1. Introduction
Shatavari (
Asparagus racemosus Willd) has long been used as an Ayurvedic herb for women’s health [
1], but empirical evidence for its effectiveness remains limited. Steroidal saponins are thought to be the principal bioactive constituents of shatavari root. These saponins are known as shatavarins I–IV, and they are glycosides of sarsasapogenin [
2]. Shatavari root also contains other chemical constituents of note, including racemosides, racemosol, racemofuran and asparagamine A, all of which display antioxidant activity [
2]. The limited research that exists suggests that shatavari also contains phytoestrogenic compounds that are capable of binding to the estradiol (E2) receptors (E2R). Indeed, E2R functionalised magnetic nanoparticles can isolate phytoestrogens of 4.6 nmol E2-equivalent activity from a 1-mg crude extract of
Asparagus racemosus [
3]. The phytoestrogens that are most relevant to human health are those for which the following is the case: (1) they are distinct from structures that occur naturally in mammals; (2) they are contained in plants that are consumed by humans; and (3) they are bioavailable and capable of binding to human E2R [
4]. Thus, the isoflavones genistein and daidzein, the coumestan coumestrol, the lignans enterolactone and enterodiol, the flavonols quercetin, kaempferol, rutin and the stilbene resveratrol represent the most prolific and relevant dietary forms of phytoestrogens in humans [
4,
5]. In silico research suggested that shatavari-derived phytoestrogens, including rutin, kaempferol, genistein, daidzein and quercetin, bind to E2R with greater affinity than a selective estrogen receptor modulator control, bazedoxifene [
6].
Shatavari administration in rats induces pro-estrogenic changes in mammary glands and genital organs [
7]. Shatavari also appears to have galactogogue activity; in a placebo-controlled study that administered 60 mg/kg/d shatavari root powder to breastfeeding mothers for 30 days, systemic prolactin concentrations were increased (three-fold) and shatavari promoted infant weight gain [
8]. However, a similar study (in which the shatavari dose was unclear) found no effect on systemic prolactin concentrations [
9]; both studies were of poor quality. The mechanism by which shatavari increases serum prolactin concentration is unknown. Little is definitively known about how herbal galactagogues act, beyond speculation that they have E2-like activity and rely upon phytoestrogen’s interaction with estrogen receptor alpha (Erα) [
10,
11].
The potential for estrogen-like effects of shatavari supplementation has implications for both muscle and bone health after the menopause. Observational data have long suggested that in women, post-menopausal estrogenic deficits may promote the age-associated decline in muscle strength and function (sarcopenia); men experience a more gradual decline in handgrip strength (HGS) across the lifespan, whereas women display a sharp decline in such strength in the middle of the sixth decade [
12]. A large meta-analysis of studies that examined muscle strength and hormone replacement therapy (HRT) demonstrated that HRT increases muscle strength [
13]. E2 withdrawal has a detrimental effect on myosin binding function and, therefore, muscle force production; conversely, E2 improves muscle force production, and may promote muscle protein synthesis [
14,
15,
16]. One study showed that shatavari (500 mg·d
−1 root extract; fresh weight equivalent not stated) enhanced strength gains in young men following eight weeks of bench press training [
17]; this was an observational study with no indication of possible mechanisms of action. However, it does suggest that shatavari may be a beneficial ergogenic aid in supporting resistance training-induced increases in muscle size and strength; no assessment of these ergogenic effects has been made in older adults.
The decreased E2 production that precipitates menopause is also associated with a decline in bone mineral density, a phenomenon that increases the risk of bone fractures [
18]. Elderly individuals that experience a hip fracture have a three-fold increase in their risk of all-cause mortality [
19]. HRT decreases the risk of osteoporosis and fractures [
20]. Clinical trials that suggested limited therapeutic benefit of HRT and an increased risk of harm (breast cancer, cardiovascular disease) were recently reassessed and HRT was once again determined to have a clinical benefit and an acceptable safety profile; despite this, the uptake of HRT was found to have declined [
20]. The reputational damage to HRT would appear to be such that evidence-based alternative therapeutic options for the prevention of osteoporotic fracture in post-menopausal women are desirable and could have a tangible public health benefit, particularly given the ageing global population [
21].
E2, Erα and estrogen receptor beta (Erβ) are known to play a central role in bone maintenance and turnover. These are processes that are reliant on a dynamic equilibrium between osteoblast and osteoclast activity. Exogenous signals, e.g., mechanical loading, are sensed by osteocytes. In response, osteoblasts increase their receptor activator of nuclear factor κB ligand (RANKL) expression, which activates pre-osteoclast RANK receptors, thus promoting mature osteoclast formation. Osteoclasts are activated to resorb bone, via the production of hydrogen ions and lysosomal enzymes. At the conclusion of the resorption phase, osteoclasts undergo apoptosis and an osteogenic environment dominates once more; crucially, osteoblasts express osteoprotegerin (OPG) in such an environment, which serves as a decoy to RANKL, thus limiting the binding of RANKL to its receptor RANK [
22]. Broadly, E2 is thought to protect bone mass and architecture in the following ways: (1) by stimulating osteoclast apoptosis and inhibiting apoptosis in osteoblasts and osteocytes; (2) by repressing the production of pro-osteoclastic cytokines (e.g., interleukin 1 (IL-1), interleukin 6 (IL-6) and tumour necrosis factor alpha (TNFα)) by osteoblasts; and (3) by increasing the expression of OPG [
18,
23]. Many of these effects are thought to be mediated via the interaction of E2 with Erα [
18].
Given the phytoestrogen content of shatavari and evidence of its estrogen-like effects in vivo, we hypothesised that six weeks shatavari supplementation in postmenopausal women would induce increases in in vivo molecular markers of skeletal muscle contractility and protein synthesis. We further hypothesised that sera from shatavari-supplemented participants would promote a positive bone turnover phenotype in primary human osteoblasts in vitro. Secondarily, in this initial six-week supplementation study, we measured HGS and knee extensor strength, hypothesising that alterations in muscle contractility might be evident in this timeframe.
2. Materials and Methods
2.1. Study Participants
This double-blind, placebo-controlled parallel design study was approved by the University of Exeter’s Sport and Health Sciences Research Ethics Committee (190206/B/01). Participants were recruited via social media adverts and via the local chapter of the University of the Third Age (convenience sampling). All participants gave written informed consent to participate. Twenty-four postmenopausal women (≥60 years) were screened and recruited; 20 participants (68.5 ± 6 years, body mass index (BMI) 23.3 ± 3.8) completed the study. In accordance with local ethical approval procedures, participants were free to withdraw without giving a reason; none were withdrawn due to an adverse event. Participants were pre-screened by telephone or email for self-reported exclusion criteria. Exclusion criteria were: BMI > 30; last menstrual period < 12 months previously; use of estrogenic or progestogenic hormone replacement therapy via oral, transdermal or subcutaneous implant routes in previous five years; diagnosis of osteoporosis or osteopenia; taking any medication for the prevention of osteoporosis, including bisphosphonates and non-bisphosphonates (e.g., raloxifene, denosumab, teriparatide or calcitriol). Calcium and vitamin D supplementation were permitted, but no participants reported taking such supplements. Although not an inclusion criterion, all participants completed an International Physical Activity Questionnaire (IPAQ)—Short Form as part of the participant screening process. All were classified as having ‘high’ levels of physical activity according to the evidence-based tool produced by di Blasio et al [
24].
2.2. Study Design and Supplementation Protocol
Participants attended a screening and consent visit during which eligibility was again confirmed and informed consent was obtained. Body weight and height were measured. Participants were familiarised with tests of HGS and leg strength, completing the full protocol described in subsequent sections. Participants returned for visit two having fasted overnight and abstained from caffeine consumption in the preceding 24 h. They completed tests of HGS and leg strength. A blood sample was obtained via venepuncture and a Vastus lateralis (VL) skeletal muscle biopsy was performed. A researcher who was not involved in data collection or analysis randomised participants to receive either placebo or shatavari supplementation for six weeks using a random number generator. Participants consumed two capsules per day containing either placebo (magnesium stearate, 500 mg/capsule) or shatavari (500 mg powder/capsule equivalent to 13,250 mg fresh weight shatavari root). The capsules were visually identical and did not have an odour. Compliance was assessed by providing an excess of capsules and counting the number that remained at the end of the study period. No participant missed more than 3/42 doses over the six-week supplementation period, with 11/20 participants returning the correct number of capsules. Visit three took place six weeks after visit two; the study protocol was repeated as before.
2.3. Measurement of Handgrip Strength
HGS was measured using a handgrip dynamometer according to the Southampton Protocol [
25]. Participants sat in a standard chair and rested their forearms on the armrest. The wrist was positioned just over the end of the armrest, in a neutral position with the thumb facing upwards. Each hand was tested three times and the tested hand was alternated. The best score for either hand was used for data analysis. One participant reported an injury to their dominant limb during the supplementation period. Their best efforts with their non-dominant limb were included in the data analysis.
2.4. Lower Limb Dynamometry
Assessment of lower limb knee extensor muscle strength was performed using an isokinetic dynamometer (Biodex System 3; Biodex Medical Systems, New York, NY, USA). At the familiarisation visit, participant, chair and dynamometer positioning were personalised, such that the participants sat in the chair with the ankle of their dominant leg fixed to the knee extensor attachment, knee joint in-line with the dynamometer pivot point. Leg dominance was established by asking participants which leg they would preferentially use to kick a ball. Participants’ lower backs were in contact with the chair backrest, which was positioned such that the posterior aspect of their knee had approximately 2 cm clearance from the edge of the seat. Participants were strapped into the chair and instructed to complete all movements with their arms folded across their chests, to isolate the knee extensor effort. All isokinetic movements took place over a range of motion of 80° from full flexion + 1° at the knee, at 60°·s−1 for both the concentric and eccentric phases. A warmup consisting of five sets of five concentric repetitions (1 min rest between sets) was conducted, with participants instructed to complete this at 50% effort. Following the familiarisation visit, a limit of 60% isometric maximum voluntary contraction (MVC) was placed on these sets, such the dynamometer would stop a repetition if too much effort was produced. Participants then completed three sets of three maximal isokinetic repetitions (concentric and eccentric effort; 1 min rest between sets). Following this, the knee was fixed at 90° of flexion and three MVCs were performed, separated by 1 min of rest. Force data were recorded and analysed in Spike2 ver.6 software (CED, Cambridge, UK). The best MVC, concentric isokinetic and eccentric isokinetic values attained were used for analysis.
2.5. Blood Sampling and Analysis of Bone Turnover Markers
Antecubital venous blood samples were collected into serum separator (SST) and lithium heparin (LH) containing vacutainer tubes. LH tubes were centrifuged at 4500 rpm for 15 min at 4 °C; plasma samples were aliquoted into microcentrifuge tubes. SST tubes were maintained at room temperature for 30 min prior to centrifugation at 4500 rpm for 15 min at 4 °C; serum was aliquoted into microcentrifuge tubes. Plasma and serum samples were stored at −80 °C until analysis. Carboxy-terminal cross-linking telopeptide of type I collagen (β-CTX) and procollagen type I N-terminal propeptide (P1NP) were quantified using electrochemiluminescence immunoassay on a Cobas e601 analyser (Roche Diagnostics, Mannheim, Germany), performed at the Bioanalytical Facility, University of East Anglia. The interassay coefficient of variation (CV) for β-CTX was <3% between 0.01 and 6.0 µg/L, with a sensitivity of 0.01 µng/L. The inter-assay CV for P1NP was <3% between 5 and 1200 µg/L, with a sensitivity of 5 µg/L.
2.6. Vastus lateralis Biopsy
VL muscle biopsy samples were taken from the exercised (dominant) leg using the suction-modified percutaneous Bergstrom needle technique [
26]. The leg from which the biopsy was taken was sterilised with povidone-iodine 10%
w/
w cutaneous solution (Ecolab, Northwich, UK) and anaesthetised by infiltration of the skin and subcutaneous adipose tissue with 2% lidocaine (B Braun, Sheffield, UK). An incision of approximately 0.8 cm was made, and skeletal muscle (~150 mg) collected. The incision was closed with adhesive butterfly stitches and covered with a waterproof dressing. Skeletal muscle was dissected free of any obvious blood and adipose tissue and immediately snap frozen in liquid nitrogen. Muscle samples were stored at –80 °C until analysis.
2.7. Immunoblotting
Detailed immunoblotting methods are provided in the
Supplementary Material (Supplementary Methods, Supplementary Materials Table S1). Quantifications of protein phosphorylation were normalised to total protein signal. To ensure the veracity of the phosphorylation signals obtained, the same blots were not stripped and re-probed for total and phosphorylated forms of a given protein, rather the electrophoresis and blotting was repeated. Samples were quantified in duplicate. Blot images were never adjusted to manipulate brightness/contrast, except for Coomassie stained blots, where minor adjustments were made and applied across all lanes.
2.8. Primary Human Osteoblast Cell Culture
Six femoral heads were obtained via the Royal Devon and Exeter Tissue Bank (ethical approval RDETB-REC no 6/SC/0162), which is managed within the NIHR Exeter Clinical Research Facility. Femoral head donors were postmenopausal women (73 ± 9.9 years BMI 29.6 ± 8.6) undergoing hip replacement. Exclusion criteria were: last menstrual period < 12 months previously; use of estrogenic or progestogenic hormone replacement therapy via oral, transdermal or subcutaneous implant routes in previous five years; diagnosis of osteoporosis or osteopenia; taking any medication for the prevention of osteoporosis, including bisphosphonates and non-bisphosphonates (e.g., raloxifene, denosumab, teriparatide or calcitriol). Calcium and vitamin D supplementation were permitted. Bone chips were removed from the subchondral region using a Friedman Rongeur and were washed five times in PBS containing 100 U/mL penicillin streptomycin. Bone chips were removed to 25 cm2 vented flasks containing osteoblast differentiation media: Dulbecco’s Modified Eagle Medium (DMEM) High Glucose (D6546, Merck, Gillingham, UK) 10% Fetal Bovine Serum (FBS) (qualified E.U.-approved, South-American origin) (10270106, GIBCO, Waltham, MA, USA), 100 Units/mL Penicillin/streptomycin (15070-063, GIBCO), 2 mM L-Glutamine, G7513 (Merck), 1% Minimum Essential Medium (MEM) non-essential amino acids (11140050, GIBCO, Waltham, MA, USA), β-glycerophosphate disodium salt hydrate (2 mM) (G9422, Merck, Gillingham, UK), L-Ascorbic acid (50 µg/mL) (A4403, Merck, Gillingham, UK), Dexamethasone (10 nM) (D4902, Merck, Gillingham, UK). Bone chips were cultured in a humidified incubator (37 °C, 5% CO2). Media were changed after five days and every three days thereafter, with bone chips being removed when osteoblast coverage reached approximately 30%.
2.9. Assessment of Osteoblast Differentiation
Osteoblasts were seeded at 6 × 10
3 cells per well in a 24-well plate and fed with osteoblast differentiation media containing Primary human osteoblasts were stimulated for 14 days with sera (10%
v/
v in osteoblast differentiation medium) pooled from eight participants in each supplementation group (baseline and six-week timepoints). A 10% FBS condition was included in each experiment as a previously established control to demonstrate effective differentiation [
27]. Osteoblasts were fed by 1:2 demi depletion on days 3, 6, 9 and 12; the experimental endpoint was day 14. All experiments were conducted in four to six biological replicates, with triplicate technical replicates (wells) stimulated with each serum condition for each biologically independent culture. Supernatants were retained for the quantification of protein secretion.
2.10. Osteoblast Cytokine Secretion
Macrophage colony stimulating factor (M-CSF), osteoprotegerin (OPG), IL-6 and IL-1β were quantified using enzyme-linked immunosorbent assays (ELISA) according to the manufacturer’s instructions (see
Supplementary Materials Table S2).
2.11. Osteoblast Bone Nodule Formation
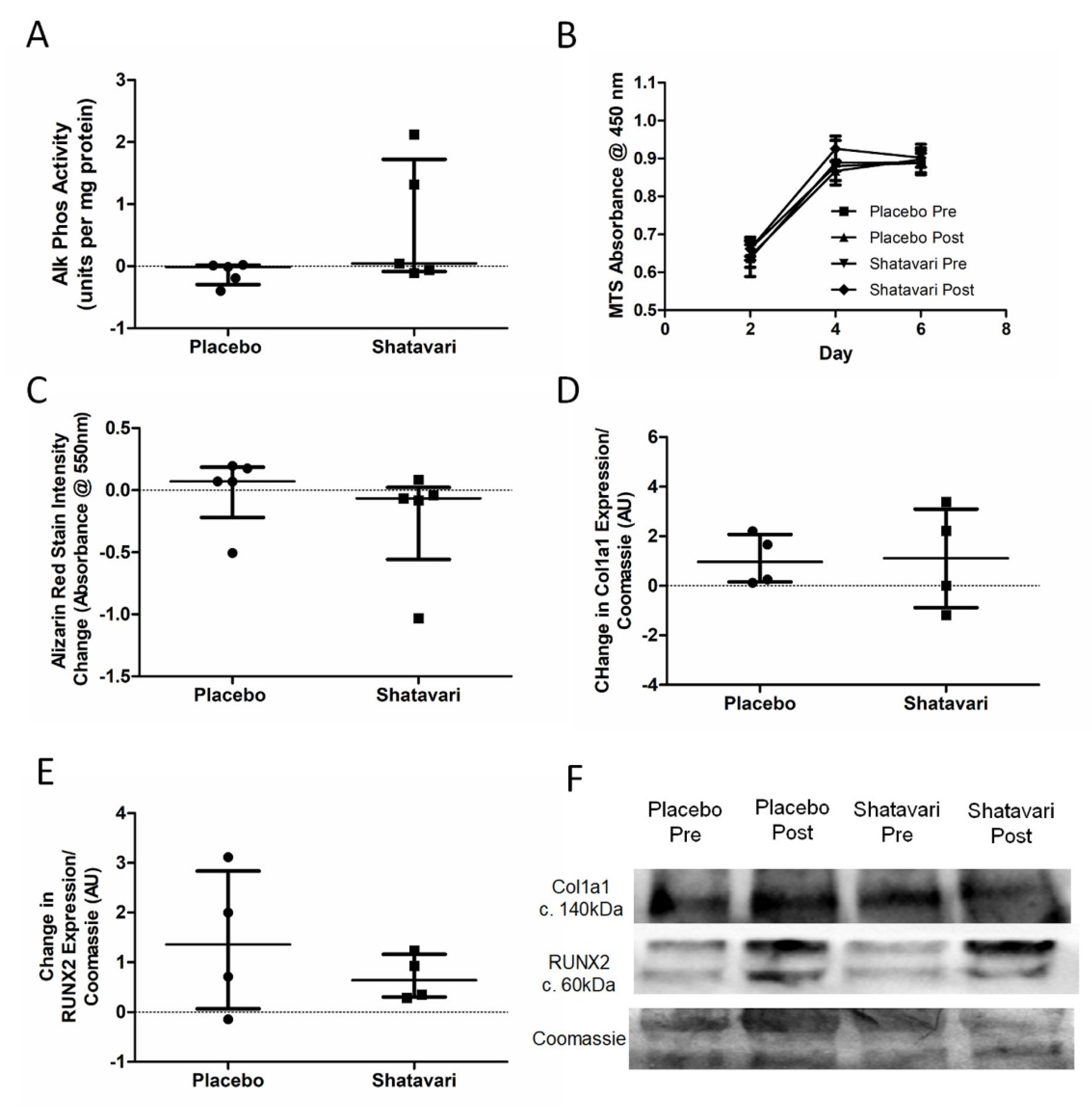
Osteoblast bone nodule formation was quantified via alizarin red staining at day 14 of differentiation. Following removal of the supernatants, osteoblasts were washed three times in phosphate buffered saline (PBS) and were stained with alizarin red staining solution (0.5% w/v alizarin red (Merck, Gillingham, UK) in 1% ammonia solution, pH 4.5) for 10 min at room temperature. Osteoblasts were again washed three times in PBS and were then incubated in 10% w/v cetyl pyridinium chloride (C0732, Merck, Gillingham, UK; warmed to 37 °C) for 10 min at room temperature. This supernatant was further diluted 1:10 in 10% w/v cetyl pyridinium chloride (warmed to 37 °C) and the absorbance of 100 μL of each solution was quantified in duplicate at 550 nm on a microplate reader (BMG LABTECH, Aylesbury, UK).
2.12. Osteoblast Metabolism
Osteoblast metabolism was quantified at day 14 of serum stimulation via an alkaline phosphatase (ALP) activity assay. Osteoblasts were lysed in radioimmunoprecipitation assay (RIPA) buffer and lysates were diluted 1:4 with 1 mM MgCl2. Seven alkaline phosphatase standards (3 units/mL top standard, serially diluted 1:2 in 1:4 RIPA:1 mM MgCl2) were prepared from a stock solution (Human placenta alkaline phosphatase, P3895, Merck, Gillingham, UK). Ten microlitres of sample or standard was added to a 96 well plate in duplicate and 100 μL alkaline phosphatase substrate (P7998, Merck, Gillingham, UK) added per well and incubated at 37 °C for 15 min. The reaction was stopped by the addition of 0.1 N NaOH and the absorbance of each well was measured at 405 nm. ALP concentrations were calculated from the standard curve and corrected for the dilution in MgCl2. The protein concentration of the lysates was quantified via bicinchoninic acid (BCA) assay and alkaline phosphatase activity expressed per unit protein.
2.13. Osteoblast Proliferation
Osteoblast proliferation was quantified via a colourimetric tetrazolium salt-based cell proliferation assay (MTS assay; CellTiter 96
® Aqueous One Solution; Promega, Madison, WI, USA). Osteoblast proliferation was assessed at two-day intervals for six days. Osteoblast differentiation medium was replaced with 100 μL serum-free differentiation medium to prevent FBS interference with absorbance readings [
28]. Twenty microliters of MTS reagent was added to each well the plate was incubated at 37 °C, 5% CO
2 for 3 h. Each condition was assayed in triplicate. The absorbance of each well was quantified at 450 nm using an absorbance microplate reader.
2.14. Statistical Analyses
Using standard deviations derived from unpublished immunoblotting data from older adults, we calculated that we required 10 participants per group to be able to detect a 1.6-fold change in myogenin protein expression; we recruited 12 participants per group to account for dropouts. Given the evidence from the literature of an effect of HRT on myogenin expression, and the lack of evidence regarding the effects of shatavari in skeletal muscle, myogenin expression was originally designated as our primary outcome measure. However, the shutdown of our lab facility due to the COVID-19 pandemic led to the loss of a large number of consumables, including the degradation of many of our immunoblotting antibody stocks that required extensive subsequent troubleshooting. Despite myogenin expression being the designated primary outcome measure, we were forced to use our limited muscle sample stocks to quantify outcomes that required less extensive troubleshooting. Data are largely quoted mean differences (pre-supplementation to post-supplementation change) for both placebo and shatavari conditions, which were analysed by means of an unpaired t-test comparison of difference scores, and they are presented as mean ± standard deviation. Where there existed non-normal distributions, unequal variances or ‘outlier’ data points data were analysed by Mann–Whitney U test and are presented as median ± interquartile range of difference scores. Data were analysed using GraphPad Prism v5.03 (GraphPad Software, San Diego, CA, USA). Shapiro–Wilk tests were used to evaluate the normality of data sets and the Levene test was employed to assess the equality of variances within data groups. Outlier data were not excluded. A p-value of <0.05 was considered statistically significant.
4. Discussion
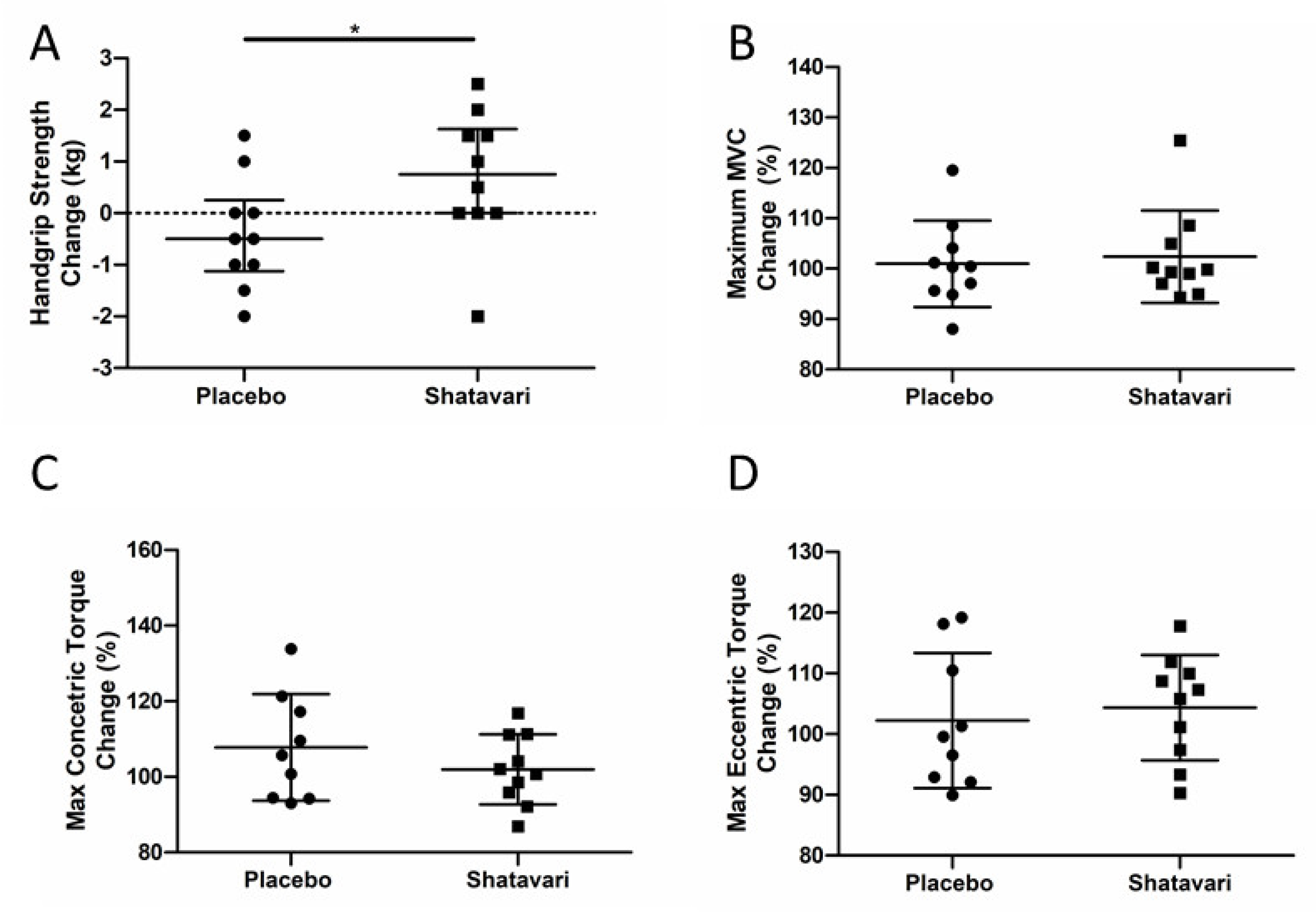
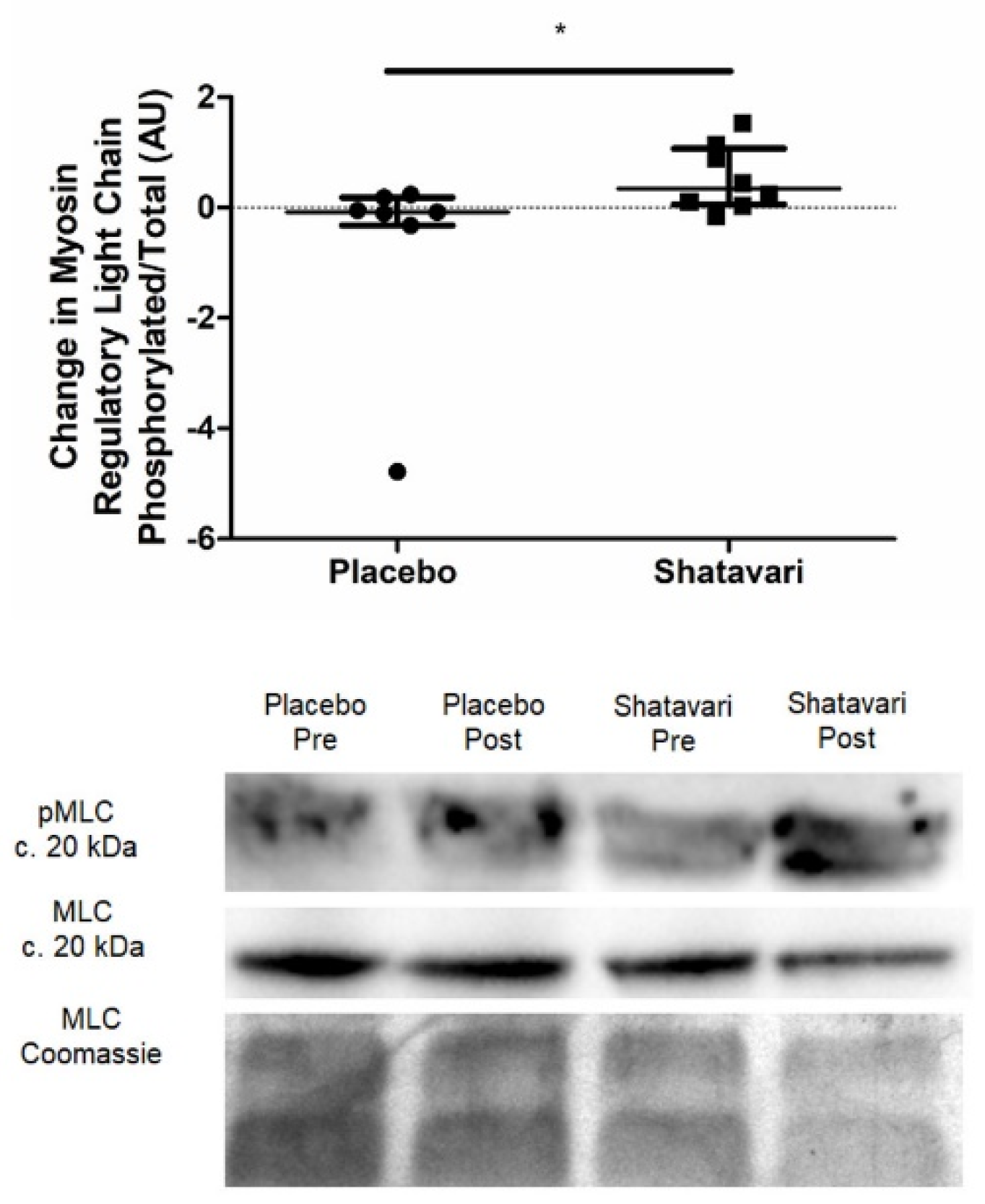
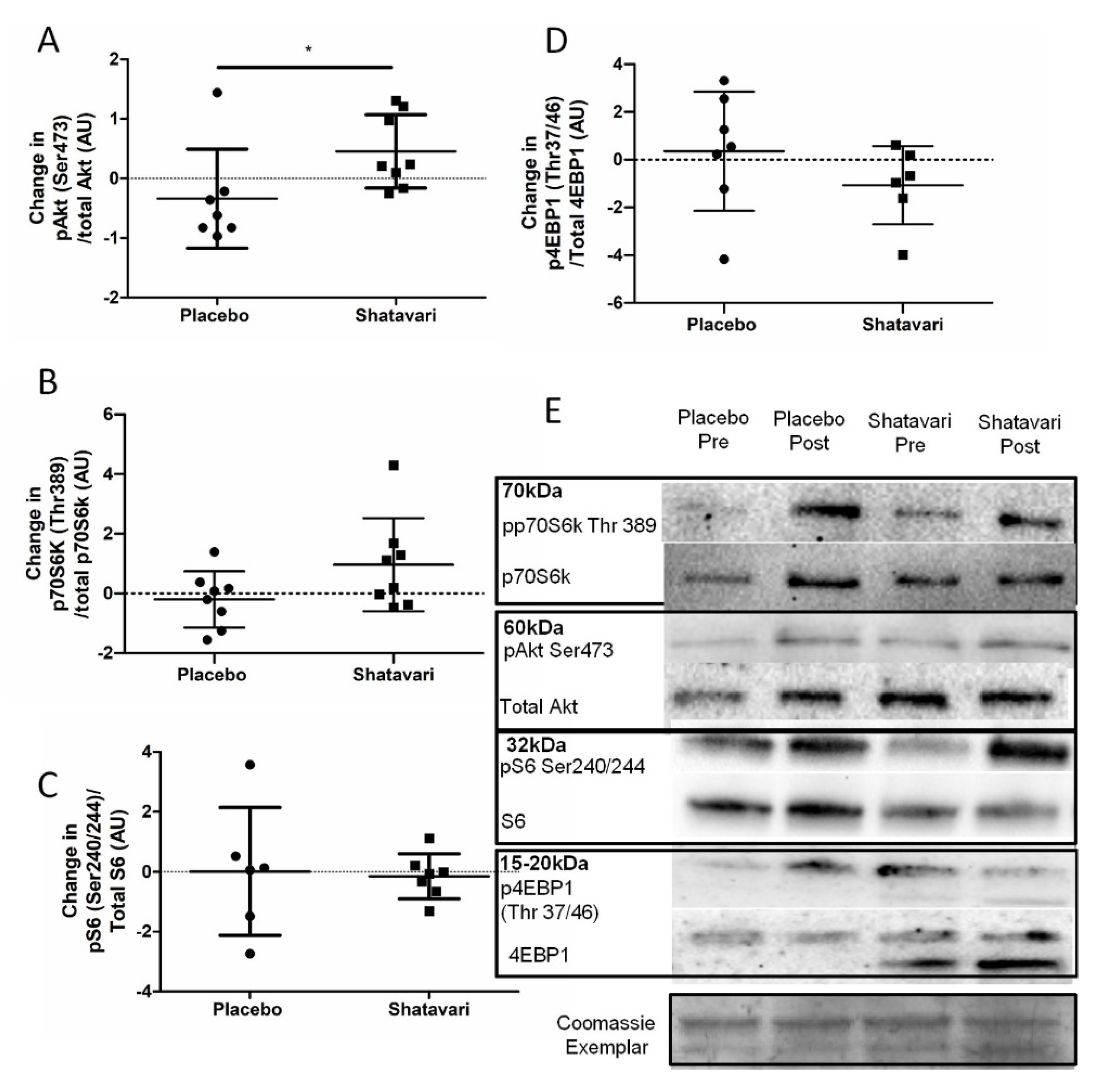
This is the first study to assess the effect of shatavari on skeletal muscle function in a cohort of older adults and the first offer insights into the molecular changes that may underpin its functional effects. To our knowledge, this is also the first study to assess the effect of shatavari supplementation on markers of bone turnover. Six weeks of shatavari supplementation improved HGS in this cohort of postmenopausal women. Although no improvement in knee extensor strength was found, we demonstrated an increase in pMLC within the VL. We also demonstrated an increase in AktSer473 phosphorylation, although no alteration in phosphorylation status of other major effectors of muscle protein synthesis (p70S6kThr389, 4EBP1Thr37/46, S6Ser240/244) was observed. We did not find any effect of shatavari-supplemented sera on markers of primary human osteoblast function.
An increase in HGS and pMLC over a period of six weeks is congruent with what is known about the effect of E2 and pMLC on muscle strength. Such phosphorylation modulates striated muscle contraction by promoting movement of the myosin head towards the thin filament, thus increasing the probability of myosin interaction with actin [
29,
30,
31]. Age-dependent reductions in pMLC have been documented and appear to particularly affect women [
32,
33]. Our observation that shatavari increased pMLC and muscle strength over six weeks is consistent with the time course of such changes documented elsewhere; Lai et al. showed that E2 administration over 4–6 weeks rescued extensor digitorum longus and soleus pMLC and force generation in bilaterally ovariectomized mice [
34]. Further, deletion of ERα reduces muscle contractility and force production in female mice [
35]. More work is required to definitively establish the link between shatavari-induced pMLC changes and human skeletal muscle contractility, e.g., via the ex vivo assessment of muscle fibre contractility and using in vivo techniques such as peripheral nerve stimulation and transcranial magnetic stimulation to assess central drive, excitability and muscle contractility.
Perhaps surprisingly in the context of the shatavari-induced changes in HGS, knee extensor strength was not increased despite an increase in
VL pMLC. Our subjective experience during this study was that the learning effect of our familiarisation session for this older adult cohort was not as effective as the same familiarisation protocol that we use regularly in younger cohorts. Indeed, there may be greater variability in reaching maximum voluntary activation in older age and our ‘best of three’ approach to such tests [
36] may need to be modified in such cohorts to take the best value from a greater number of attempts [
37]. Testing of HGS is an inherently more ‘accessible’ movement for those unfamiliar with resistance exercise and our a priori decision to use the best value from all six trials (three dominant, three non-dominant), in accordance with the ‘Southampton Protocol’ for such testing, may have mitigated this variability [
25]. Indeed, the CV for the best HGS scores achieved at visits 2 and 3 for the placebo group was 2.9 ± 2.3%; the CV for the best MVC value was 4.1 ± 4%, although this did not reach statistical significance (
p = 0.46,
t-test). Notably the CVs for concentric (8.2 ± 6.5%) and eccentric (6.6 ± 4.1%) isokinetic measures of muscle strength were significantly larger than that for HGS (
p = 0.03,
t-test for both). The lack of a forearm skeletal biopsy in the context of our HGS observations is unfortunate; however, the
VL remains the most accessible skeletal muscle for biopsy outside of a secondary/tertiary medical care facility [
38].
Intriguingly, we also observed an increase in Akt
Ser473 phosphorylation, but no alteration in phosphorylation status of other major effectors of muscle protein synthesis signalling. Interestingly in the context of this increase in Akt
Ser473 phosphorylation, it was suggested that one way in which pMLC may be increased is via the PI3K/MAPK/Akt/MLCK pathway [
34]. However, no definitive activation of the proteins downstream of mTOR was observed. We note, however, that our analysis of p70S6k
Thr389 showed an increase in its expression with shatavari supplementation with
p = 0.09. We do not consider non-significant results such as this worthy of excessive weighting in a discussion of results or in the drawing of conclusions. However, we highlight this result in the context of the relatively low number of viable biopsy samples that were available for analysis. We powered our analyses (using standard deviations derived from unpublished immunoblotting data from older adults) to be able to detect a 1.6-fold change in protein expression with 10 participants per group; we recruited 12 participants per group to account for dropouts (two dropouts occurred per group) and biopsy failures. However, even with a ‘second pass’ following a failed biopsy, we had a muscle tissue extraction failure rate of 7.5% in this population; thus, these analyses did not have the statistical power that we had anticipated. Work published since the conception of our study has shown that c. 40% of healthy older female participants are at risk of a failed muscle biopsy. Wilson et al. highlighted low muscle mass as the principal contributor to this failure [
39]. Our samples also often contained a high proportion of subcutaneous or intramuscular adipose tissue, which accumulates in skeletal muscle with advancing age [
40]. Despite these caveats, we demonstrated novel functional effects of shatavari supplementation in skeletal muscle, along with some important mechanistic insights. We consider that the potential for shatavari supplementation to enhance muscle protein synthesis should be further explored in longer-term resistance training studies, given our evidence of Akt
Ser473 phosphorylation following shatavari supplementation, coupled with the observations of Anders et al. that shatavari enhanced strength gains in young men following eight weeks of bench press training [
17]. Indeed, the balance of current evidence suggests that E2 increases the anabolic response to exercise over the longer term (for an excellent review, see [
41]. It remains to be seen what sex differences exist in the effect of shatavari on skeletal muscle function. The work of Anders et al. suggests that its effects are not limited to the female sex, despite the primacy of testosterone and its derivatives as endocrine determinants of male skeletal muscle function [
42]. It should be borne in mind that shatavari also contains steroidal saponins, racemosides, racemosol, racemofuran and asparagamine A; little is known about their effects on skeletal muscle. We did not carry out a detailed analysis of the supplement used in this study; however, having established promising biological actions of this ‘off the shelf’ form of the supplement, we recommend that this analysis should be conducted for future work. This will allow researchers to begin to disentangle complex issues such as the constituents of shatavari that are responsible for its effects in vivo and will facilitate a more standardised approach to research into its effects (e.g., dosing).
Indeed, the chemical complexity of shatavari and its metabolism in vivo is another area that warrants extensive further research. Determining the constituents of shatavari that evoke physiological responses is not a simple task. The bioavailability of phytoestrogen-containing supplements is variable and poorly understood. The intestinal flora produce both aglycone primary metabolites and secondary metabolites that potentially have a greater affinity for E2R than their parent compounds [
43]. Indeed, this is why primary human osteoblasts in this study were treated with sera from women who had supplemented with placebo or shatavari for six weeks, with their final dose taken one hour prior to venepuncture (i.e., an acute-on-chronic serum supplementation condition). In vitro studies of bioactive supplements almost universally apply extracts of the supplements directly to cell cultures. For the reasons outlined above, we consider this approach to be inappropriate and lacking in physiological relevance.
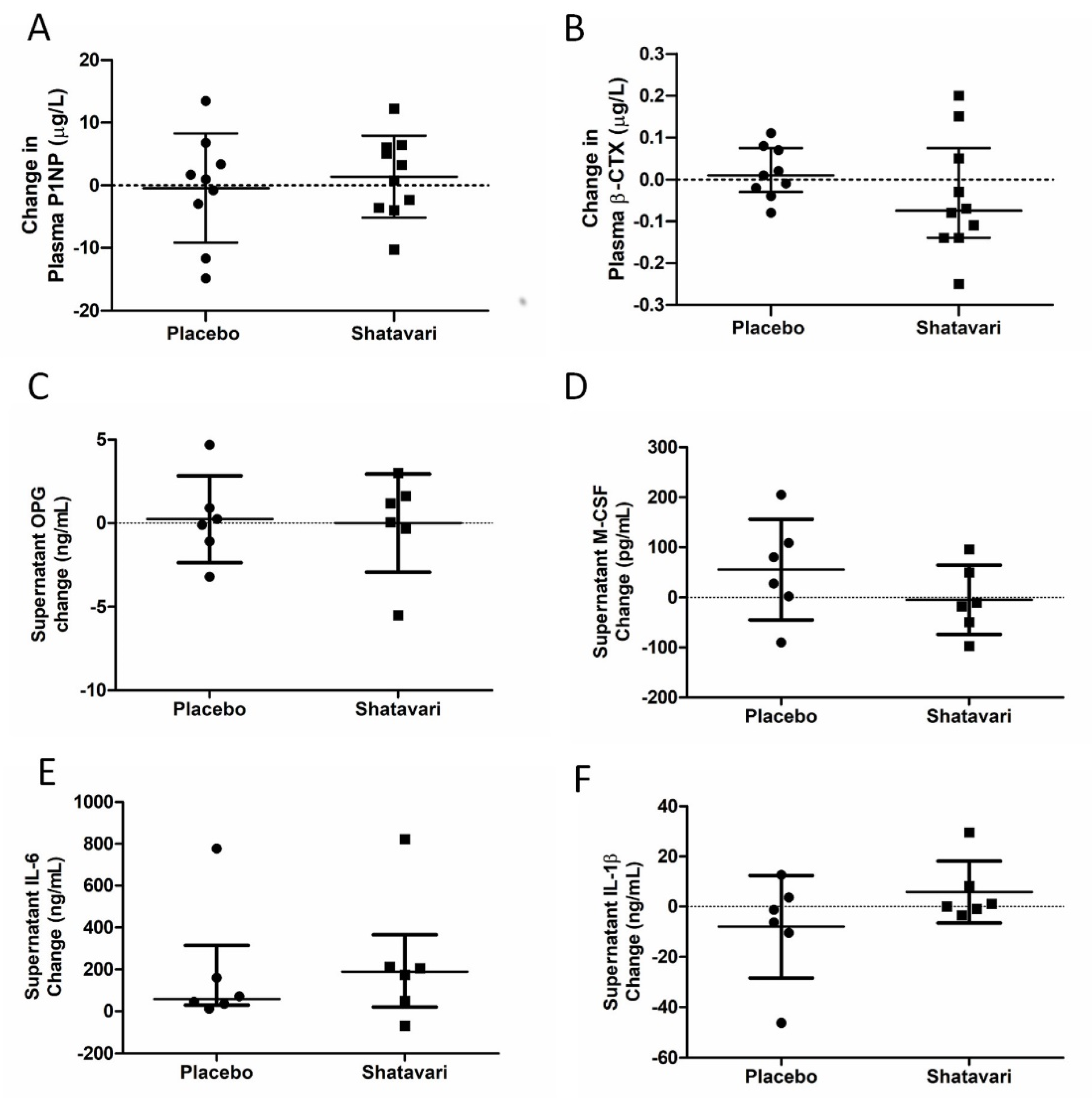
The decrease in E2 production that potentiates sarcopenia in women after the menopause is also associated with a decline in bone mineral density. Given this observation, we also examined systemic and in vitro markers of bone turnover. Here, we utilised a previously established model for assessing metabolic activity and bone nodule formation in human primary osteoblasts [
27]. We did not find any effect of shatavari-supplemented sera on markers of primary human osteoblast function, neither did we observe any changes in the osteoblast secretion of cytokines that are known to alter osteoclast activity. In this six-week supplementation study, these in vitro assays of bone turnover activity were the bone outcomes of primary interest and considered the most likely to be altered by supplement condition. However, we also quantified in vivo markers of bone turnover (P1NP and β-CTX), with no effect of supplementation being observed. Given the in vitro results that we describe, this is most likely due to a genuine lack of effect on shatavari on bone turnover. However, despite considerable efforts, in vitro models cannot perfectly reproduce human physiology and it remains possible that shatavari supplementation could have an effect on bone turnover in the longer term. Transdermal E2 treatments do induce changes in such markers over a similar timeframe. However, within-subject variability for these measures can be high (least significant change P1NP 21%, β-CTX 132%) [
44] and the potency of any effect of shatavari on such measures might be expected to be less than pharmacological doses of E2.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




