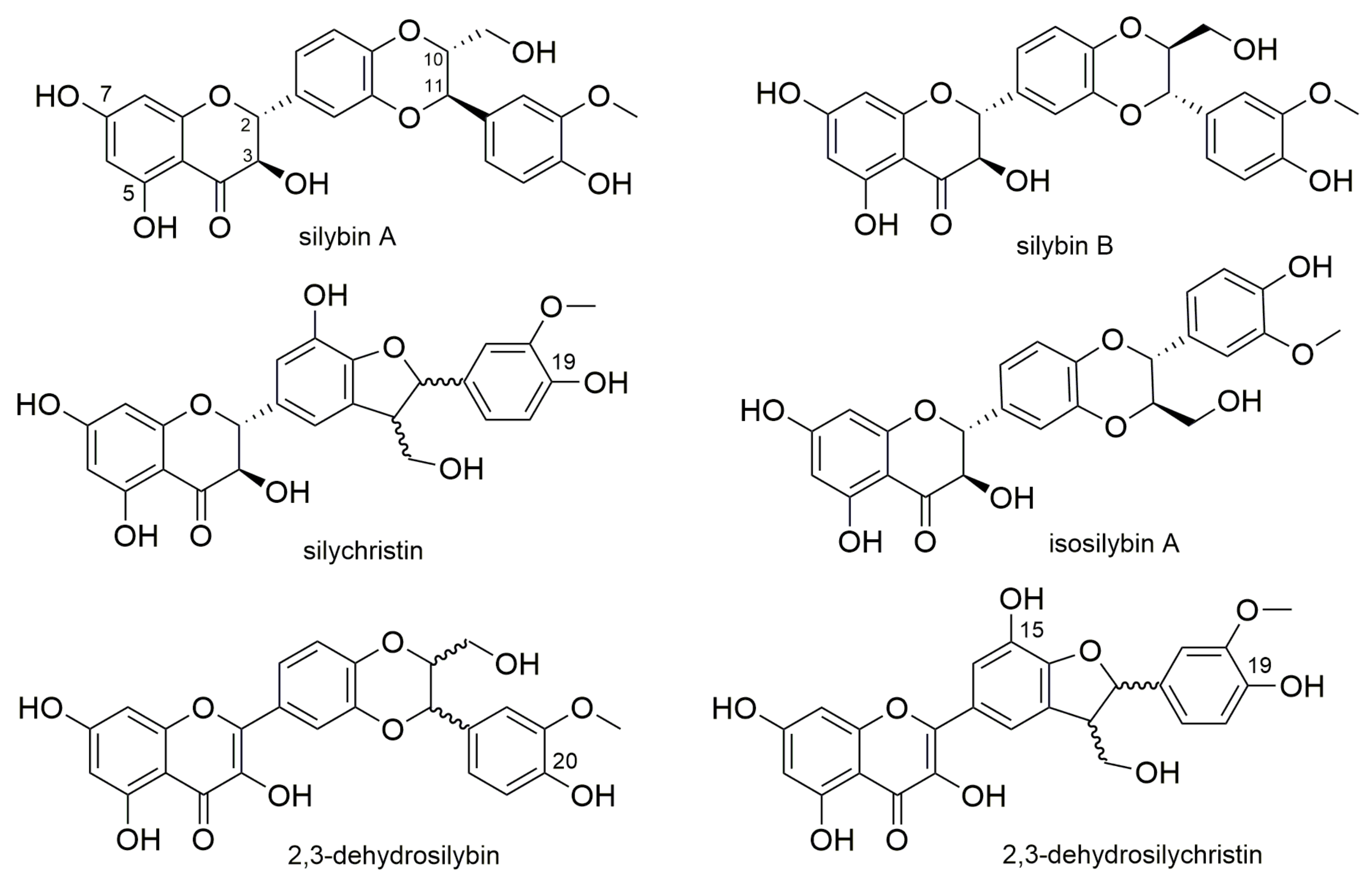
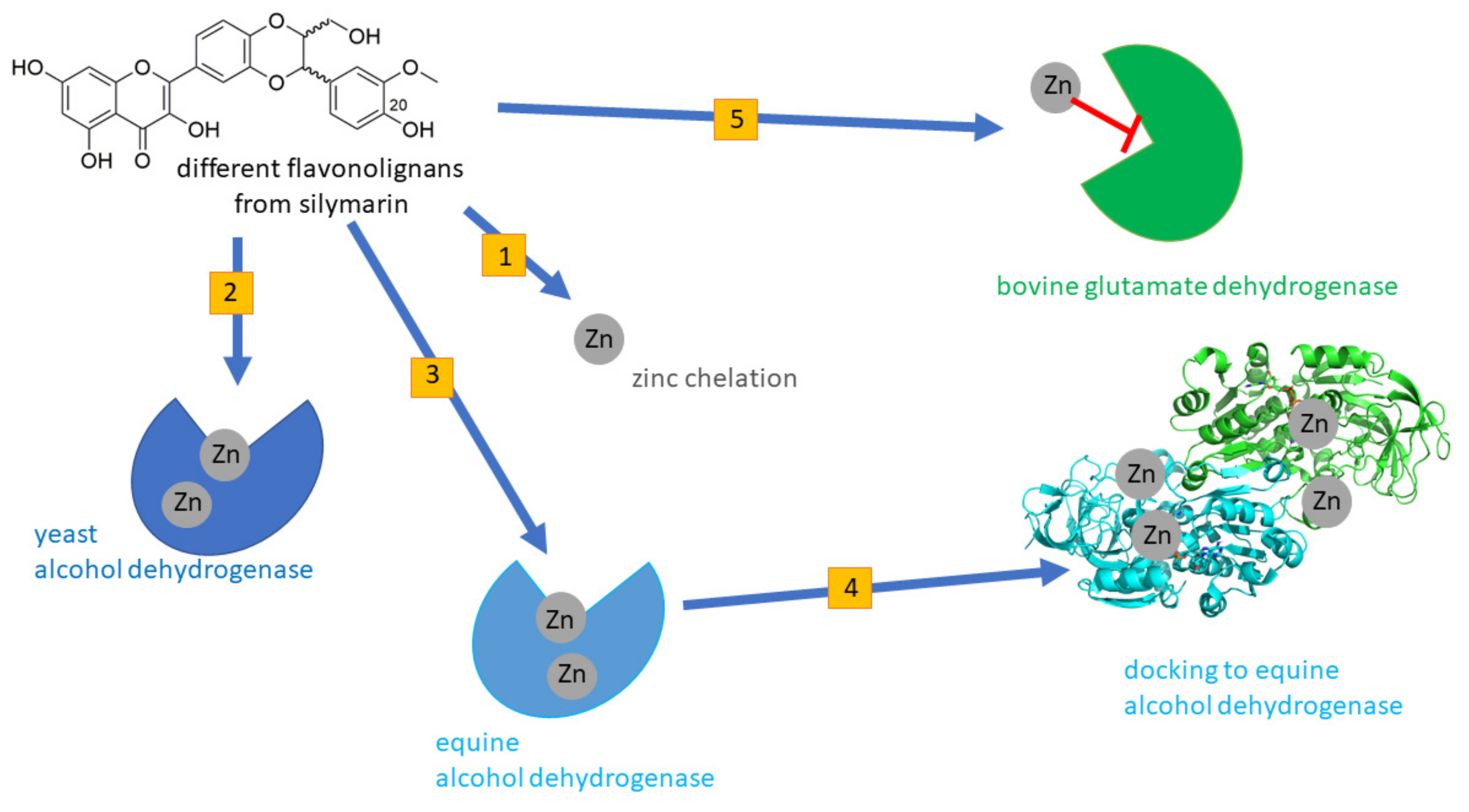
Silymarin Dehydroflavonolignans Chelate Zinc and Partially Inhibit Alcohol Dehydrogenase

 , , ,
, , ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
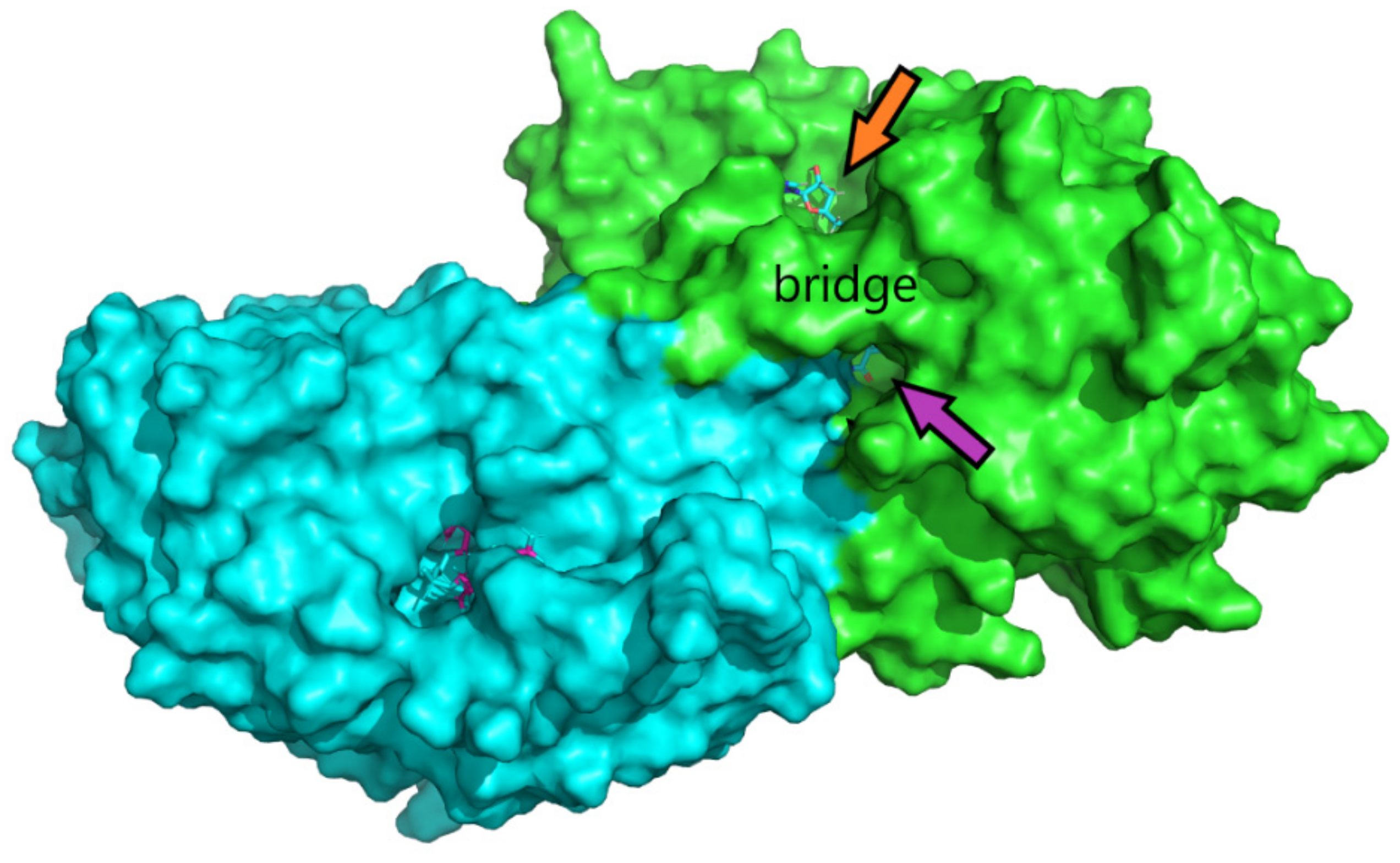{kind=link}
{kind=link}
{kind=link}
{kind=link}
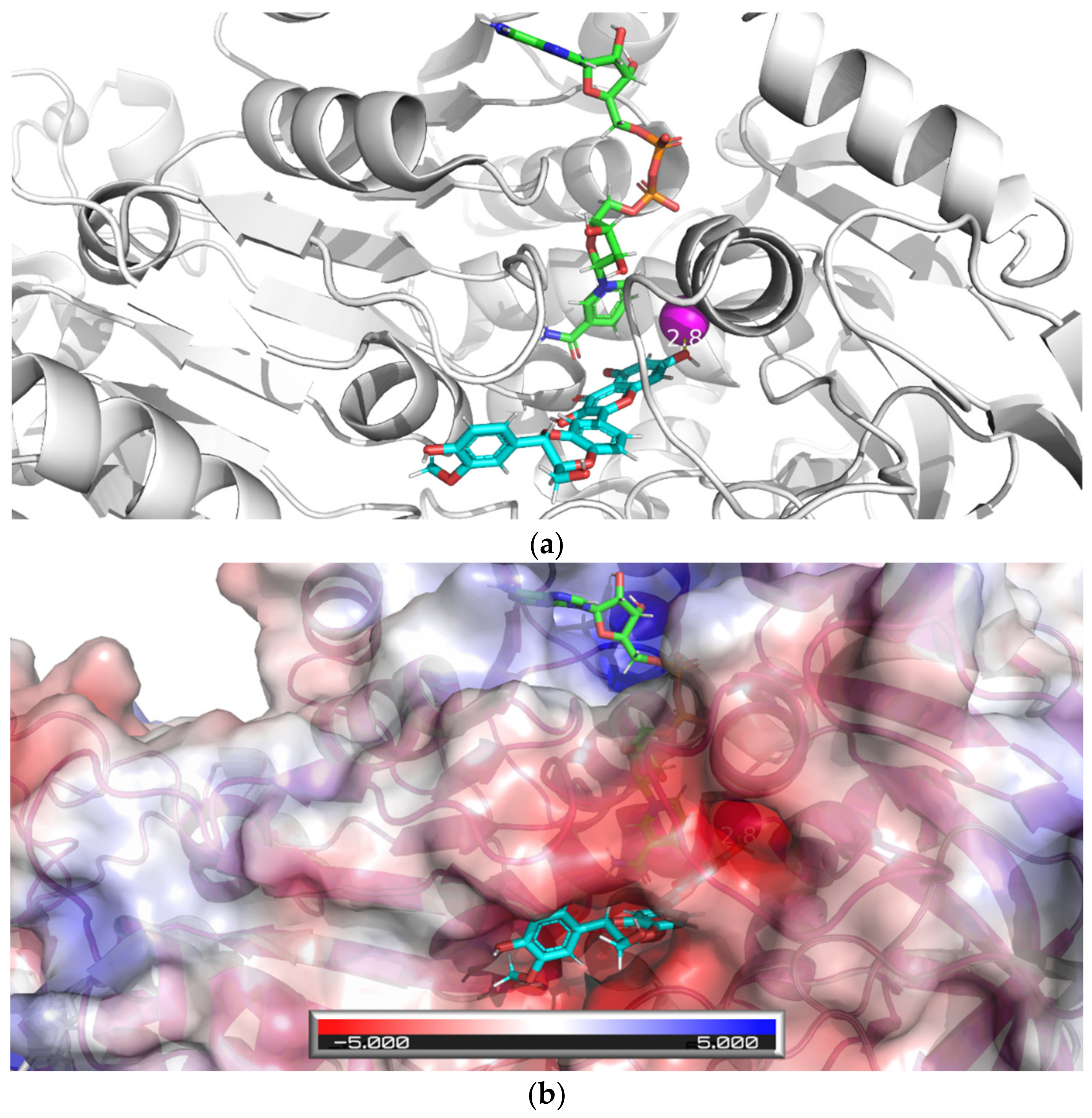{kind=link}
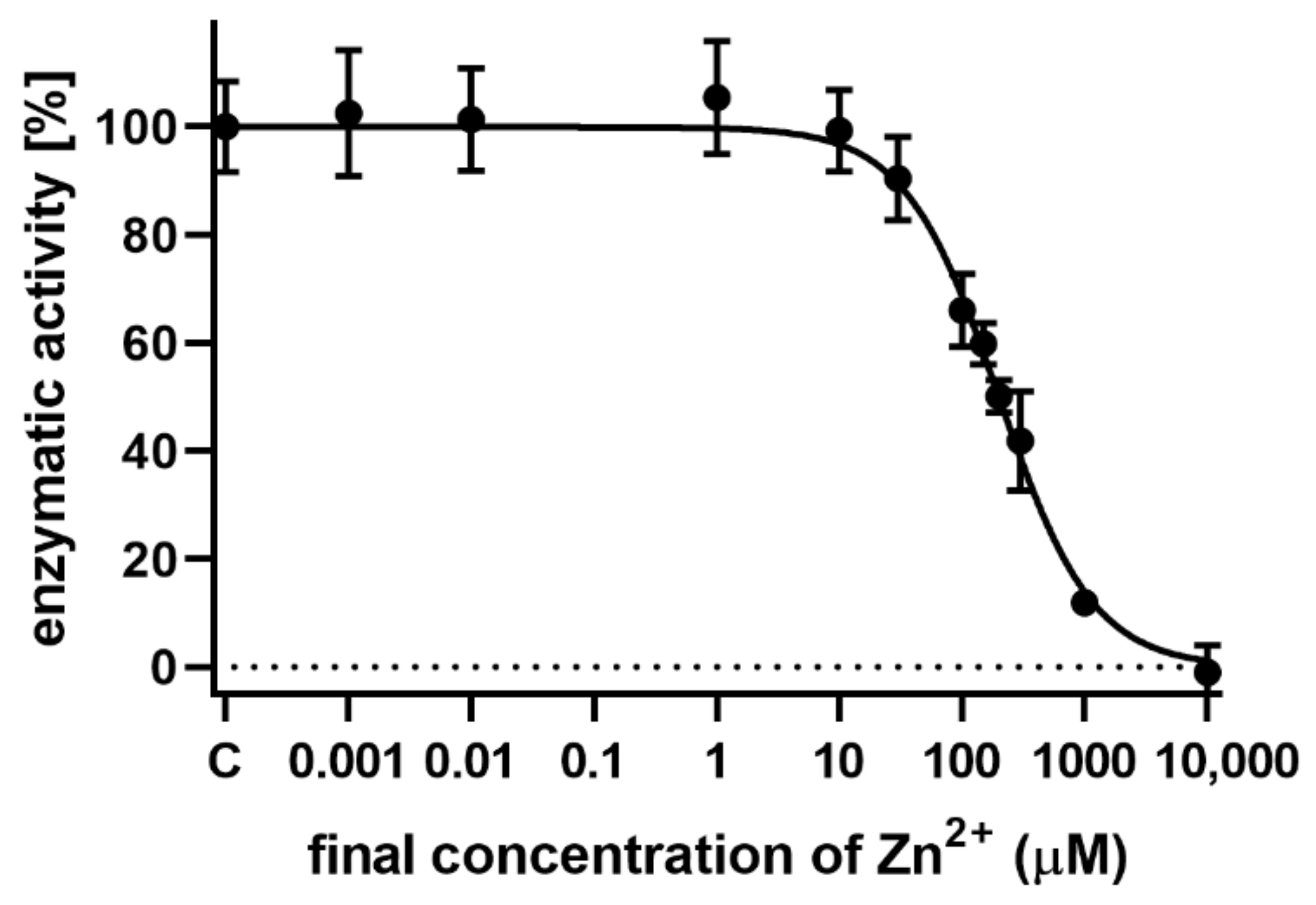{kind=link}
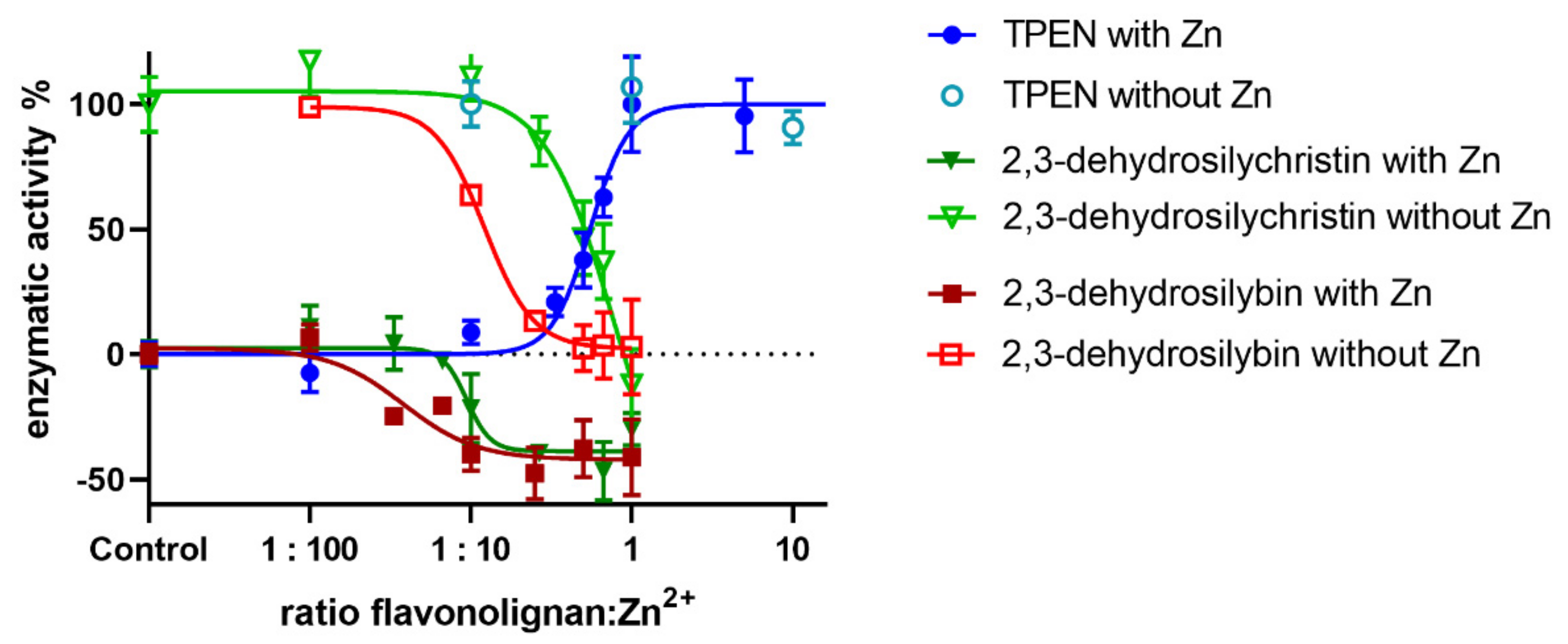{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Zn Chelation In Vitro
2.3. Alcohol Dehydrogenase
2.3.1. Yeast ADH
2.3.2. Equine ADH
2.4. Bovine Glutamate Dehydrogenase
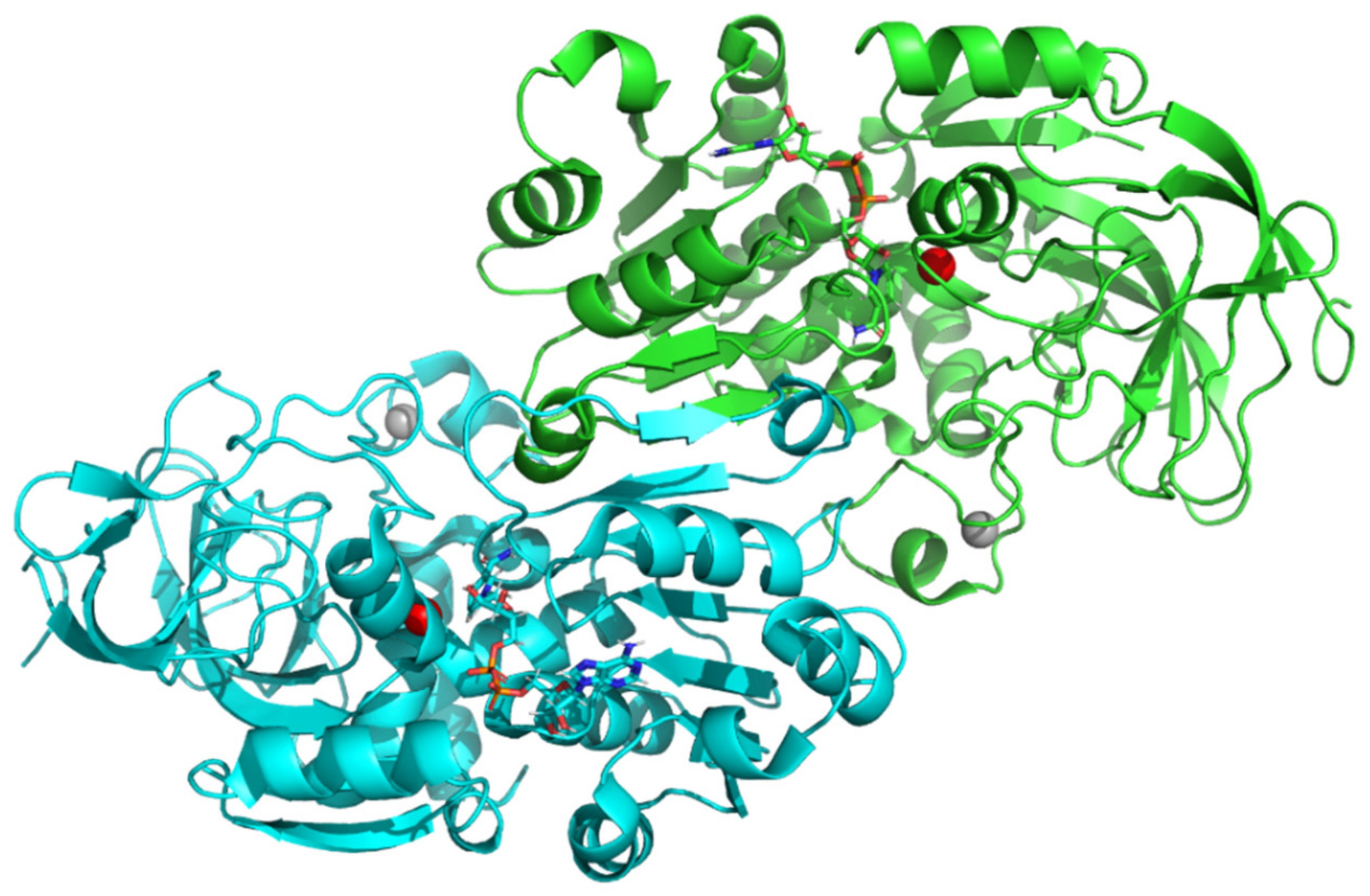
2.5. Molecular Docking In Silico
2.6. Calculations of Experimental Results
2.7. Statistical Analysis
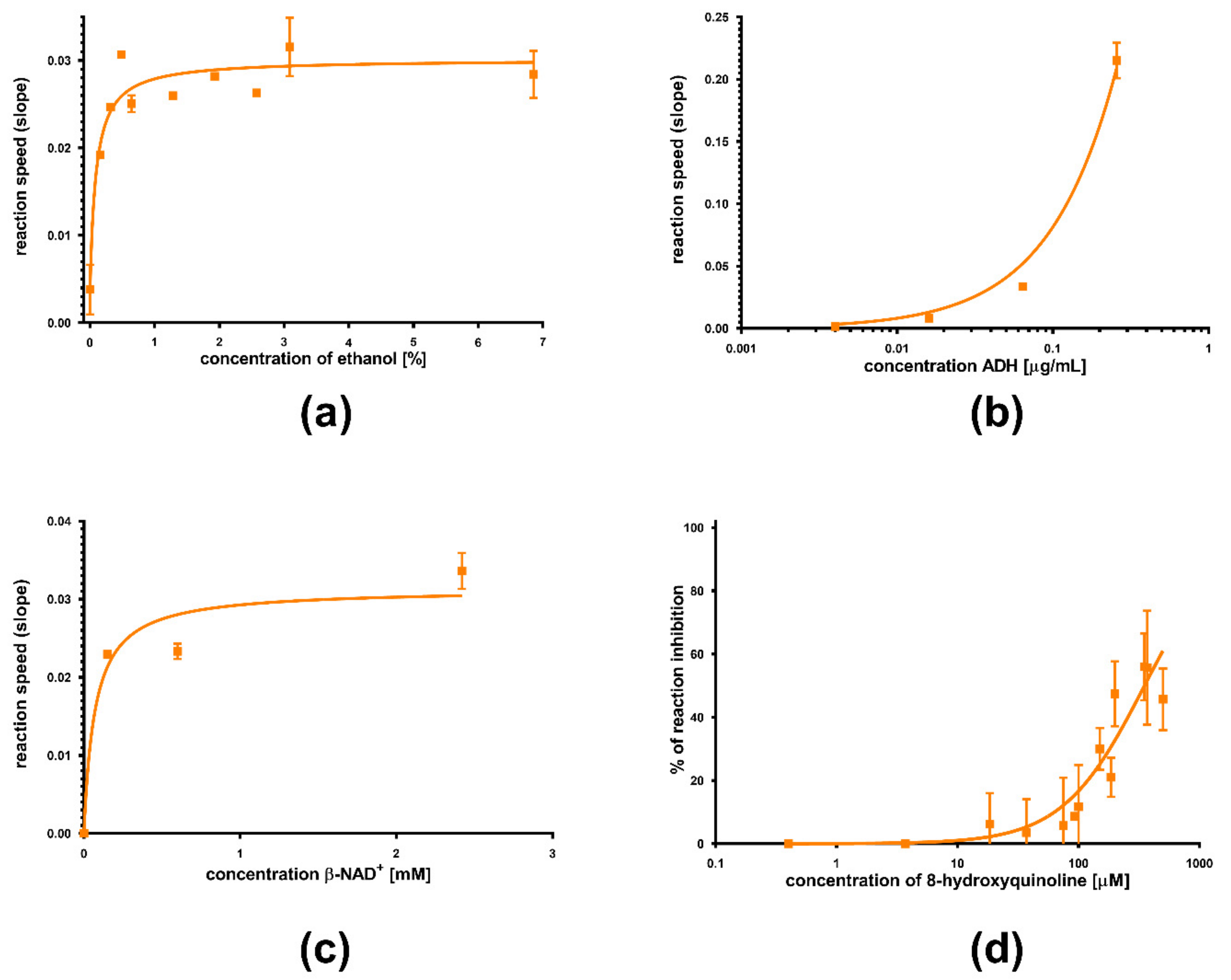
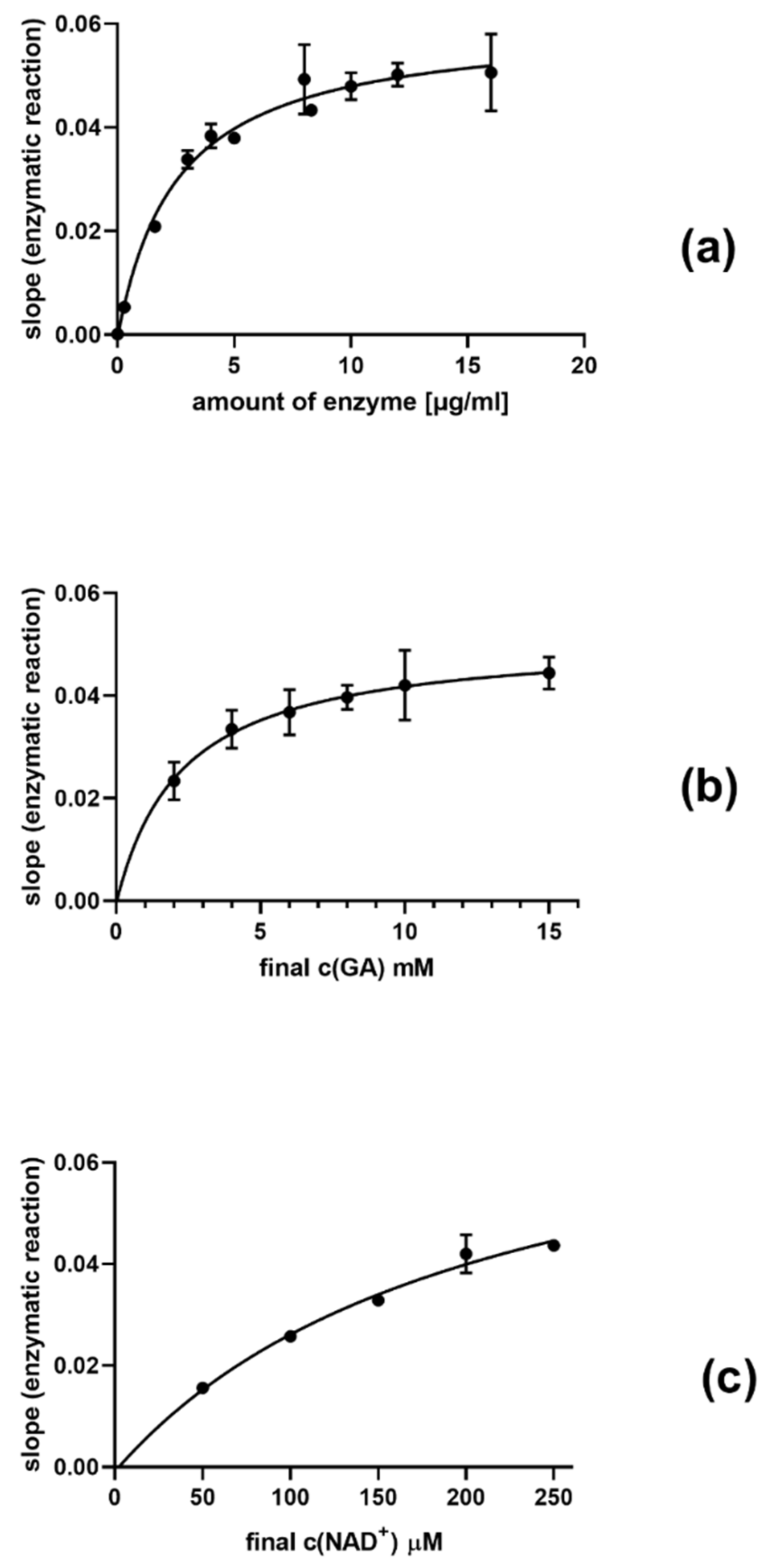
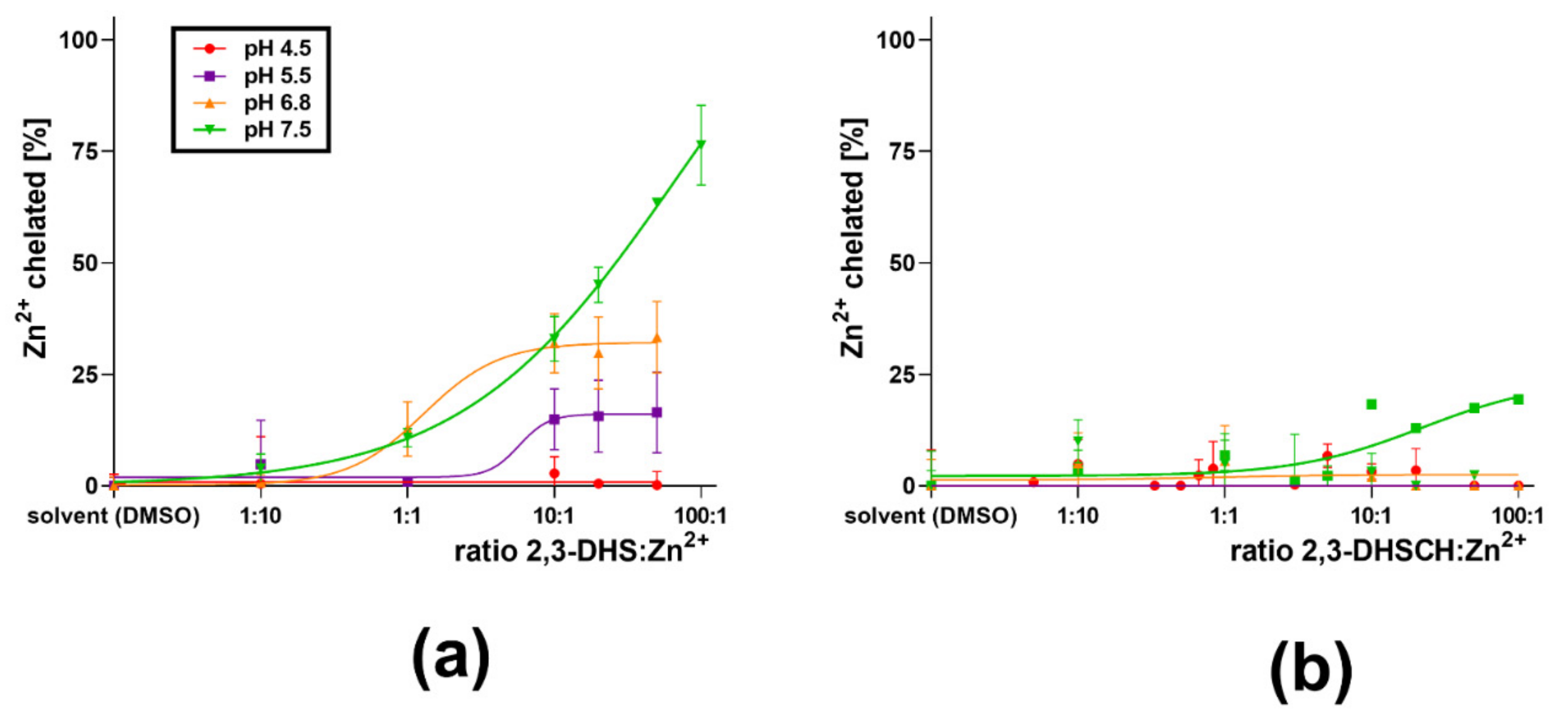
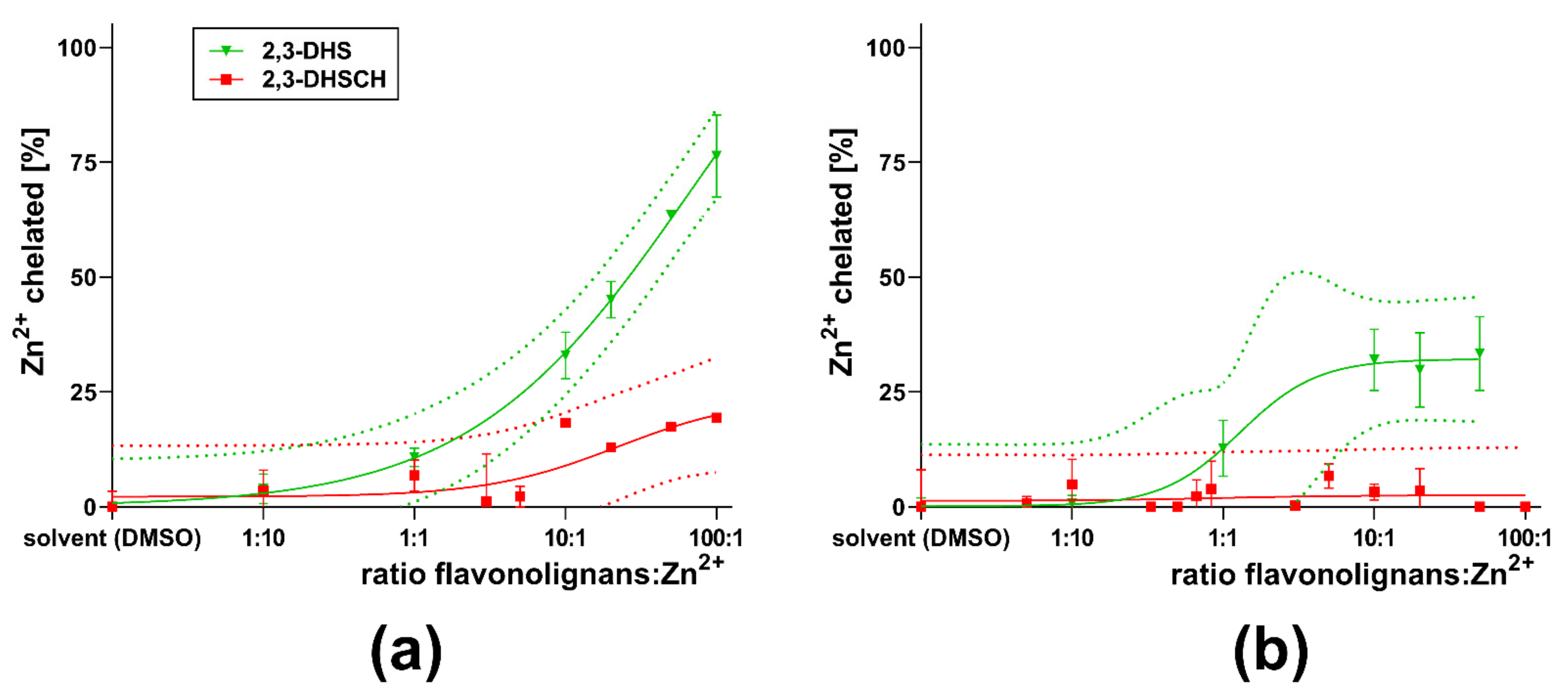
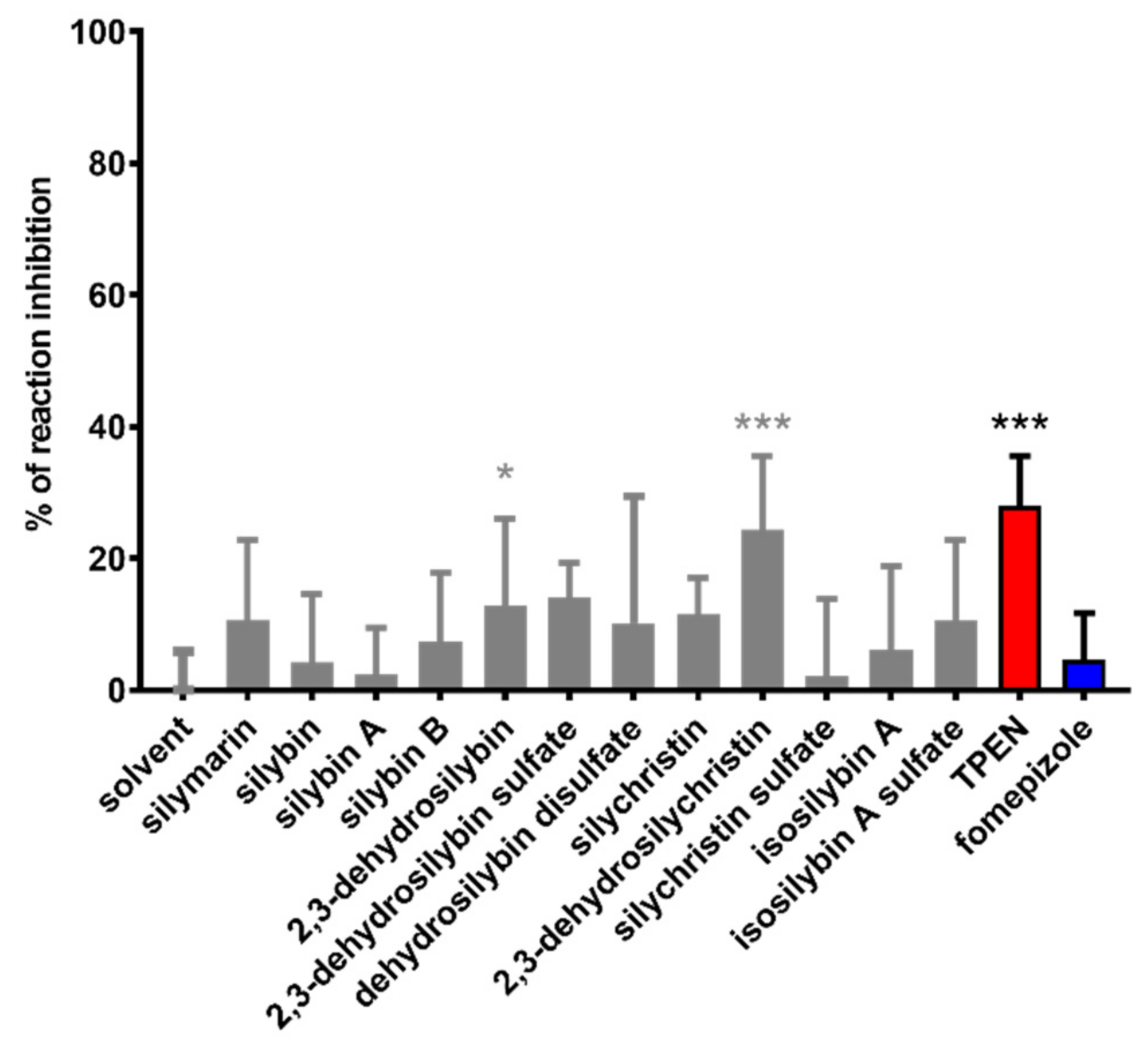
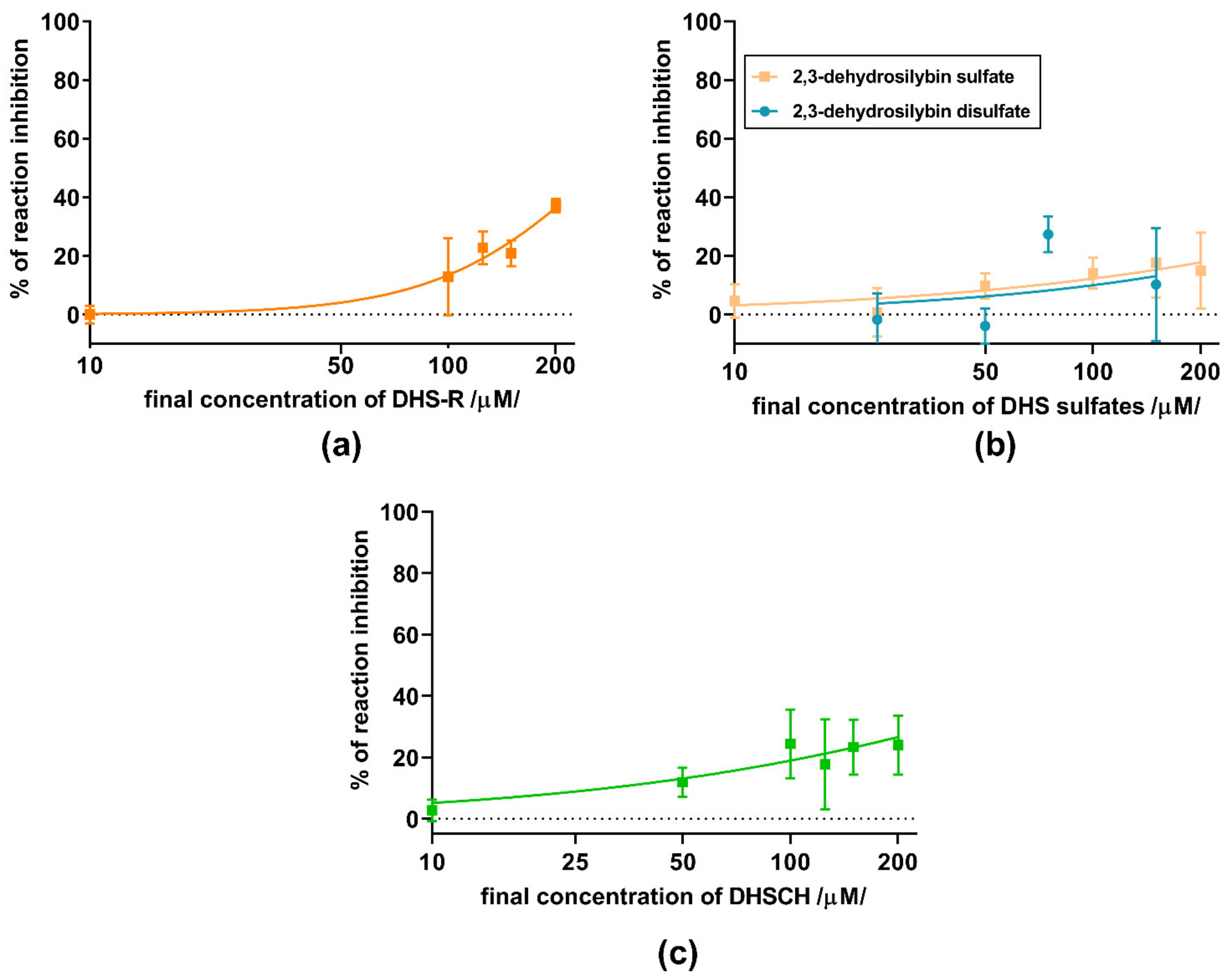
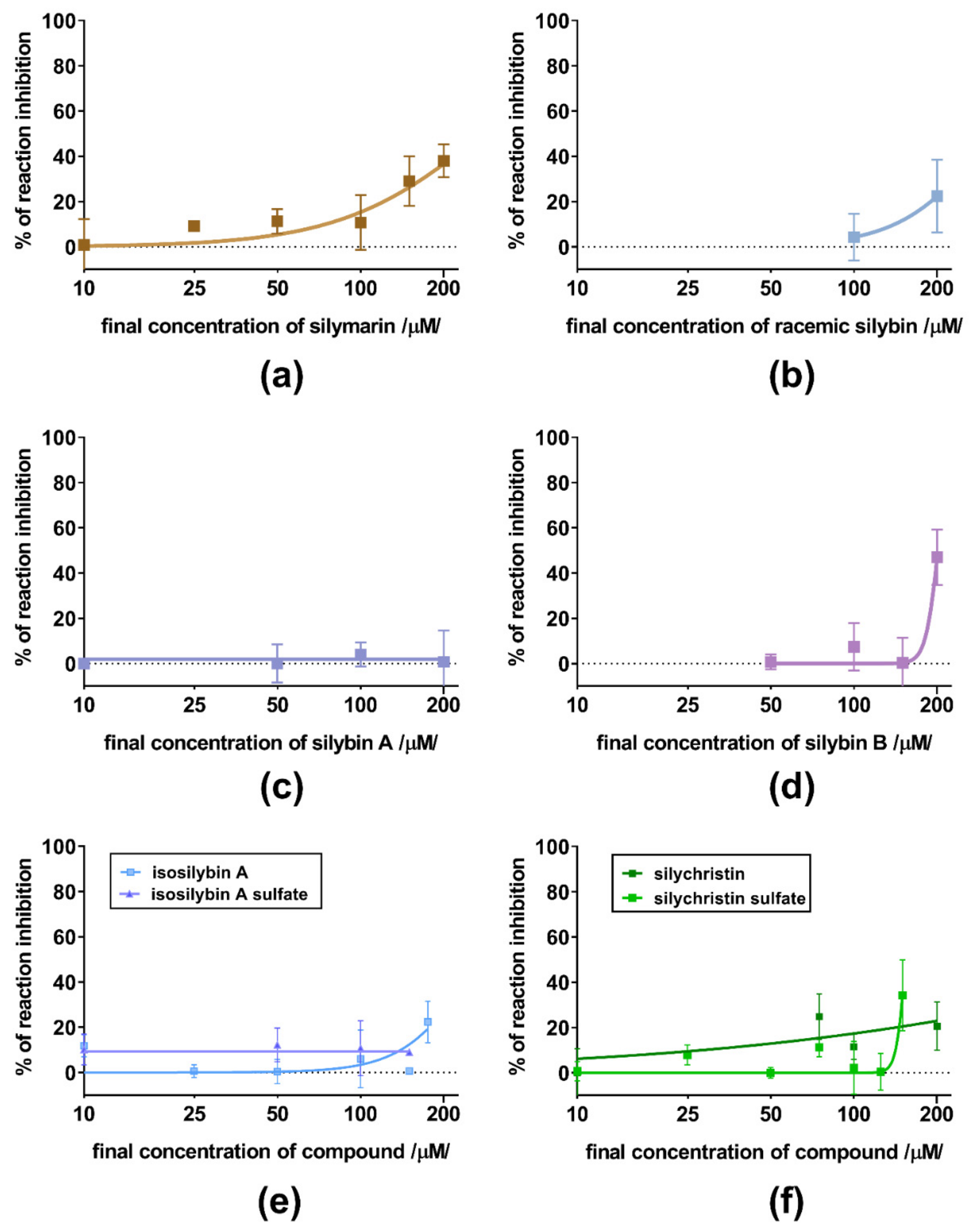
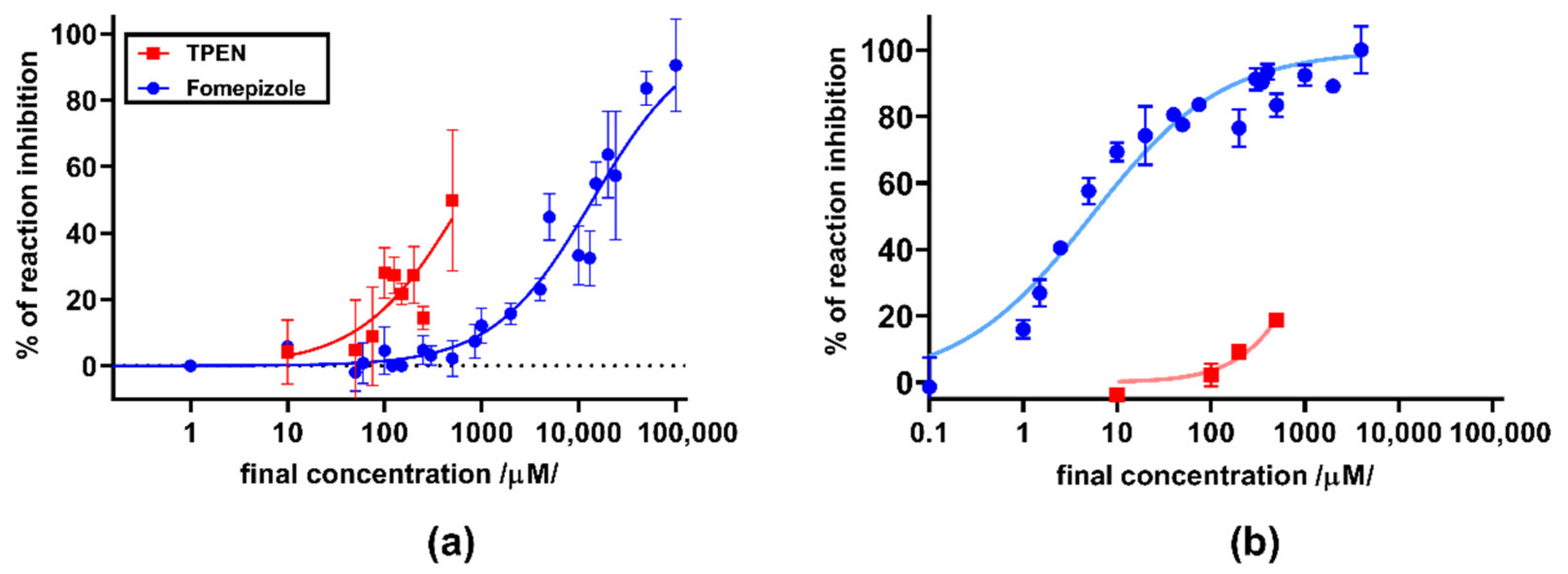
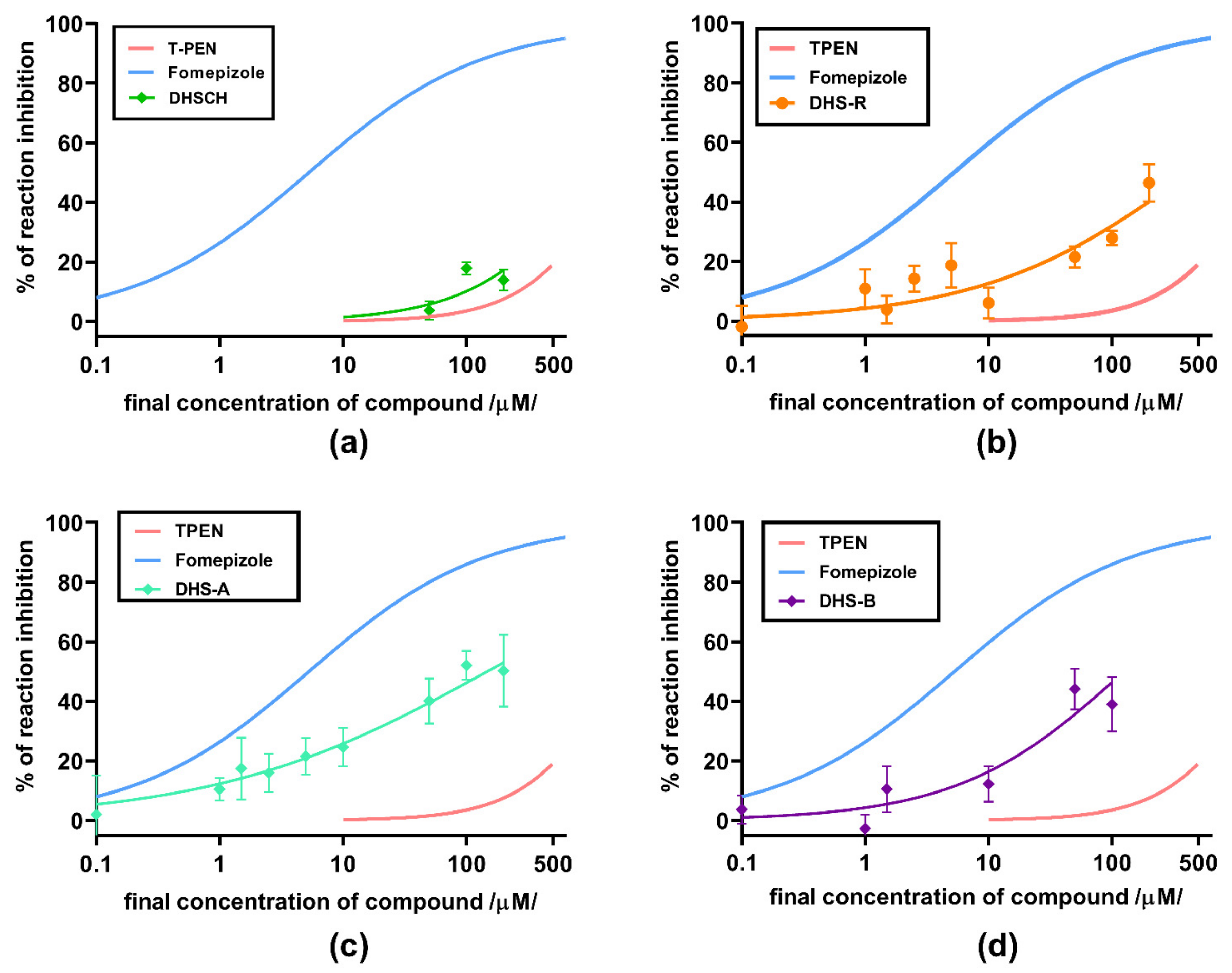
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chambers, C.S.; Holečková, V.; Petrásková, L.; Biedermann, D.; Valentová, K.; Buchta, M.; Křen, V. The silymarin composition… and why does it matter? Food Res. Int. 2017, 100, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Filippopoulou, K.; Papaevgeniou, N.; Lefaki, M.; Paraskevopoulou, A.; Biedermann, D.; Křen, V.; Chondrogianni, N. 2,3-Dehydrosilybin A/B as a pro-longevity and anti-aggregation compound. Free Radic. Biol. Med. 2017, 103, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Tilley, C.; Deep, G.; Agarwal, C.; Wempe, M.F.; Biedermann, D.; Valentová, K.; Křen, V.; Agarwal, R. Silibinin and its 2,3-dehydro-derivative inhibit basal cell carcinoma growth via suppression of mitogenic signaling and transcription factors activation. Mol. Carcinog. 2016, 55, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Y.J.; Zhou, Z.Y.; Zhang, G.L.; Lu, X.X. Metal coordination protocol for the synthesis of-2,3-dehydrosilybin and 19-O-demethyl-2,3-dehydrosilybin from silybin and their antitumor activities. Tetrahedron Lett. 2018, 59, 1666–1669. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.R.; Sekhon, A.; Oka, Y.; Chen, G.L.; Alrubati, N.; Kaur, J.; Orozco, A.; Zhang, Q.; Wang, G.D.; Chen, Q.H. 23-O-Substituted-2,3-dehydrosilybins selectively suppress androgen receptor-positive LNCaP prostate cancer cell proliferation. Nat. Prod. Commun. 2020, 15, 1–10. [Google Scholar] [CrossRef]
- Gillessen, A.; Schmidt, H.H. Silymarin as supportive treatment in liver diseases: A narrative review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef] [Green Version]
- Ferenci, P.; Dragosics, B.; Dittrich, H.; Frank, H.; Benda, L.; Lochs, H.; Meryn, S.; Base, W.; Schneider, B. Randomized controlled trial of silymarin treatment in patients with cirrhosis of the liver. J. Hepatol. 1989, 9, 105–113. [Google Scholar] [CrossRef]
- Fehér, J.; Deák, G.; Müzes, G.; Láng, I.; Niederland, V.; Nékám, K.; Kárteszi, M. Liver-protective action of silymarin therapy in chronic alcoholic liver diseases. Orv. Hetil. 1989, 130, 2723–2727. [Google Scholar]
- Müzes, G.; Deák, G.; Láng, I.; Nékám, K.; Niederland, V.; Fehér, J. Effect of silimarin (Legalon) therapy on the antioxidant defense mechanism and lipid peroxidation in alcoholic liver disease (double blind protocol). Orv. Hetil. 1990, 131, 863–866. [Google Scholar]
- Crabb, D.W.; Matsumoto, M.; Chang, D.; You, M. Overview of the role of alcohol dehydrogenase and aldehyde dehydrogenase and their variants in the genesis of alcohol-related pathology. Proc. Nutr. Soc. 2004, 63, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Crabb, D.W.; Liangpunsakul, S. Acetaldehyde generating enzyme systems: Roles of alcohol dehydrogenase, CYP2E1 and catalase, and speculations on the role of other enzymes and processes. Novartis Found. Symp. 2007, 285, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, T.; Kusumanchi, P.; Han, S.; Yang, Z.; Liangpunsakul, S. Alcohol metabolizing enzymes, microsomal ethanol oxidizing system, cytochrome P450 2E1, catalase, and aldehyde dehydrogenase in alcohol-associated liver disease. Biomedicines 2020, 8, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagi, J.H.; Vallee, B.L. The role of zinc in alcohol dehydrogenase. V. The effect of metal-binding agents on the structure of the yeast alcohol dehydrogenase molecule. J. Biol. Chem. 1960, 235, 3188–3192. [Google Scholar]
- Yang, Y.; Zhou, H.M. Effect of zinc ions on conformational stability of yeast alcohol dehydrogenase. Biochemistry 2001, 66, 47–54. [Google Scholar] [CrossRef]
- Tvrdý, V.; Catapano, M.C.; Rawlik, T.; Karlíčková, J.; Biedermann, D.; Křen, V.; Mladěnka, P.; Valentová, K. Interaction of isolated silymarin flavonolignans with iron and copper. J. Inorg. Biochem. 2018, 189, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Jörnvall, H. Differences between alcohol dehydrogenases. Structural properties and evolutionary aspects. Eur. J. Biochem. 1977, 72, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Vonwartburg, J.P.; Bethune, J.L.; Vallee, B.L. Human liver--alcohol dehydrogenase. Kinetic and physicochemical properties. Biochemistry 1964, 3, 1775–1782. [Google Scholar] [CrossRef]
- Pietruszko, R.; Crawford, K.; Lester, D. Comparison of substrate specificity of alcohol dehydrogenases from human liver, horse liver, and yeast towards saturated and 2-enoic alcohols and aldehydes. Arch. Biochem. Biophys. 1973, 159, 50–60. [Google Scholar] [CrossRef]
- Petrásková, L.; Káňová, K.; Biedermann, D.; Křen, V.; Valentová, K. Simple and rapid HPLC separation and quantification of flavonoid, flavonolignans, and 2,3-dehydroflavonolignans in silymarin. Foods 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gažák, R.; Marhol, P.; Purchartová, K.; Monti, D.; Biedermann, D.; Riva, S.; Cvak, L.; Křen, V. Large-scale separation of silybin diastereoisomers using lipases. Process Biochem. 2010, 45, 1657–1663. [Google Scholar] [CrossRef]
- Gažák, R.; Trouillas, P.; Biedermann, D.; Fuksová, K.; Marhol, P.; Kuzma, M.; Křen, V. Base-catalyzed oxidation of silybin and isosilybin into 2,3-dehydro derivatives. Tetrahedron Lett. 2013, 54, 315–317. [Google Scholar] [CrossRef]
- Biedermann, D.; Buchta, M.; Holečková, V.; Sedlák, D.; Valentová, K.; Cvačka, J.; Bednárová, L.; Křenková, A.; Kuzma, M.; Škuta, C.; et al. Silychristin: Skeletal alterations and biological activities. J. Nat. Prod. 2016, 79, 3086–3092. [Google Scholar] [CrossRef] [PubMed]
- Valentová, K.; Purchartová, K.; Rydlová, L.; Roubalová, L.; Biedermann, D.; Petrásková, L.; Křenková, A.; Pelantová, H.; Holečková-Moravcová, V.; Tesařová, E.; et al. Sulfated metabolites of flavonolignans and 2,3-dehydroflavonolignans: Preparation and properties. Int. J. Mol. Sci. 2018, 19, 2349. [Google Scholar] [CrossRef] [Green Version]
- Catapano, M.C.; Tvrdý, V.; Karlíčková, J.; Mercolini, L.; Mladěnka, P. A simple, cheap but reliable method for evaluation of zinc chelating properties. Bioorg. Chem. 2018, 77, 287–292. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Lai, L.; Wang, S. Further development and validation of empirical scoring functions for structure-based binding affinity prediction. J. Comput. Aided Mol. Des. 2002, 16, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, L.D. Pymol. Available online: http://www.pymol.org/pymol (accessed on 20 August 2021).
- Bailey, J.; Powell, L.; Sinanan, L.; Neal, J.; Li, M.; Smith, T.; Bell, E. A novel mechanism of V-type zinc inhibition of glutamate dehydrogenase results from disruption of subunit interactions necessary for efficient catalysis. FEBS J. 2011, 278, 3140–3151. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.; Costa, V.M.; Carvalho, A.; Baptista, P.; de Pinho, P.G.; de Lourdes Bastos, M.; Carvalho, F. Amanita phalloides poisoning: Mechanisms of toxicity and treatment. Food Chem. Toxicol. 2015, 86, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Saller, R.; Brignoli, R.; Melzer, J.; Meier, R. An updated systematic review with meta-analysis for the clinical evidence of silymarin. Forsch Komplementmed. 2008, 15, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Mendoza, N.; Madrigal-Santillán, E.; Morales-González, A.; Esquivel-Soto, J.; Esquivel-Chirino, C.; García-Luna, Y.G.-R.M.; Gayosso-de-Lucio, J.A.; Morales-González, J.A. Hepatoprotective effect of silymarin. World J. Hepatol. 2014, 6, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Tuma, D.J.; Thiele, G.M.; Xu, D.; Klassen, L.W.; Sorrell, M.F. Acetaldehyde and malondialdehyde react together to generate distinct protein adducts in the liver during long-term ethanol administration. Hepatology 1996, 23, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Crabb, D.W.; Galli, A.; Fischer, M.; You, M. Molecular mechanisms of alcoholic fatty liver: Role of peroxisome proliferator-activated receptor alpha. Alcohol 2004, 34, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, A.; Bustamante, J.C.; Videla, C.; Guerra, R. Effect of silybin dihemisuccinate on the ethanol metabolizing systems of the rat liver. Cell Biochem. Funct. 1989, 7, 173–178. [Google Scholar] [CrossRef]
- Hsieh, Y.L.; Yeh, Y.H.; Lee, Y.T.; Huang, C.Y. Protective effects of cholestin on ethanol induced oxidative stress in rats. J. Sci. Food Agric. 2015, 95, 799–808. [Google Scholar] [CrossRef]
- Nosova, T.; Jousimies-Somer, H.; Kaihovaara, P.; Jokelainen, K.; Heine, R.; Salaspuro, M. Characteristics of alcohol dehydrogenases of certain aerobic bacteria representing human colonic flora. Alcohol Clin. Exp. Res. 1997, 21, 489–494. [Google Scholar] [CrossRef]
- Theorell, H.; Yonetani, T.; Sjöberg, B. On the effects of some heterocyclic compounds on the enzymic activity of liver alcohol dehydrogenase. Acta Chem. Scand. 1969, 23, 255–260. [Google Scholar] [CrossRef]
- Bosron, W.F.; Li, T.K. Catalytic properties of human liver alcohol dehydrogenase isoenzymes. Enzyme 1987, 37, 19–28. [Google Scholar] [CrossRef]
- Pietruszko, R. Human liver alcohol dehydrogenase—Inhibition of methanol activity by pyrazole, 4-methylpyrazole, 4-hydroxymethylpyrazole and 4-carboxypyrazole. Biochem. Pharmacol. 1975, 24, 1603–1607. [Google Scholar] [CrossRef]
- Li, T.K.; Theorell, H. Human liver alcohol dehydrogenase—Inhibition by pyrazole and pyrazole analogs. Acta Chem. Scand. 1969, 23, 892–902. [Google Scholar] [CrossRef]
- Theorell, H.; Yonetani, T. Liver alcohol dehydrogenase-DNP-pyrazole complex: A model of a ternary intermediate in the enzyme reaction. Biochem. Z. 1963, 338, 537–553. [Google Scholar]
- Xie, P.T.; Hurley, T.D. Methionine-141 directly influences the binding of 4-methylpyrazole in human sigma sigma alcohol dehydrogenase. Protein Sci. 1999, 8, 2639–2644. [Google Scholar] [CrossRef] [PubMed]
- Jörnvall, H.; Eklund, H.; Brändén, C.I. Subunit conformation of yeast alcohol dehydrogenase. J. Biol. Chem. 1978, 253, 8414–8419. [Google Scholar] [CrossRef]
- Jörnvall, H.; Hempel, J.; von Bahr-Lindström, H.; Höög, J.O.; Vallee, B.L. Alcohol and aldehyde dehydrogenases: Structures of the human liver enzymes, functional properties and evolutionary aspects. Alcohol Alcohol. Suppl. 1987, 1, 13–23. [Google Scholar]
- Meeusen, J.W.; Nowakowski, A.; Petering, D.H. Reaction of metal-binding ligands with the zinc proteome: Zinc sensors and N,N,N’,N’-tetrakis(2-pyridylmethyl)ethylenediamine. Inorg. Chem. 2012, 51, 3625–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vimalraj, S.; Rajalakshmi, S.; Saravanan, S.; Raj Preeth, D.; Vasanthi, R.L.A.; Shairam, M.; Chatterjee, S. Synthesis and characterization of zinc-silibinin complexes: A potential bioactive compound with angiogenic, and antibacterial activity for bone tissue engineering. Colloids Surf. B Biointerfaces 2018, 167, 134–143. [Google Scholar] [CrossRef]
- Lapouge, C.; Dangleterre, L.; Cornard, J.P. Spectroscopic and theoretical studies of the Zn(II) chelation with hydroxyflavones. J. Phys. Chem. A 2006, 110, 12494–12500. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.F.; De Giovani, W.F. Synthesis, spectral and electrochemical properties of Al(III) and Zn(II) complexes with flavonoids. Spectrochim Acta A Mol. Biomol. Spectrosc. 2005, 61, 1985–1990. [Google Scholar] [CrossRef]
- Říha, M.; Karlíčková, J.; Filipský, T.; Macáková, K.; Rocha, L.; Bovicelli, P.; Silvestri, I.P.; Saso, L.; Jahodář, L.; Hrdina, R.; et al. In vitro evaluation of copper-chelating properties of flavonoids. RSC Adv. 2014, 4, 32628–32638. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, C.H.; Allen, A.; Stanley, C.A.; Smith, T.J. The structure and allosteric regulation of mammalian glutamate dehydrogenase. Arch. Biochem. Biophys. 2012, 519, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Allen, A.; Smith, T.J. High throughput screening reveals several new classes of glutamate dehydrogenase inhibitors. Biochemistry 2007, 46, 15089–15102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calani, L.; Brighenti, F.; Bruni, R.; Del Rio, D. Absorption and metabolism of milk thistle flavanolignans in humans. Phytomedicine 2012, 20, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Tong, S.S.; Xu, Y.; Wang, L.; Fu, M.; Ge, Y.R.; Yu, J.N.; Xu, X.M. Proliposomes for oral delivery of dehydrosilymarin: Preparation and evaluation in vitro and in vivo. Acta Pharmacol. Sin. 2011, 32, 973–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]





















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tvrdý, V.; Hrubša, M.; Jirkovský, E.; Biedermann, D.; Kutý, M.; Valentová, K.; Křen, V.; Mladěnka, P. Silymarin Dehydroflavonolignans Chelate Zinc and Partially Inhibit Alcohol Dehydrogenase. Nutrients 2021, 13, 4238. https://doi.org/10.3390/nu13124238
Tvrdý V, Hrubša M, Jirkovský E, Biedermann D, Kutý M, Valentová K, Křen V, Mladěnka P. Silymarin Dehydroflavonolignans Chelate Zinc and Partially Inhibit Alcohol Dehydrogenase. Nutrients. 2021; 13(12):4238. https://doi.org/10.3390/nu13124238
Chicago/Turabian StyleTvrdý, Václav, Marcel Hrubša, Eduard Jirkovský, David Biedermann, Michal Kutý, Kateřina Valentová, Vladimír Křen, and Přemysl Mladěnka. 2021. "Silymarin Dehydroflavonolignans Chelate Zinc and Partially Inhibit Alcohol Dehydrogenase" Nutrients 13, no. 12: 4238. https://doi.org/10.3390/nu13124238
APA StyleTvrdý, V., Hrubša, M., Jirkovský, E., Biedermann, D., Kutý, M., Valentová, K., Křen, V., & Mladěnka, P. (2021). Silymarin Dehydroflavonolignans Chelate Zinc and Partially Inhibit Alcohol Dehydrogenase. Nutrients, 13(12), 4238. https://doi.org/10.3390/nu13124238