
Baseline Insulin Resistance Is a Determinant of the Small, Dense Low-Density Lipoprotein Response to Diets Differing in Saturated Fat, Protein, and Carbohydrate Contents

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Design
2.2. Laboratory Assessment
2.3. Statistics
3. Results
3.1. Baseline Characteristics
3.2. Post-Diet Characteristics
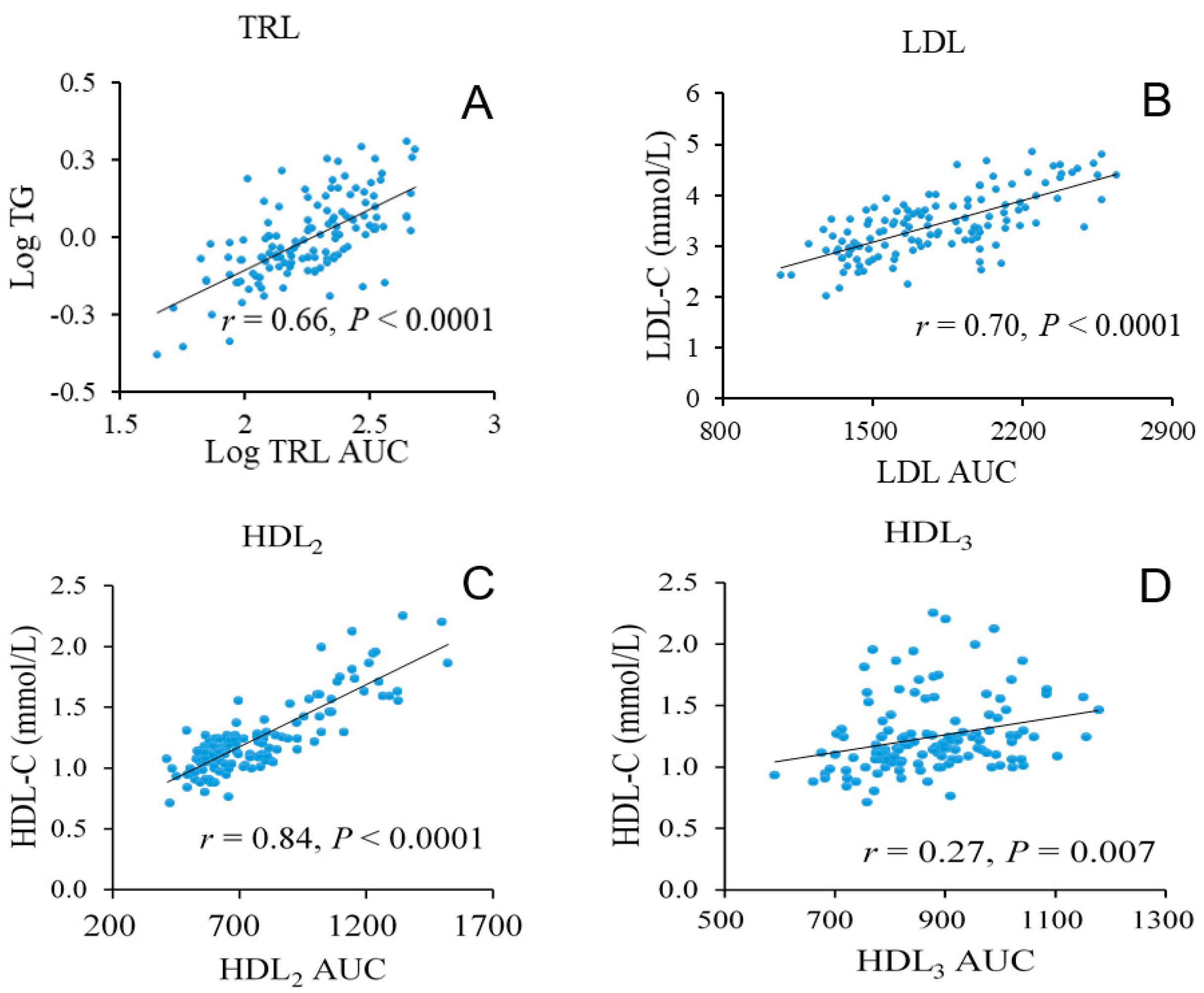
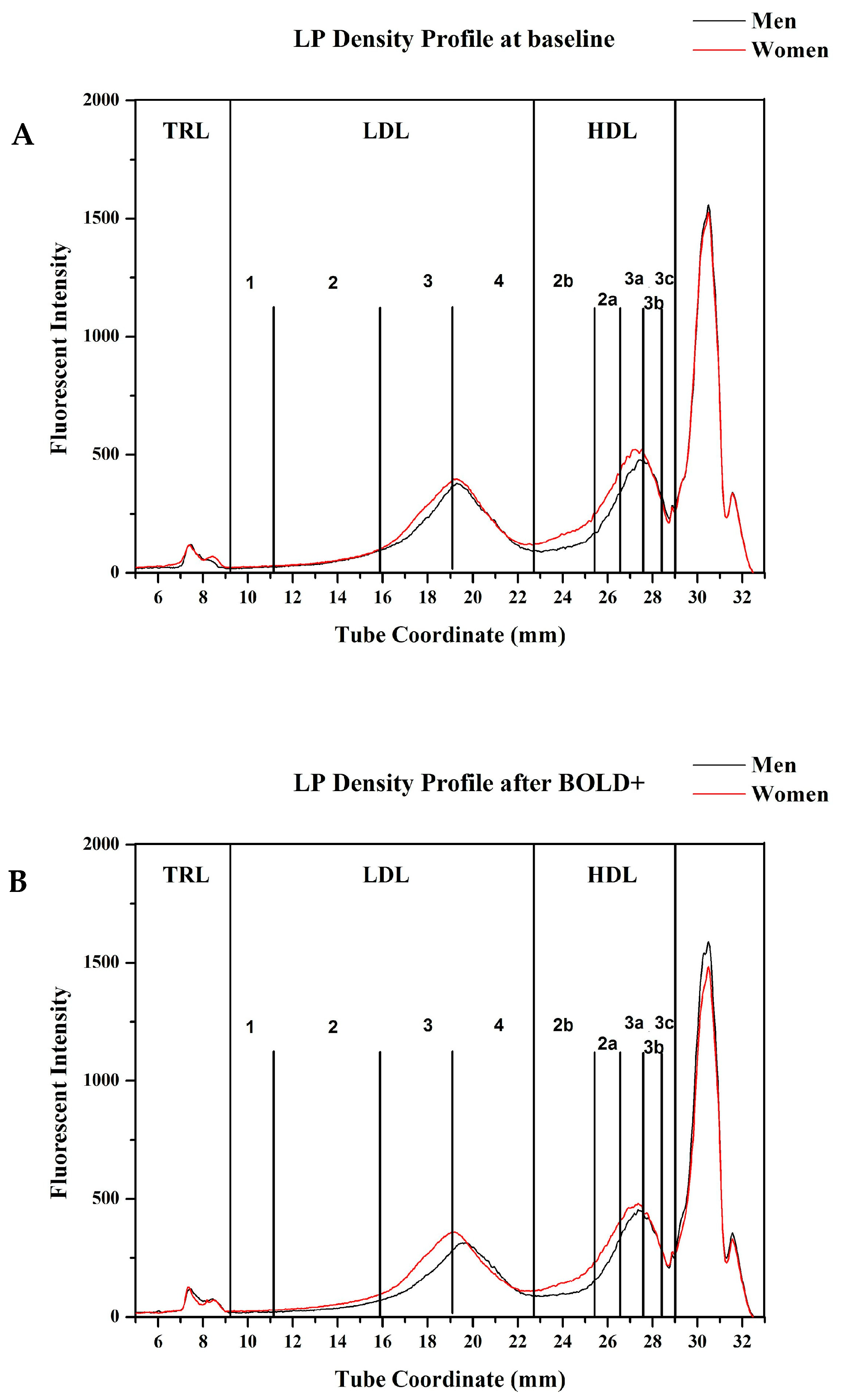
3.3. Lipoprotein Density Distribution
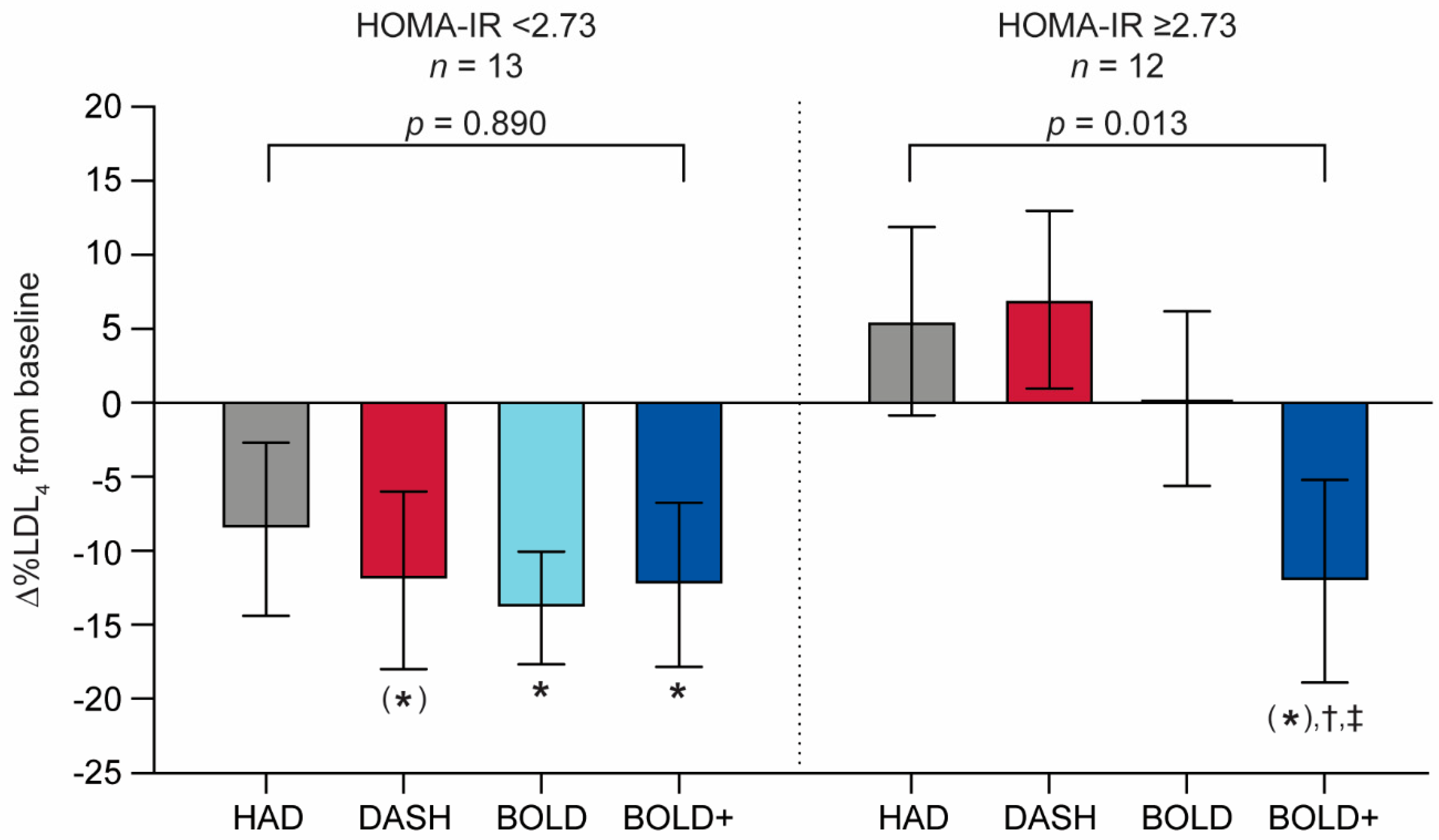
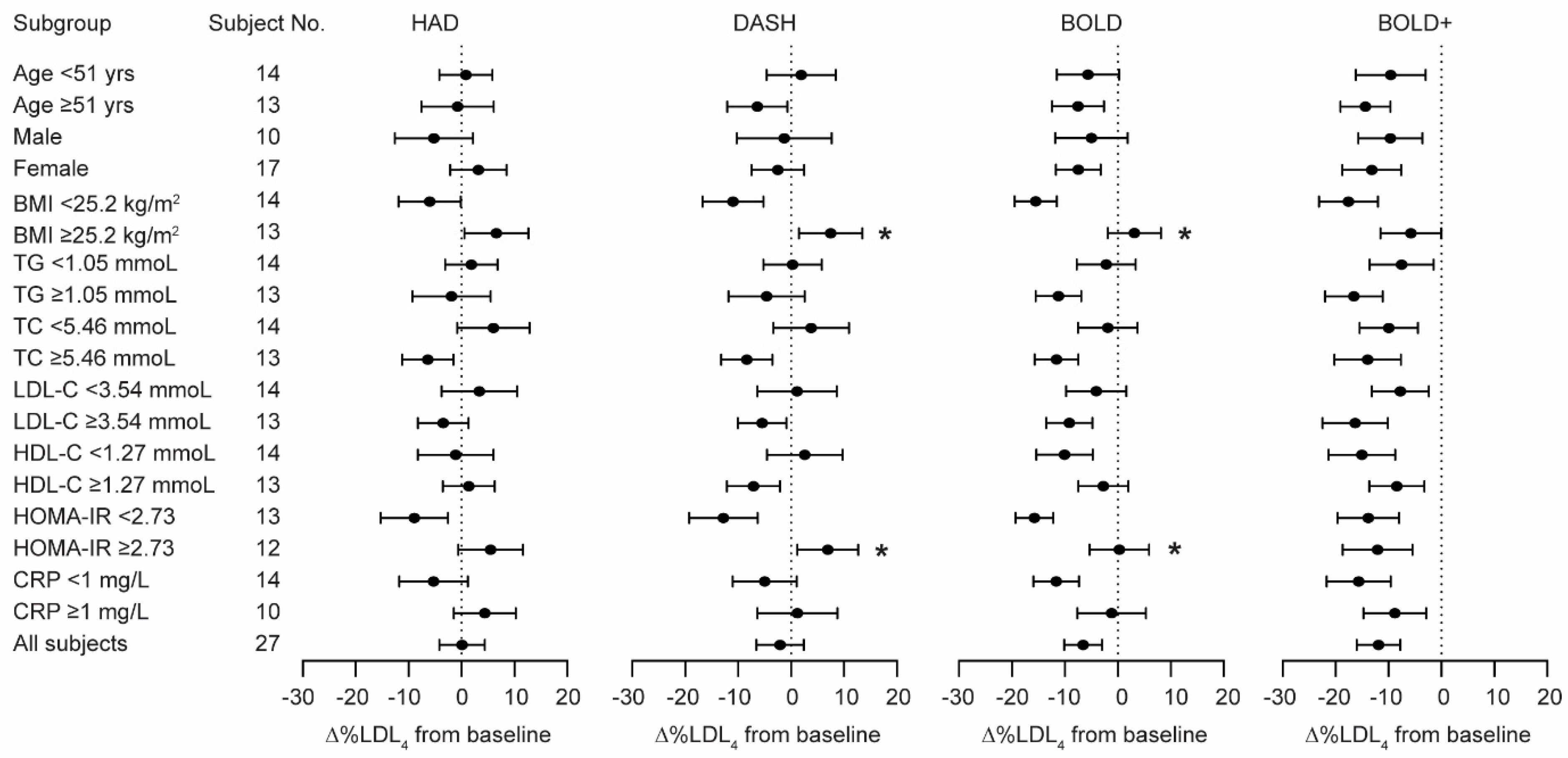
3.4. LDL4 Responsiveness
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intra-Assay (n = 3) | Inter-Assay (n = 5) | |||||
|---|---|---|---|---|---|---|
| TG | Low | Medium | High | Low | Medium | High |
| Lipoprotein distribution calculated as absolute AUC | ||||||
| TRL | 18.6 | 13.9 | 14.9 | 19.8 | 11.3 | 10.6 |
| LDL1 | 33.6 | 21.8 | 12.9 | 32.6 | 21.7 | 8.2 |
| LDL2 | 10.1 | 5.7 | 8.6 | 11.3 | 6.8 | 4.1 |
| LDL3 | 2.4 | 5.1 | 8.0 | 2.9 | 4.4 | 4.7 |
| LDL4 | 6.2 | 9.4 | 8.4 | 5.4 | 5.1 | 7.6 |
| HDL2b | 1.5 | 4.6 | 6.4 | 0.2 | 1.4 | 2.9 |
| HDL2a | 3.4 | 4.7 | 6.1 | 2.0 | 1.5 | 0.4 |
| HDL3a | 3.2 | 4.6 | 4.4 | 1.9 | 1.7 | 1.9 |
| HDL3b | 4.3 | 3.3 | 1.8 | 3.0 | 3.4 | 6.8 |
| HDL3c | 13.9 | 10.8 | 9.3 | 14.4 | 9.5 | 13.4 |
| Average | 9.7 | 8.4 | 8.1 | 9.3 | 6.7 | 6.1 |
| Lipoprotein distribution calculated as % total AUC | ||||||
| TRL | 10.8 | 8.8 | 22.9 | 9.6 | 8.8 | 19.1 |
| LDL1 | 21.9 | 19.2 | 12.2 | 21.4 | 18.1 | 11.2 |
| LDL2 | 5.9 | 4.3 | 8.0 | 4.4 | 3.4 | 4.1 |
| LDL3 | 5.0 | 5.6 | 3.9 | 5.1 | 4.9 | 5.5 |
| LDL4 | 4.3 | 4.3 | 5.9 | 3.4 | 3.5 | 4.5 |
| HDL2b | 3.2 | 1.9 | 7.2 | 2.2 | 1.9 | 5.9 |
| HDL2a | 8.7 | 4.2 | 3.8 | 7.7 | 5.0 | 5.8 |
| HDL3a | 6.1 | 2.3 | 4.3 | 5.5 | 3.4 | 3.8 |
| HDL3b | 4.6 | 2.0 | 7.3 | 4.2 | 0.8 | 4.0 |
| HDL3c | 11.7 | 8.4 | 9.9 | 9.9 | 8.2 | 6.1 |
| Average | 8.2 | 6.1 | 8.5 | 7.3 | 5.8 | 7.0 |
| Lower HOMA-IR | Higher HOMA-IR | |
|---|---|---|
| HOMA-IR | 2.1 ± 0.1 | 3.4 ± 0.2 * |
| SEX, M:F, n | 5:8 | 5:9 |
| Age (years) | 51.1 ± 2.3 | 50.8 ± 2.5 |
| BMI | 24.2 ± 0.6 | 26.9 ± 0.9 * |
| Triacylglycerol (mg/dL) | 86.1 ± 5.9 | 106.2 ± 8.2 |
| Total cholesterol (mg/dL) | 219.8 ± 7.6 | 207.2 ± 8.5 |
| LDL cholesterol (mg/dL) | 145.3 ± 5.5 | 137.2 ± 7.2 |
| HDL cholesterol (mg/dL) | 57.3 ± 4.2 | 48.9 ± 3.3 |
| hsCRP (mg/L) | 1.1 ± 0.3 | 1.6 ± 0.4 |
References
- La Belle, M.; Krauss, R.M. Differences in carbohydrate content of low density lipoproteins associated with low density lipoprotein subclass patterns. J. Lipid Res. 1990, 31, 1577–1588. [Google Scholar] [CrossRef]
- Chait, A.; Brazg, R.L.; Tribble, D.L.; Krauss, R.M. Susceptibility of small, dense, low-density lipoproteins to oxidative modification in subjects with the atherogenic lipoprotein phenotype, pattern B. Am. J. Med. 1993, 94, 350–356. [Google Scholar] [CrossRef]
- Nigon, F.; Lesnik, P.; Rouis, M.; Chapman, M.J. Discrete subspecies of human low density lipoproteins are heterogeneous in their interaction with the cellular LDL receptor. J. Lipid Res. 1991, 32, 1741–1753. [Google Scholar] [CrossRef]
- Borén, J.; Chapman, M.J.; Krauss, R.M.; Packard, C.J.; Bentzon, J.F.; Binder, C.J.; Daemen, M.J.; Demer, L.L.; Hegele, R.A.; Nicholls, S.J.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease: Pathophysiological, genetic, and therapeutic insights: A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2020, 41, 2313–2330. [Google Scholar] [CrossRef]
- Dreon, D.M.; Fernstrom, H.A.; Miller, B.; Krauss, R.M. Apolipoprotein-E isoform phenotype and LDL subclass response to a reduced-fat diet. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 105–111. [Google Scholar] [CrossRef]
- Dreon, D.M.; Fernstrom, H.A.; Williams, P.T.; Krauss, R.M. Reduced LDL particle size in children consuming a very-low-fat diet is related to parental LDL-subclass patterns. Am. J. Clin. Nutr. 2000, 71, 1611–1616. [Google Scholar] [CrossRef]
- KasimKarakas, S.E.; Lane, E.; Almario, R.; Mueller, W.; Walzem, R. Effects of dietary fat restriction on particle size of plasma lipoproteins in postmenopausal women. Metab. Clin. Exp. 1997, 46, 431–436. [Google Scholar] [CrossRef]
- Krauss, R.M.; Blanche, P.J.; Rawlings, R.S.; Fernstrom, H.S.; Williams, P.T. Separate effects of reduced carbohydrate intake and weight loss on atherogenic dyslipidemia. Am. J. Clin. Nutr. 2006, 83, 1025–1031. [Google Scholar] [CrossRef] [Green Version]
- Krauss, R.M.; Dreon, D.M. Low-density-lipoprotein subclasses and response to a low-fat diet in healthy-men. Am. J. Clin. Nutr. 1995, 62, S478–S487. [Google Scholar] [CrossRef] [Green Version]
- Chapman, M.J.; Laplaud, P.M.; Luc, G.; Forgez, P.; Bruckert, E.; Goulinet, S.; Lagrange, D. Further resolution of the low density lipoprotein spectrum in normal human plasma: Physicochemical characteristics of discrete subspecies separated by density gradient ultracentrifugation. J. Lipid Res. 1988, 29, 442–458. [Google Scholar] [CrossRef]
- Nunn, A.V.; Bell, J.D.; Guy, G.W. Lifestyle-induced metabolic inflexibility and accelerated ageing syndrome: Insulin resistance, friend or foe? Nutr. Metab. 2009, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Gower, B.A.; Bergman, R.; Stefanovski, D.; Darnell, B.; Ovalle, F.; Fisher, G.; Sweatt, S.K.; Resuehr, H.S.; Pelkman, C. Baseline insulin sensitivity affects response to high-amylose maize resistant starch in women: A randomized, controlled trial. Nutr. Metab. 2016, 13, 2. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Qi, Y.; Huang, C.; Wu, M.; Wang, C.; Li, F.; Yang, C.; Yan, L.; Ren, M.; Sun, K. Associations of lipid parameters with insulin resistance and diabetes: A population-based study. Clin. Nutr. 2018, 37, 1423–1429. [Google Scholar] [CrossRef]
- Palomba, S.; Falbo, A.; Chiossi, G.; Muscogiuri, G.; Fornaciari, E.; Orio, F.; Tolino, A.; Colao, A.; La Sala, G.B.; Zullo, F. Lipid profile in nonobese pregnant women with polycystic ovary syndrome: A prospective controlled clinical study. Steroids 2014, 88, 36–43. [Google Scholar] [CrossRef]
- American Diabetes, A. 2. Classification and diagnosis of diabetes. Diabetes Care 2016, 39 (Suppl. 1), S13–S22. [Google Scholar] [CrossRef] [Green Version]
- Ferrannini, E.; Mari, A. Physiology of insulin secretion. In Williams Textbook of Endocrinology, 14th ed.; Shlomo Melmed, S., Auchus, R.J., Goldfine, A.B., Koenig, R.J., Rosen, C.J., Eds.; Elsevier: Philadelphia, PA, USA, 2020; pp. 1338–1348.e33. [Google Scholar]
- Appel, L.J.; Moore, T.J.; Obarzanek, E.; Vollmer, W.M.; Svetkey, L.P.; Sacks, F.M.; Bray, G.A.; Vogt, T.M.; Cutler, J.A.; Windhauser, M.M.; et al. A clinical trial of the effects of dietary patterns on blood pressure. DASH Collaborative Research Group. N. Engl. J. Med. 1997, 336, 1117–1124. [Google Scholar] [CrossRef] [Green Version]
- Conlin, P.R.; Chow, D.; Miller, E.R.; Svetkey, L.P.; Lin, P.H.; Harsha, D.W.; Moore, T.J.; Sacks, F.M.; Appel, L.J.; Grp, D.R. The effect of dietary patterns on blood pressure control in hypertensive patients: Results from the Dietary Approaches to Stop Hypertension (DASH) trial. Am. J. Hypertens. 2000, 13, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Kaluza, J.; Akesson, A.; Wolk, A. Processed and unprocessed red meat consumption and risk of heart failure prospective study of men. Circ. Heart. Fail. 2014, 7, 552–557. [Google Scholar] [CrossRef] [Green Version]
- Maki, K.C.; Slavin, J.L.; Rains, T.M.; Kris-Etherton, P.M. Limitations of observational evidence: Implications for evidence-based dietary recommendations. Adv. Nutr. 2014, 5, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Rohrmann, S.; Overvad, K.; Bueno-De-Mesquita, H.B.; Jakobsen, M.U.; Egeberg, R.; Tjonneland, A.; Nailler, L.; Boutron-Ruault, M.C.; Clavel-Chapelon, F.; Krogh, V.; et al. Meat consumption and mortality—results from the European prospective investigation into cancer and nutrition. BMC Med. 2013, 11, 63. [Google Scholar] [CrossRef]
- Roussell, M.A.; Hill, A.M.; Gaugler, T.L.; West, S.G.; Heuvel, J.P.; Alaupovic, P.; Gillies, P.J.; Kris-Etherton, P.M. Beef in an optimal lean diet study: Effects on lipids, lipoproteins, and apolipoproteins. Am. J. Clin. Nutr. 2012, 95, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roussell, M.A.; Hill, A.M.; Gaugler, T.L.; West, S.G.; Ulbrecht, J.S.; Vanden Heuvel, J.P.; Gillies, P.J.; Kris-Etherton, P.M. Effects of a DASH-like diet containing lean beef on vascular health. J. Hum. Hypertens. 2014, 28, 600–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiterovich, P.O. Clinical relevance of the biochemical, metabolic, and genetic factors that influence low-density lipoprotein heterogeneity. Am. J. Cardiol. 2002, 90, 30i–47i. [Google Scholar] [CrossRef]
- Morgan, C.R.; Lazarow, A. Immunoassay of Insulin Using a Two-Antibody System. Proc. Soc. Exp. Biol. Med. 1962, 110, 29–32. [Google Scholar] [CrossRef]
- Kraus, R.M.; Stallings, H.W.; Yeager, R.C.; Gavin, T.P. Circulating plasma VEGF response to exercise in sedentary and endurance-trained men. J. Appl. Physiol. 2004, 96, 1445–1450. [Google Scholar] [CrossRef]
- Wallace, T.M.; Levy, J.C.; Matthews, D.R. Use and abuse of HOMA modeling. Diabetes Care 2004, 27, 1487–1495. [Google Scholar] [CrossRef] [Green Version]
- Heinze, C.R.; Hawkins, M.G.; Gillies, L.A.; Wu, X.; Walzem, R.L.; German, J.B.; Klasing, K.C. Effect of dietary omega-3 fatty acids on red blood cell lipid composition and plasma metabolites in the cockatiel, Nymphicus hollandicus. J. Anim. Sci. 2012, 90, 3068–3079. [Google Scholar] [CrossRef]
- Berneis, K.K.; Krauss, R.M. Metabolic origins and clinical significance of LDL heterogeneity. J. Lipid Res. 2002, 43, 1363–1379. [Google Scholar] [CrossRef] [Green Version]
- Guerin, M.; Bruckert, E.; Dolphin, P.J.; Turpin, G.; Chapman, M.J. Fenofibrate reduces plasma cholesteryl ester transfer from HDL to VLDL and normalizes the atherogenic, dense LDL profile in combined hyperlipidemia. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 763–772. [Google Scholar] [CrossRef]
- Orsoni, A.; Saheb, S.; Levels, J.H.; Dallinga-Thie, G.; Atassi, M.; Bittar, R.; Robillard, P.; Bruckert, E.; Kontush, A.; Carrie, A.; et al. LDL-apheresis depletes apoE-HDL and pre-beta1-HDL in familial hypercholesterolemia: Relevance to atheroprotection. J. Lipid Res. 2011, 52, 2304–2313. [Google Scholar] [CrossRef] [Green Version]
- Veniant, M.M.; Sullivan, M.A.; Kim, S.K.; Ambroziak, P.; Chu, A.; Wilson, M.D.; Hellerstein, M.K.; Rudel, L.L.; Walzem, R.L.; Young, S.G. Defining the atherogenicity of large and small lipoproteins containing apolipoprotein B100. J. Clin. Investig. 2000, 106, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Lamarche, B.; St-Pierre, A.C.; Ruel, I.L.; Cantin, B.; Dagenais, G.R.; Despres, J.P. A prospective, population-based study of low density lipoprotein particle size as a risk factor for ischemic heart disease in men. Can. J. Cardiol. 2001, 17, 859–865. [Google Scholar]
- St-Pierre, A.C.; Ruel, I.L.; Cantin, B.; Dagenais, G.R.; Bernard, P.M.; Despres, J.P.; Lamarche, B. Comparison of various electrophoretic characteristics of LDL particles and their relationship to the risk of ischemic heart disease. Circulation 2001, 104, 2295–2299. [Google Scholar] [CrossRef] [Green Version]
- Dumesnil, J.G.; Turgeon, J.; Tremblay, A.; Poirier, P.; Gilbert, M.; Gagnon, L.; St-Pierre, S.; Garneau, C.; Lemieux, I.; Pascot, A.; et al. Effect of a low-glycaemic index-low-fat-high protein diet on the atherogenic metabolic risk profile of abdominally obese men. Br. J. Nutr. 2001, 86, 557–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtado, J.D.; Campos, H.; Appel, L.J.; Miller, E.R.; Laranjo, N.; Carey, V.J.; Sacks, F.M. Effect of protein, unsaturated fat, and carbohydrate intakes on plasma apolipoprotein B and VLDL and LDL containing apolipoprotein C-III: Results from the OmniHeart Trial. Am. J. Clin. Nutr. 2008, 87, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, M.; Champagne, C.M.; Tulley, R.T.; Rood, J.C.; Most, M.M. Individual variability in cardiovascular disease risk factor responses to low-fat and low-saturated-fat diets in men: Body mass index, adiposity, and insulin resistance predict changes in LDL cholesterol. Am. J. Clin. Nutr. 2005, 82, 957–963. [Google Scholar] [CrossRef]
- Maki, K.C.; Palacios, O.M.; Lindner, E.; Nieman, K.M.; Bell, M.; Sorce, J. Replacement of refined starches and Added sugars with egg protein and unsaturated fats increases insulin sensitivity and lowers triglycerides in overweight or obese adults with elevated triglycerides. J. Nutr. 2017, 147, 1267–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniyappa, R.; Lee, S.; Chen, H.; Quon, M.J. Current approaches for assessing insulin sensitivity and resistance in vivo: Advantages, limitations, and appropriate usage. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E15–E26. [Google Scholar] [CrossRef] [Green Version]
- Ascaso, J.F.; Pardo, S.; Real, J.T.; Lorente, R.I.; Priego, A.; Carmena, R. Diagnosing insulin resistance by simple quantitative methods in subjects with normal glucose metabolism. Diabetes Care 2003, 26, 3320–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, L.R.; Olsen, R.H.; Anholm, C.; Walzem, R.L.; Fenger, M.; Eugen-Olsen, J.; Haugaard, S.B.; Prescott, E. Weight loss is superior to exercise in improving the atherogenic lipid profile in a sedentary, overweight population with stable coronary artery disease: A randomized trial. Atherosclerosis 2016, 246, 221–228. [Google Scholar] [CrossRef]
- Hill, A.M.; Harris Jackson, K.A.; Roussell, M.A.; West, S.G.; Kris-Etherton, P.M. Type and amount of dietary protein in the treatment of metabolic syndrome: A randomized controlled trial. Am. J. Clin. Nutr. 2015, 102, 757–770. [Google Scholar] [CrossRef] [Green Version]
- Berglund, L.; Oliver, E.H.; Fontanez, N.; Holleran, S.; Matthews, K.; Roheim, P.S.; Ginsberg, H.N.; Ramakrishman, R.; Lefevre, M.; Investigators, D. HDL-subpopulation patterns in response to reductions in dietary total and saturated fat intakes in healthy subjects. Am. J. Clin. Nutr. 1999, 70, 992–1000. [Google Scholar]
- Davidson, W.S.; Silva, R.A.; Chantepie, S.; Lagor, W.R.; Chapman, M.J.; Kontush, A. Proteomic analysis of defined HDL subpopulations reveals particle-specific protein clusters: Relevance to antioxidative function. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Aikawa, M.; Libby, P.; Alcaide, P.; Luscinskas, F.W.; Sacks, F.M. Apolipoprotein CIII in apolipoprotein B lipoproteins enhances the adhesion of human monocytic cells to endothelial cells. Circulation 2006, 113, 691–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, M.K.; Rimm, E.B.; Furtado, J.D.; Sacks, F.M. Apolipoprotein C-III as a potential modulator of the association between HDL-cholesterol and incident coronary heart disease. J. Am. Heart. Assoc. 2012, 1, jah3-e000232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Department of Agriculture, A.R.S. FoodData Central; United States Department of Agriculture: Washington, DC, USA, 2019. [Google Scholar]
- Combs, G.F.; McClung, J.P. Vitamins: Fundamental Aspects in Nutrition and Health, 5th ed.; Academic Press: Cambridge, UK, 2017. [Google Scholar] [CrossRef]
- O’Dell, B.L.; Sunde, R.A. Handbook of Nutritionaly Essential Mineral Elements; Marcel Dekker Inc.: New York, NY, USA, 1997; Volume 1, p. 692. [Google Scholar]


| HAD | DASH | BOLD | BOLD+ | |
|---|---|---|---|---|
| 95% Lean beef (g/day) | 20 | 28 | 113 | 153 |
| Calories (kcal) | 2097 | 2106 | 2100 | 2104 |
| Protein (%E) | 17 | 18 | 19 | 27 |
| Carbohydrate (%E) | 50 | 55 | 54 | 45 |
| Fat (%E) | 33 | 27 | 28 | 28 |
| SFA (%E) | 12 | 6 | 6 | 6 |
| MUFA (%E) | 11 | 9 | 11 | 12 |
| PUFA (%E) | 7 | 8 | 7 | 7 |
| Men (n = 10) | Women (n = 17) | Total (n = 27) | |
|---|---|---|---|
| Age (years) | 50.9 ± 2.6 | 50.9 ± 2.2 | 50.9 ± 1.7 |
| BMI | 26.5 ± 0.8 | 25.1 ± 0.9 | 25.6 ± 0.6 |
| TG (mmol/L) | 1.08 ± 0.10 | 1.10 ± 0.08 | 1.09 ± 0.06 |
| TC (mmol/L) | 4.98 ± 0.15 | 5.84 ± 0.18 * | 5.52 ± 0.15 |
| LDL-C (mmol/L) | 3.26 ± 0.13 | 3.89 ± 0.14 * | 3.65 ± 0.12 |
| HDL-C (mmol/L) | 1.23 ± 0.07 | 1.45 ± 0.10 | 1.37 ± 0.07 |
| ApoB (mg/dL) | 89.0 ± 2.73 | 97.9 ± 1.82 * | 94.5 ± 8.46 |
| Glucose (mmol/L) | 4.94 ± 0.08 | 4.50 ± 0.07 | 4.68 ± 0.05 |
| Insulin (mU/L) | 12.70 ± 1.50 | 13.20 ± 0.70 | 13.00 ± 0.70 |
| HOMA-IR | 2.79 ± 0.34 | 2.65 ± 0.16 | 2.71 ± 0.17 |
| hsCRP (mg/L) | 1.00 ± 0.16 | 1.09 ± 0.22 | 1.06 ± 0.15 |
| HAD (n = 27) | DASH (n = 27) | BOLD (n = 27) | BOLD+ (n = 27) | |
|---|---|---|---|---|
| BMI | 24.5 ± 0.70 | 25.3 ± 0.60 | 25.5 ± 0.60 | 25.4 ± 0.60 |
| TG (mmol/L) | 1.02 ± 0.07 | 1.04 ± 0.06 | 1.01 ± 0.06 | 0.98 ± 0.06 |
| TC (mmol/L) | 5.26 ± 0.16 | 5.01 ± 0.16 * | 5.04 ± 0.17 * | 4.96 ± 0.16 *,† |
| LDL-C (mmol/L) | 3.51 ± 0.12 | 3.30 ± 0.11 * | 3.34 ± 0.13 * | 3.29 ± 0.12 * |
| HDL-C (mmol/L) | 1.28 ± 0.06 | 1.19 ± 0.06 *,† | 1.22 ± 0.06 * | 1.20 ± 0.06 *,† |
| ApoB (mg/dL) | 95.1 ± 2.8 | 92.2 ± 2.8 | 92.0 ± 3.5 | 91.8 ± 2.9 |
| Glucose (mmol/L) | 4.83 ± 0.08 | 4.75 ± 0.07 | 4.83 ± 0.07 | 4.91 ± 0.08 |
| Insulin (IU/mL) | 12.8 ± 0.8 | 12.3 ± 0.7 | 13.6 ± 0.8 | 13.4 ± 0.7 |
| HOMA-IR | 2.76 ± 0.20 | 2.64 ± 0.14 | 2.93 ± 0.17 | 2.94 ± 0.17 |
| hsCRP (mg/L) | 1.07 ± 0.25 | 0.99 ± 0.19 | 0.92 ± 0.13 | 0.93 ± 0.12 |
| Baseline (n = 27) | HAD (n = 27) | DASH (n = 27) | BOLD (n = 27) | BOLD+ (n = 27) | |
|---|---|---|---|---|---|
| Lipoprotein distribution calculated as absolute AUC | |||||
| TRL | 216 ± 16 | 204 ± 22 | 214 ± 14 | 181 ± 14 | 195 ± 23 |
| LDL1 | 56 ± 5 | 44 ± 3 | 47 ± 4 | 45 ± 4 | 52 ± 5 |
| LDL2 | 274 ± 15 | 240 ± 15 | 232 ± 14 * | 226 ± 14 * | 253 ± 21 |
| LDL3 | 780 ± 49 | 730 ± 47 | 665 ± 48 * | 667 ± 48 * | 687 ± 38 |
| LDL4 | 844 ± 39 | 822 ± 35 | 797 ± 21 | 761 ± 21 | 723 ± 33 *,† |
| HDL2b | 369 ± 28 | 370 ± 28 | 337 ± 22 *,† | 331 ± 22 *,† | 345 ± 29 |
| HDL2a | 438 ± 22 | 422 ± 24 | 406 ± 20 | 408 ± 20 | 420 ± 26 |
| HDL3a | 422 ± 12 | 395 ± 12 | 408 ± 12 | 394 ± 12 | 399 ± 12 |
| HDL3b | 364 ± 9 | 343 ± 9 | 353 ± 8 * | 329 ± 8 * | 336 ± 9 * |
| HDL3c | 128 ± 5 | 127 ± 5 | 129 ± 4 | 118 ± 4 | 124 ± 5 |
| Lipoprotein distribution calculated as % total AUC | |||||
| TRL | 5.5 ± 0.4 | 5.6 ± 0.6 | 5.9 ± 0.4 | 5.3 ± 0.4 | 5.6 ± 0.6 |
| LDL1 | 1.5 ± 0.1 | 1.2 ± 0.1 | 1.3 ± 0.1 | 1.3 ± 0.1 | 1.5 ± 0.1 |
| LDL2 | 7.1 ± 0.3 | 6.4 ± 0.3 | 6.4 ± 0.3 | 6.5 ± 0.3 | 7.0 ± 0.3 |
| LDL3 | 19.8 ± 0.9 | 19.4 ± 0.8 | 18.3 ± 1.0 | 18.9 ± 1.0 | 19.1 ± 0.7 |
| LDL4 | 21.8 ± 0.8 | 22.5 ± 0.8 | 22.4 ± 0.4 | 22.4 ± 0.4 | 20.8 ± 0.5 †,‡,§ |
| HDL2b | 9.3 ± 0.4 | 9.9 ± 0.4 | 9.3 ± 0.3 | 9.4 ± 0.3 | 9.5 ± 0.5 |
| HDL2a | 11.2 ± 0.2 | 11.3 ± 0.2 | 11.3 ± 0.2 | 11.7 ± 0.2 | 11.8 ± 0.3 |
| HDL3a | 10.9 ± 0.2 | 10.7 ± 0.2 | 11.5 ± 0.2 *,† | 11.5 ± 0.2 *,† | 11.5 ± 0.3 † |
| HDL3b | 9.5 ± 0.2 | 9.4 ± 0.3 | 9.9 ± 0.2 | 9.6 ± 0.2 | 9.7 ± 0.3 |
| HDL3c | 3.4 ± 0.1 | 3.5 ± 0.2 | 3.6 ± 0.1 | 3.5 ± 0.1 | 3.6 ± 0.1 |
| HAD | DASH | BOLD | BOLD+ | |||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| TG | 0.16 | 0.428 | 0.02 | 0.922 | 0.15 | 0.445 | 0.16 | 0.427 |
| TC | 0.16 | 0.412 | 0.27 | 0.161 | 0.13 | 0.502 | 0.21 | 0.300 |
| LDL-C | 0.12 | 0.536 | 0.22 | 0.271 | 0.24 | 0.222 | 0.25 | 0.205 |
| HDL-C | 0.21 | 0.299 | 0.21 | 0.288 | 0.18 | 0.356 | 0.05 | 0.813 |
| HOMA-IR | 0.36 | 0.065 | 0.56 | 0.003 | 0.47 | 0.018 | 0.13 | 0.533 |
| hsCRP | 0.35 | 0.093 | 0.23 | 0.282 | 0.32 | 0.125 | 0.18 | 0.395 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Roussell, M.A.; Hill, A.M.; Kris-Etherton, P.M.; Walzem, R.L. Baseline Insulin Resistance Is a Determinant of the Small, Dense Low-Density Lipoprotein Response to Diets Differing in Saturated Fat, Protein, and Carbohydrate Contents. Nutrients 2021, 13, 4328. https://doi.org/10.3390/nu13124328
Wu X, Roussell MA, Hill AM, Kris-Etherton PM, Walzem RL. Baseline Insulin Resistance Is a Determinant of the Small, Dense Low-Density Lipoprotein Response to Diets Differing in Saturated Fat, Protein, and Carbohydrate Contents. Nutrients. 2021; 13(12):4328. https://doi.org/10.3390/nu13124328
Chicago/Turabian StyleWu, Xiuzhi, Michael A. Roussell, Alison M. Hill, Penny M. Kris-Etherton, and Rosemary L. Walzem. 2021. "Baseline Insulin Resistance Is a Determinant of the Small, Dense Low-Density Lipoprotein Response to Diets Differing in Saturated Fat, Protein, and Carbohydrate Contents" Nutrients 13, no. 12: 4328. https://doi.org/10.3390/nu13124328
APA StyleWu, X., Roussell, M. A., Hill, A. M., Kris-Etherton, P. M., & Walzem, R. L. (2021). Baseline Insulin Resistance Is a Determinant of the Small, Dense Low-Density Lipoprotein Response to Diets Differing in Saturated Fat, Protein, and Carbohydrate Contents. Nutrients, 13(12), 4328. https://doi.org/10.3390/nu13124328