
Higher-Dose DHA Supplementation Modulates Immune Responses in Pregnancy and Is Associated with Decreased Preterm Birth

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
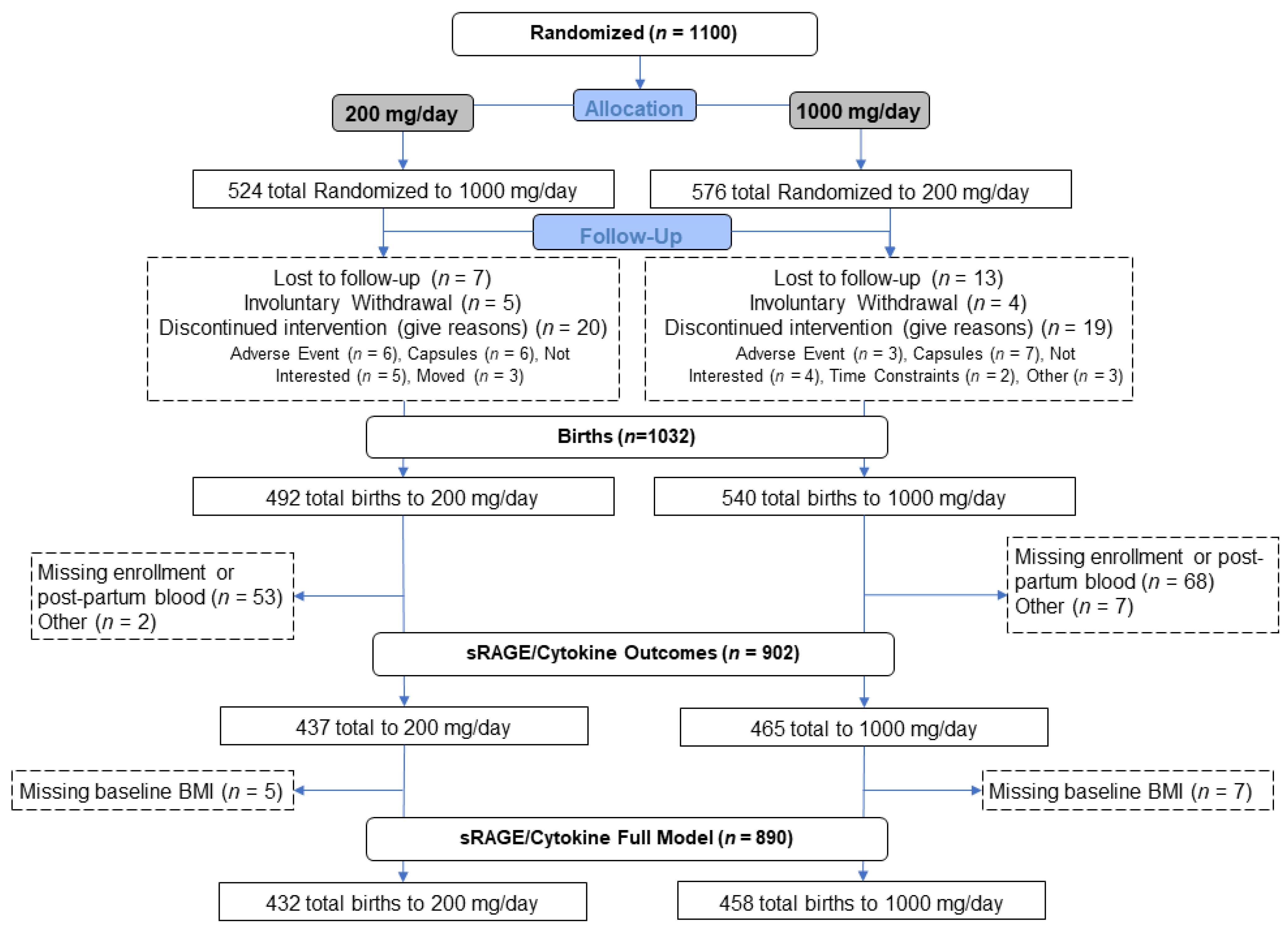
2.1. Study Design
2.2. Blood Collection
2.3. ELISA
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prairie, E.; Cote, F.; Tsakpinoglou, M.; Mina, M.; Quiniou, C.; Leimert, K.; Olson, D.; Chemtob, S. The determinant role of IL-6 in the establishment of inflammation leading to spontaneous preterm birth. Cytokine Growth Factor Rev. 2021, 59, 118–130. [Google Scholar] [CrossRef]
- Romero, R.; Espinoza, J.; Kusanovic, J.P.; Gotsch, F.; Hassan, S.; Erez, O.; Chaiworapongsa, T.; Mazor, M. The preterm parturition syndrome. BJOG 2006, 113 (Suppl. 3), 17–42. [Google Scholar] [CrossRef]
- Calder, P.C. Immunomodulation by omega-3 fatty acids. Prostaglandins Leukot Essent Fat. Acids 2007, 77, 327–335. [Google Scholar] [CrossRef]
- Middleton, P.; Gomersall, J.C.; Gould, J.F.; Shepherd, E.; Olsen, S.F.; Makrides, M. Omega-3 fatty acid addition during pregnancy. Cochrane Database Syst. Rev. 2018, 11, CD003402. [Google Scholar] [CrossRef]
- Valentine, C.J.; Dingess, K.A.; Kleiman, J.; Morrow, A.L.; Rogers, L.K. A Randomized Trial of Maternal Docosahexaenoic Acid Supplementation to Reduce Inflammation in Extremely Preterm Infants. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 388–392. [Google Scholar] [CrossRef]
- Yavin, E. Versatile roles of docosahexaenoic acid in the prenatal brain: From pro- and anti-oxidant features to regulation of gene expression. Prostaglandins Leukot Essent Fat. Acids 2006, 75, 203–211. [Google Scholar] [CrossRef]
- Calder, P.C. The relationship between the fatty acid composition of immune cells and their function. Prostaglandins Leukot Essent Fat. Acids 2008, 79, 101–108. [Google Scholar] [CrossRef]
- Himmelfarb, J.; Phinney, S.; Ikizler, T.A.; Kane, J.; McMonagle, E.; Miller, G. Gamma-tocopherol and docosahexaenoic acid decrease inflammation in dialysis patients. J. Ren. Nutr. 2007, 17, 296–304. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef]
- McCann, J.C.; Ames, B.N. Is docosahexaenoic acid, an n-3 long-chain polyunsaturated fatty acid, required for development of normal brain function? An overview of evidence from cognitive and behavioral tests in humans and animals. Am. J. Clin. Nutr. 2005, 82, 281–295. [Google Scholar] [CrossRef]
- Rizza, S.; Tesauro, M.; Cardillo, C.; Galli, A.; Iantorno, M.; Gigli, F.; Sbraccia, P.; Federici, M.; Quon, M.J.; Lauro, D. Fish oil supplementation improves endothelial function in normoglycemic offspring of patients with type 2 diabetes. Atherosclerosis 2009, 206, 569–574. [Google Scholar] [CrossRef]
- Calder, P.C. The role of marine omega-3 (n-3) fatty acids in inflammatory processes, atherosclerosis and plaque stability. Mol. Nutr. Food Res. 2012, 56, 1073–1080. [Google Scholar] [CrossRef]
- Rogers, L.K.; Graf, A.E.; Bhatia, A.; Leonhart, K.L.; Oza-Frank, R. Associations between maternal and infant morbidities and sRAGE within the first week of life in extremely preterm infants. PLoS ONE 2013, 8, e82537. [Google Scholar] [CrossRef]
- Rogers, L.K.; Valentine, C.J.; Keim, S.A. DHA supplementation: Current implications in pregnancy and childhood. Pharmacol. Res. 2013, 70, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Matesanz, N.; Park, G.; McAllister, H.; Leahey, W.; Devine, A.; McVeigh, G.E.; Gardiner, T.A.; McDonald, D.M. Docosahexaenoic acid improves the nitroso-redox balance and reduces VEGF-mediated angiogenic signaling in microvascular endothelial cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6815–6825. [Google Scholar] [CrossRef] [Green Version]
- Kar, S.; Wong, M.; Rogozinska, E.; Thangaratinam, S. Effects of omega-3 fatty acids in prevention of early preterm delivery: A systematic review and meta-analysis of randomized studies. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 198, 40–46. [Google Scholar] [CrossRef]
- Carlson, S.E.; Colombo, J.; Gajewski, B.J.; Gustafson, K.M.; Mundy, D.; Yeast, J.; Georgieff, M.K.; Markley, L.A.; Kerling, E.H.; Shaddy, D.J. DHA supplementation and pregnancy outcomes. Am. J. Clin. Nutr. 2013, 97, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Koletzko, B.; Cetin, I.; Brenna, J.T. Dietary fat intakes for pregnant and lactating women. Br. J. Nutr. 2007, 98, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Carlson, S.E.; Gajewski, B.J.; Valentine, C.J.; Kerling, E.H.; Weiner, C.P.; Cackovic, M.; Buhimschi, C.S.; Rogers, L.K.; Sands, S.A.; Brown, A.R.; et al. Higher dose docosahexaenoic acid supplementation during pregnancy and early preterm birth: A randomised, double-blind, adaptive-design superiority trial. EClinicalMedicine 2021, 36, 100905. [Google Scholar] [CrossRef]
- Kalagiri, R.R.; Carder, T.; Choudhury, S.; Vora, N.; Ballard, A.R.; Govande, V.; Drever, N.; Beeram, M.R.; Uddin, M.N. Inflammation in Complicated Pregnancy and Its Outcome. Am. J. Perinatol. 2016, 33, 1337–1356. [Google Scholar] [CrossRef]
- Germanova, A.; Koucky, M.; Hajek, Z.; Parizek, A.; Zima, T.; Kalousova, M. Soluble receptor for advanced glycation end products in physiological and pathological pregnancy. Clin. Biochem. 2010, 43, 442–446. [Google Scholar] [CrossRef]
- Chau, A.; Markley, J.C.; Juang, J.; Tsen, L.C. Cytokines in the perinatal period—Part II. Int. J. Obstet. Anesth. 2016, 26, 48–58. [Google Scholar] [CrossRef]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P.M. Maternal Immunological Adaptation During Normal Pregnancy. Front. Immunol. 2020, 11, 575197. [Google Scholar] [CrossRef]
- Allam-Ndoul, B.; Guenard, F.; Barbier, O.; Vohl, M.C. Effect of n-3 fatty acids on the expression of inflammatory genes in THP-1 macrophages. Lipids Health Dis. 2016, 15, 69. [Google Scholar] [CrossRef] [Green Version]
- Chavakis, T.; Bierhaus, A.; Al-Fakhri, N.; Schneider, D.; Witte, S.; Linn, T.; Nagashima, M.; Morser, J.; Arnold, B.; Preissner, K.T.; et al. The pattern recognition receptor (RAGE) is a counterreceptor for leukocyte integrins: A novel pathway for inflammatory cell recruitment. J. Exp. Med. 2003, 198, 1507–1515. [Google Scholar] [CrossRef]
- Buckley, S.T.; Ehrhardt, C. The receptor for advanced glycation end products (RAGE) and the lung. J. Biomed. Biotechnol. 2010, 2010, 917108. [Google Scholar] [CrossRef] [Green Version]
- Morbini, P.; Villa, C.; Campo, I.; Zorzetto, M.; Inghilleri, S.; Luisetti, M. The receptor for advanced glycation end products and its ligands: A new inflammatory pathway in lung disease? Mod. Pathol. 2006, 19, 1437–1445. [Google Scholar] [CrossRef] [Green Version]
- Schlueter, C.; Hauke, S.; Flohr, A.M.; Rogalla, P.; Bullerdiek, J. Tissue-specific expression patterns of the RAGE receptor and its soluble forms--a result of regulated alternative splicing? Biochim. Biophys. Acta 2003, 1630, 1–6. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Piper, M.G.; McMaken, S.; Mo, X.; Opalek, J.; Schmidt, A.M.; Marsh, C.B. sRAGE induces human monocyte survival and differentiation. J. Immunol. 2010, 185, 1822–1835. [Google Scholar] [CrossRef] [Green Version]
- Raucci, A.; Cugusi, S.; Antonelli, A.; Barabino, S.M.; Monti, L.; Bierhaus, A.; Reiss, K.; Saftig, P.; Bianchi, M.E. A soluble form of the receptor for advanced glycation endproducts (RAGE) is produced by proteolytic cleavage of the membrane-bound form by the sheddase a disintegrin and metalloprotease 10 (ADAM10). FASEB J. 2008, 22, 3716–3727. [Google Scholar] [CrossRef]
- Oliver, E.A.; Buhimschi, C.S.; Dulay, A.T.; Baumbusch, M.A.; Abdel-Razeq, S.S.; Lee, S.Y.; Zhao, G.; Jing, S.; Pettker, C.M.; Buhimschi, I.A. Activation of the receptor for advanced glycation end products system in women with severe preeclampsia. J. Clin. Endocrinol. Metab. 2011, 96, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Hajek, Z.; Germanova, A.; Koucky, M.; Zima, T.; Kopecky, P.; Vitkova, M.; Parizek, A.; Kalousova, M. Detection of feto-maternal infection/inflammation by the soluble receptor for advanced glycation end products (sRAGE): Results of a pilot study. J. Perinat. Med. 2008, 36, 399–404. [Google Scholar] [CrossRef]
- Kwon, J.H.; Kim, Y.H.; Kwon, J.Y.; Park, Y.W. Clinical significance of serum sRAGE and esRAGE in women with normal pregnancy and preeclampsia. J. Perinat. Med. 2011, 39, 507–513. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Y.; Yue, C.; Ye, Y.; Chen, P.; Peng, W.; Wang, Y. Accumulation of Advanced Glycation End Products Involved in Inflammation and Contributing to Severe Preeclampsia, in Maternal Blood, Umbilical Blood and Placental Tissues. Gynecol. Obstet. Investig. 2017, 82, 388–397. [Google Scholar] [CrossRef]
- Quintanilla-Garcia, C.V.; Uribarri, J.; Fajardo-Araujo, M.E.; Barrientos-Romero, J.J.; Romero-Gutierrez, G.; Reynaga-Ornelas, M.G.; Garay-Sevilla, M.E. Changes in circulating levels of carboxymethyllysine, soluble receptor for advanced glycation end products (sRAGE), and inflammation markers in women during normal pregnancy. J. Matern. Fet. Neonat. Med. 2019, 32, 4102–4107. [Google Scholar] [CrossRef]
- Rzepka, R.; Dolegowska, B.; Rajewska, A.; Salata, D.; Budkowska, M.; Kwiatkowski, S.; Torbe, A. Diagnostic Potential of Evaluation of SDF-1alpha and sRAGE Levels in Threatened Premature Labor. Biomed. Res. Int. 2016, 2016, 2719460. [Google Scholar] [CrossRef] [Green Version]
- Rzepka, R.; Dolegowska, B.; Rajewska, A.; Kwiatkowski, S.; Salata, D.; Budkowska, M.; Domanski, L.; Mikolajek-Bedner, W.; Torbe, A. Soluble and Endogenous Secretory Receptors for Advanced Glycation End Products in Threatened Preterm Labor and Preterm Premature Rupture of Fetal Membranes. Biomed. Res. Int. 2015, 2015, 568042. [Google Scholar] [CrossRef] [Green Version]
- Bastek, J.A.; Brown, A.G.; Foreman, M.N.; McShea, M.A.; Anglim, L.M.; Adamczak, J.E.; Elovitz, M.A. The soluble receptor for advanced glycation end products can prospectively identify patients at greatest risk for preterm birth. J. Matern. Fet. Neonat. Med. 2012, 25, 1762–1768. [Google Scholar] [CrossRef]
- Leimert, K.B.; Xu, W.; Princ, M.M.; Chemtob, S.; Olson, D.M. Inflammatory Amplification: A Central Tenet of Uterine Transition for Labor. Front. Cell. Infect. Microbiol. 2021, 11, 660983. [Google Scholar] [CrossRef]
- Ren, J.; Qiang, Z.; Li, Y.Y.; Zhang, J.N. Biomarkers for a histological chorioamnionitis diagnosis in pregnant women with or without group B streptococcus infection: A case-control study. BMC Pregnancy Childbirth 2021, 21, 250. [Google Scholar] [CrossRef]
- Li, W.; Zhao, X.; Li, S.; Chen, X.; Cui, H.; Chang, Y.; Zhang, R. Upregulation of TNF-alpha and IL-6 induces preterm premature rupture of membranes by activation of ADAMTS-9 in embryonic membrane cells. Life Sci. 2020, 260, 118237. [Google Scholar] [CrossRef]
- Carlson, S.E.; Gajewski, B.J.; Valentine, C.J.; Rogers, L.K.; Weiner, C.P.; DeFranco, E.A.; Buhimschi, C.S. Assessment of DHA on reducing early preterm birth: The ADORE randomized controlled trial protocol. BMC Pregnancy Childbirth 2017, 17, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| 200 mg/day n = 437 (47.6%) | 1000 mg/day n = 465 (52.4%) | Total n = 902 | |
|---|---|---|---|
| sRAGE [pg/mL], mean (SD) | |||
| Enrollment | 564.4 (286.4) | 520.6 (279.4) | 541.8 (283.5) |
| Delivery | 467.7 (277.3) | 443.1 (381.4) | 455.0 (335.1) |
| IL 6 [pg/mL], mean (SD) | |||
| Enrollment | 0.9 (1.0) | 1.3 (6.1) | 1.1 (4.4) |
| Delivery | 4.5 (7.5) | 7.7 (26.0) | 6.1 (19.4) |
| IL 1β [pg/mL], mean (SD) | |||
| Enrollment | 0.1 (0.2) | 0.1 (0.2) | 0.1 (0.2) |
| Delivery | 0.2 (0.8) | 0.2 (1.0) | 0.2 (0.9) |
| TNFα [pg/mL], mean (SD) | |||
| Enrollment | 1.7 (0.7) | 1.9 (2.1) | 1.8 (1.6) |
| Delivery | 2.1 (1.0) | 2.1 (1.0) | 2.1 (1.0) |
| INFγ [pg/mL], mean (SD) | |||
| Enrollment | 6.4 (30.8) | 5.1 (13.5) | 5.7 (23.5) |
| Delivery | 5.1 (22.7) | 4.4 (8.9) | 4.7 (17.0) |
| DHA in RBC fatty acids [%], mean (SD) | 6.5 (1.8) | 6.4 (1.8) | 6.5 (1.8) |
| Smoker (before or during pregnancy), yes n (%) | 105 (24.0) | 112 (24.1) | 217 (24.1) |
| Race, n (%) | |||
| Non-Hispanic Black | 104 (23.8) | 89 (19.1) | 193 (21.4) |
| Other | 333 (76.2) | 376 (80.9) | 709 (78.6) |
| History of Preeclampsia, yes n (%) | 33 (7.6) | 30 (6.5) | 63 (7.0) |
| BMI Group, n (%) | n = 432 | n = 458 | n = 890 |
| Obese | 138 (31.9) | 159 (34.7) | 297 (33.4) |
| Other (BMI < 30) | 294 (68.1) | 299 (65.3) | 593 (66.6) |
| Posterior Mean (95% Bayesian Credible Interval) | Bayesian Posterior Probability | |
|---|---|---|
| Gestational age at birth | ||
| Treatment 1000 vs. 200 mg per day | 0.23 (0.03, 0.42) | 0.99 |
| Enrollment sRAGE [pg/mL] | 0.0006 (0.0002, 0.0011) | 0.996 |
| Delivery sRAGE [pg/mL] | −0.0008 (−0.0012, −0.0004) | 0.00 |
| Significant variables | ||
| Maternal Race (non-Hispanic Black vs. other) | −0.44 (−0.70, −0.19) | 0.0003 |
| Pre-pregnancy BMI (obese vs. other) | −0.50 (−0.73, −0.27) | 0.00 |
| History of preeclampsia (yes vs. no) | −1.17 (−1.56, −0.78) | 0.00 |
| DHA at Enrollment [%] | 0.08 (0.02, 0.13) | 0.996 |
| Smoker (before or during pregnancy, yes vs. no) | −0.21 (−0.44, 0.03) | 0.04 |
| *,# Preterm birth (<37 weeks) | ||
| Treatment, 1000 vs. 200 mg per day | 0.59 (0.33, 0.96) | 0.98 |
| Enrollment sRAGE [pg/mL] | 0.999 (0.988, 1.0) | 0.92 |
| Delivery sRAGE [pg/mL] | 1.001 (1.0, 1.002) | 0.002 |
| Significant variables | ||
| Maternal Race (non-Hispanic Black vs. other) | 1.77 (0.95, 3.03) | 0.04 |
| Pre-pregnancy BMI (obese vs. other) | 1.60 (0.88, 2.71) | 0.07 |
| History of preeclampsia (yes vs. no) | 4.15 (1.91, 7.64) | 0.0003 |
| DHA at Enrollment [%] | 0.84 (0.70, 0.99) | 0.98 |
| Smoker (before or during pregnancy, yes vs. no) | 1.40 (0.75, 2.34) | 0.16 |
| Posterior Mean (95% Bayesian Credible Interval) | Bayesian Posterior Probability | |
|---|---|---|
| Gestational age at birth | ||
| Enrollment IL-6 [pg/mL] | 0.003 (−0.022, 0.028) | 0.58 |
| Delivery IL-6 [pg/mL] | −0.0005 (−0.0063, 0.0055) | 0.44 |
| Enrollment IL-1β [pg/mL] | −0.18 (−0.61, 0.25) | 0.20 |
| Delivery IL-1β [pg/mL] | −0.03 (−0.14, 0.07) | 0.26 |
| Enrollment TNFα [pg/mL] | 0.02 (−0.04, 0.09) | 0.76 |
| Delivery TNFα [pg/mL] | −0.08 (−0.19, 0.02) | 0.07 |
| Enrollment INFγ [pg/mL] | −0.001 (−0.005, 0.003) | 0.27 |
| Delivery INFγ [pg/mL] | −0.005 (−0.010, 0.001) | 0.06 |
| * Preterm birth (<37 weeks) | ||
| Enrollment IL-6 [pg/mL] | 0.89 (0.67, 1.04) | 0.87 |
| Delivery IL-6 [pg/mL] | 0.99 (0.95, 1.01) | 0.85 |
| Enrollment IL-1β [pg/mL] | 1.53 (0.56, 3.00) | 0.20 |
| Delivery IL-1β [pg/mL] | 0.91 (0.60, 1.16) | 0.73 |
| Enrollment TNFα [pg/mL] | 0.73 (0.45, 1.04) | 0.95 |
| Delivery TNFα [pg/mL] | 1.33 (1.00, 1.74) | 0.03 |
| Enrollment INFγ [pg/mL] | 0.99 (0.95, 1.01) | 0.83 |
| Delivery INFγ [pg/mL] | 0.996 (0.972, 1.010) | 0.62 |
| Observed Difference (Delivery-Enrollment) | Posterior Mean (95% Bayesian Credible Interval) | Bayesian Posterior Probability (1000 mg Is Greater than 200 mg) | |||
|---|---|---|---|---|---|
| 200 mg | 1000 mg | 200 mg | 1000 mg | ||
| sRAGE [pg/mL] | −96.76 (244.43) | −77.49 (324.63) | −96.94 (−663.9, 472.4) | −77.55 (−651.5, 496.7) | 0.84 |
| IL-6 [pg/mL] | 3.66 (7.33) | 6.36 (23.91) | 3.65 (−31.60, 39.08) | 6.36 (−29.18, 42.01) | 0.99 |
| IL-1β [pg/mL] | 0.15 (0.84) | 0.18 (1.05) | 0.15 (−1.73, 2.03) | 0.18 (−1.72, 2.08) | 0.70 |
| TNFα [pg/mL] | 0.39 (0.71) | 0.28 (2.14) | 0.39 (−2.78, 3.57) | 0.28 (−2.93, 3.5) | 0.16 |
| INFγ [pg/mL] | −1.32 (38.29) | −0.66 (16.19) | −1.34 (−58.62, 56.53) | −0.67 (−58.37, 56.98) | 0.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valentine, C.J.; Khan, A.Q.; Brown, A.R.; Sands, S.A.; Defranco, E.A.; Gajewski, B.J.; Carlson, S.E.; Reber, K.M.; Rogers, L.K. Higher-Dose DHA Supplementation Modulates Immune Responses in Pregnancy and Is Associated with Decreased Preterm Birth. Nutrients 2021, 13, 4248. https://doi.org/10.3390/nu13124248
Valentine CJ, Khan AQ, Brown AR, Sands SA, Defranco EA, Gajewski BJ, Carlson SE, Reber KM, Rogers LK. Higher-Dose DHA Supplementation Modulates Immune Responses in Pregnancy and Is Associated with Decreased Preterm Birth. Nutrients. 2021; 13(12):4248. https://doi.org/10.3390/nu13124248
Chicago/Turabian StyleValentine, Christina J., Aiman Q. Khan, Alexandra R. Brown, Scott A. Sands, Emily A. Defranco, Byron J. Gajewski, Susan E. Carlson, Kristina M. Reber, and Lynette K. Rogers. 2021. "Higher-Dose DHA Supplementation Modulates Immune Responses in Pregnancy and Is Associated with Decreased Preterm Birth" Nutrients 13, no. 12: 4248. https://doi.org/10.3390/nu13124248
APA StyleValentine, C. J., Khan, A. Q., Brown, A. R., Sands, S. A., Defranco, E. A., Gajewski, B. J., Carlson, S. E., Reber, K. M., & Rogers, L. K. (2021). Higher-Dose DHA Supplementation Modulates Immune Responses in Pregnancy and Is Associated with Decreased Preterm Birth. Nutrients, 13(12), 4248. https://doi.org/10.3390/nu13124248