
Multi-Strain Probiotic Supplementation with a Product Containing Human-Native S. salivarius K12 in Healthy Adults Increases Oral S. salivarius

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Study Design
2.3. Power Analysis
2.4. Sample Collection
2.5. DNA Extraction
2.6. QPCR
2.7. 16S rRNA Gene Amplicon Sequencing Library Preparation
2.8. 16S rRNA Gene Amplicon Sequencing Data Analysis
2.9. Statistics
3. Results
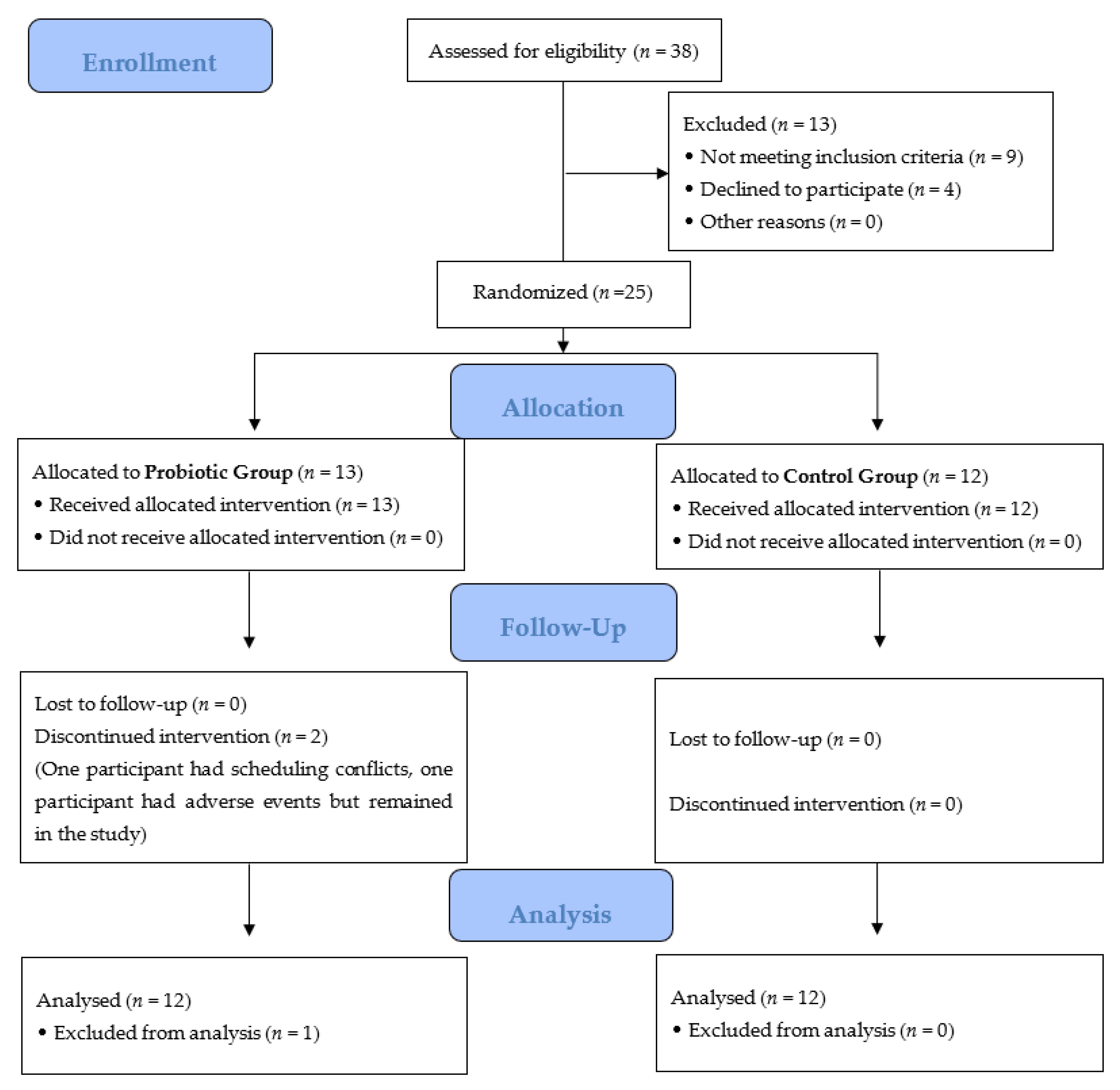
3.1. Study Participation
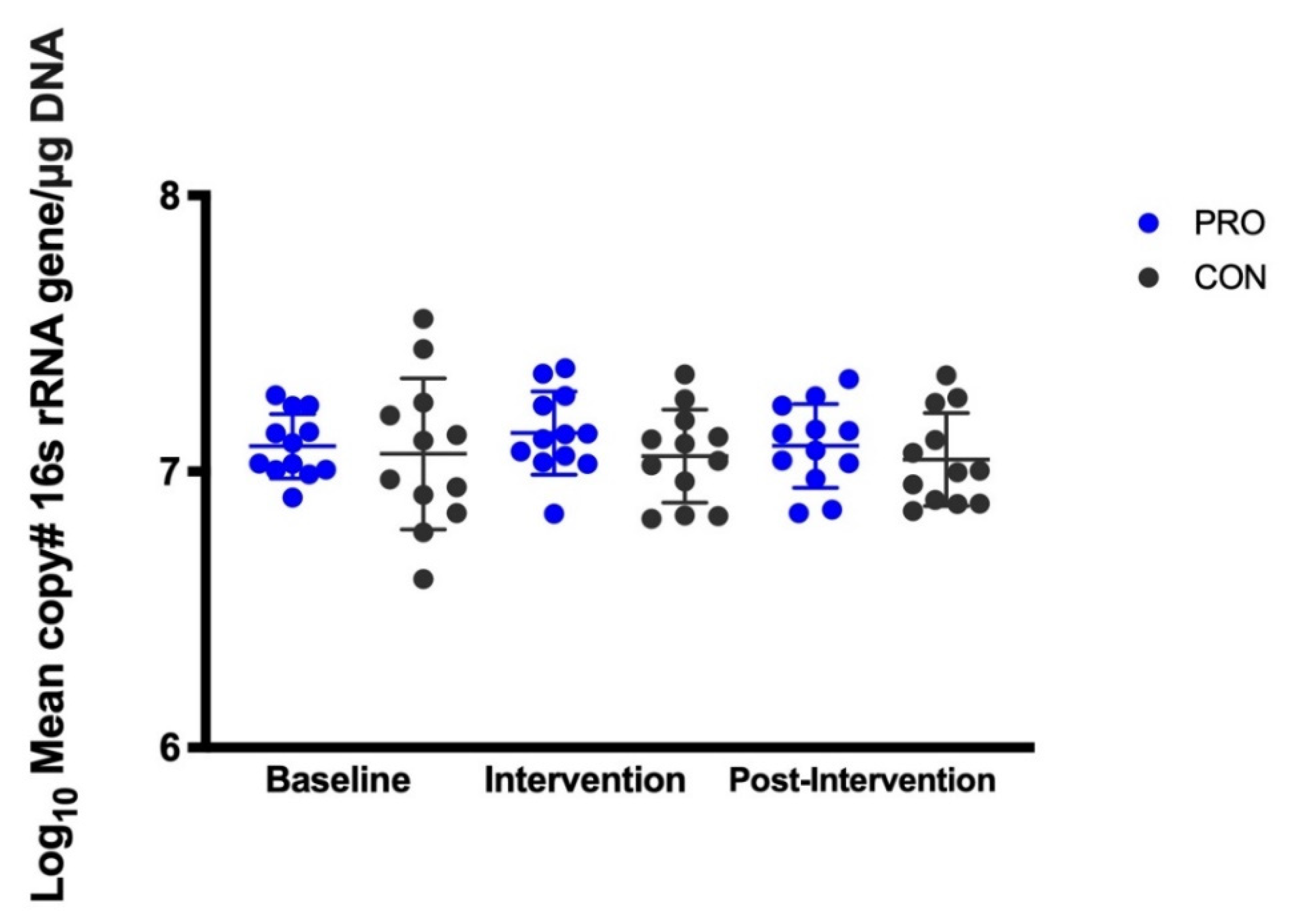
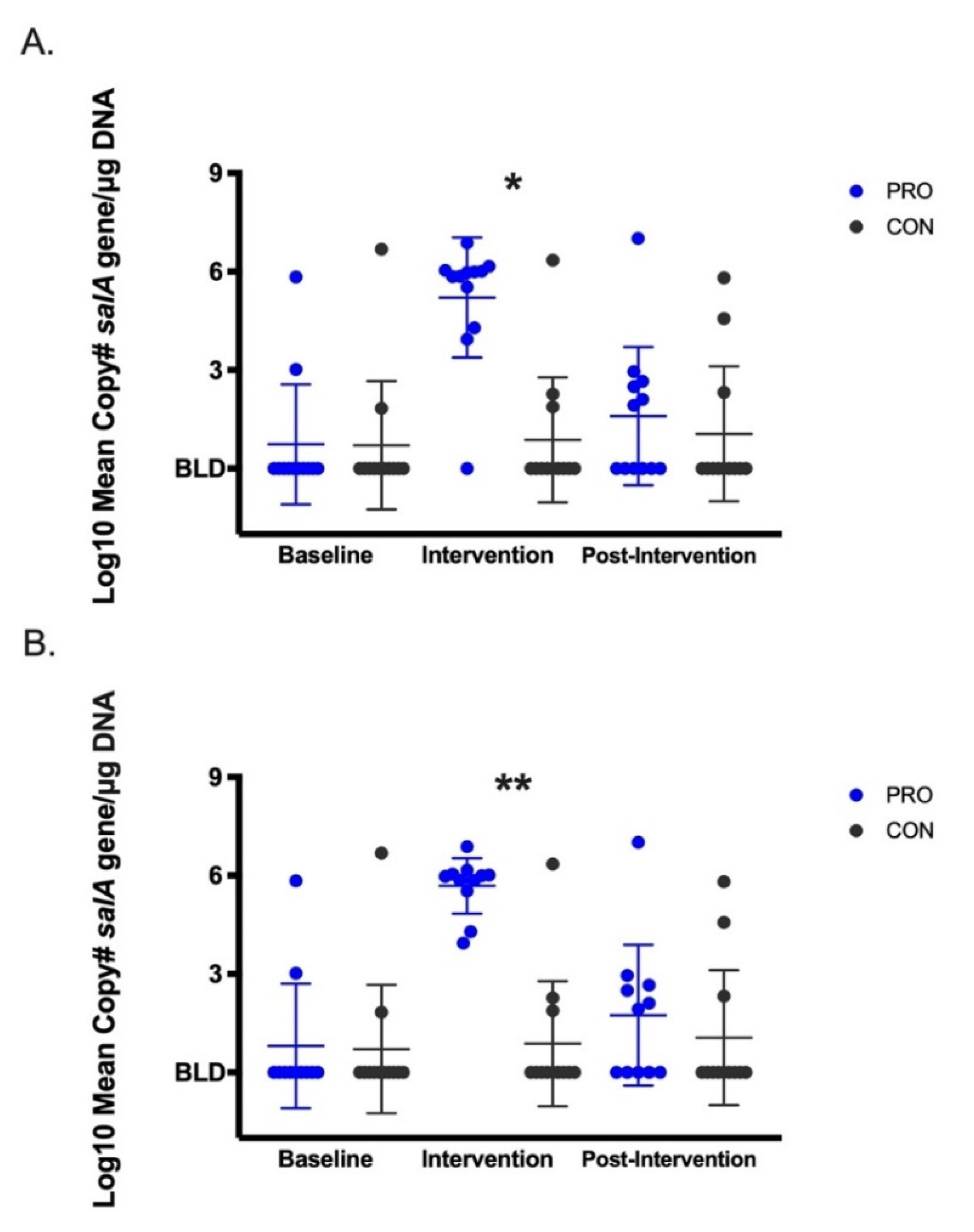
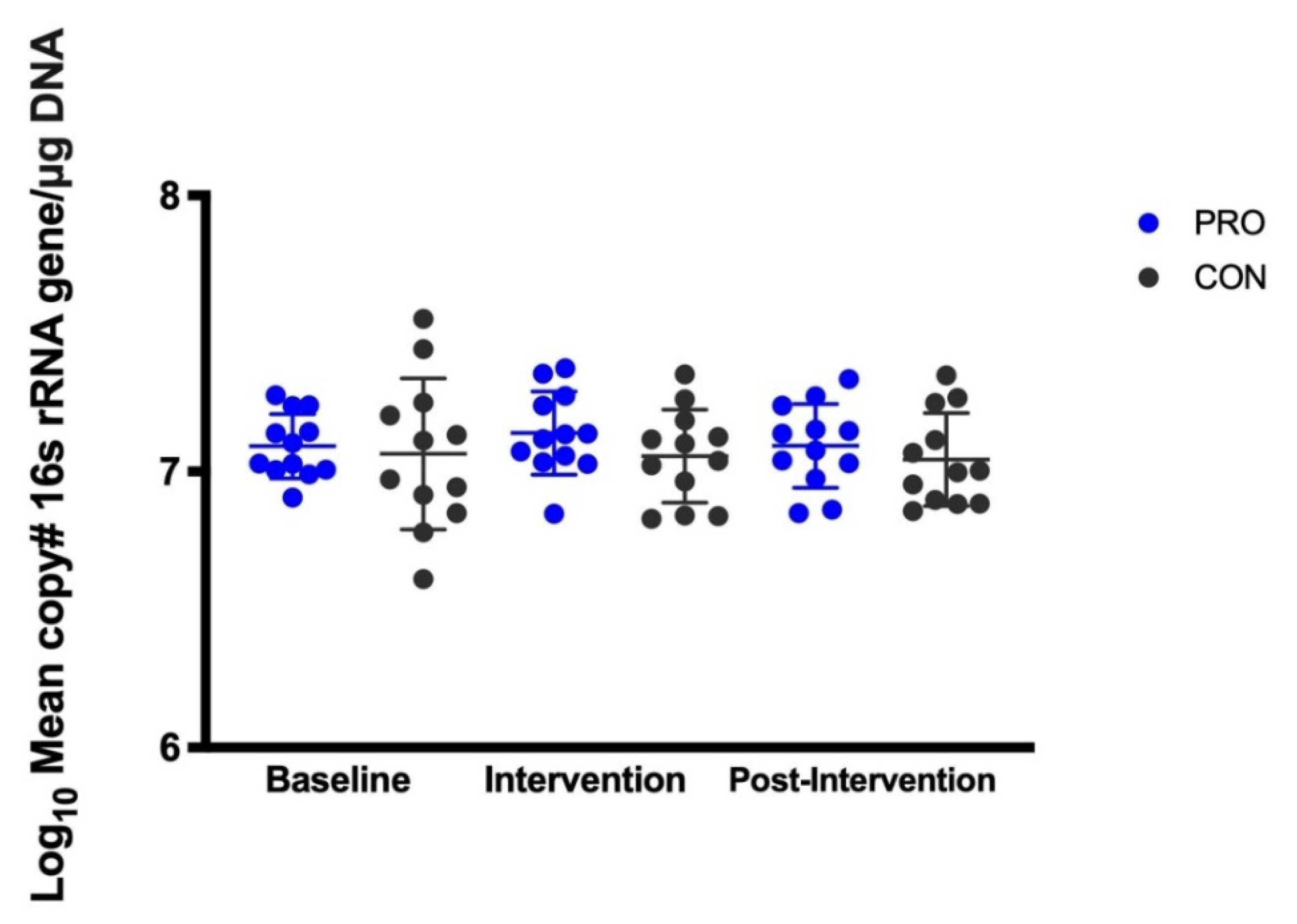
3.2. Salivary Microbes Measured by QPCR
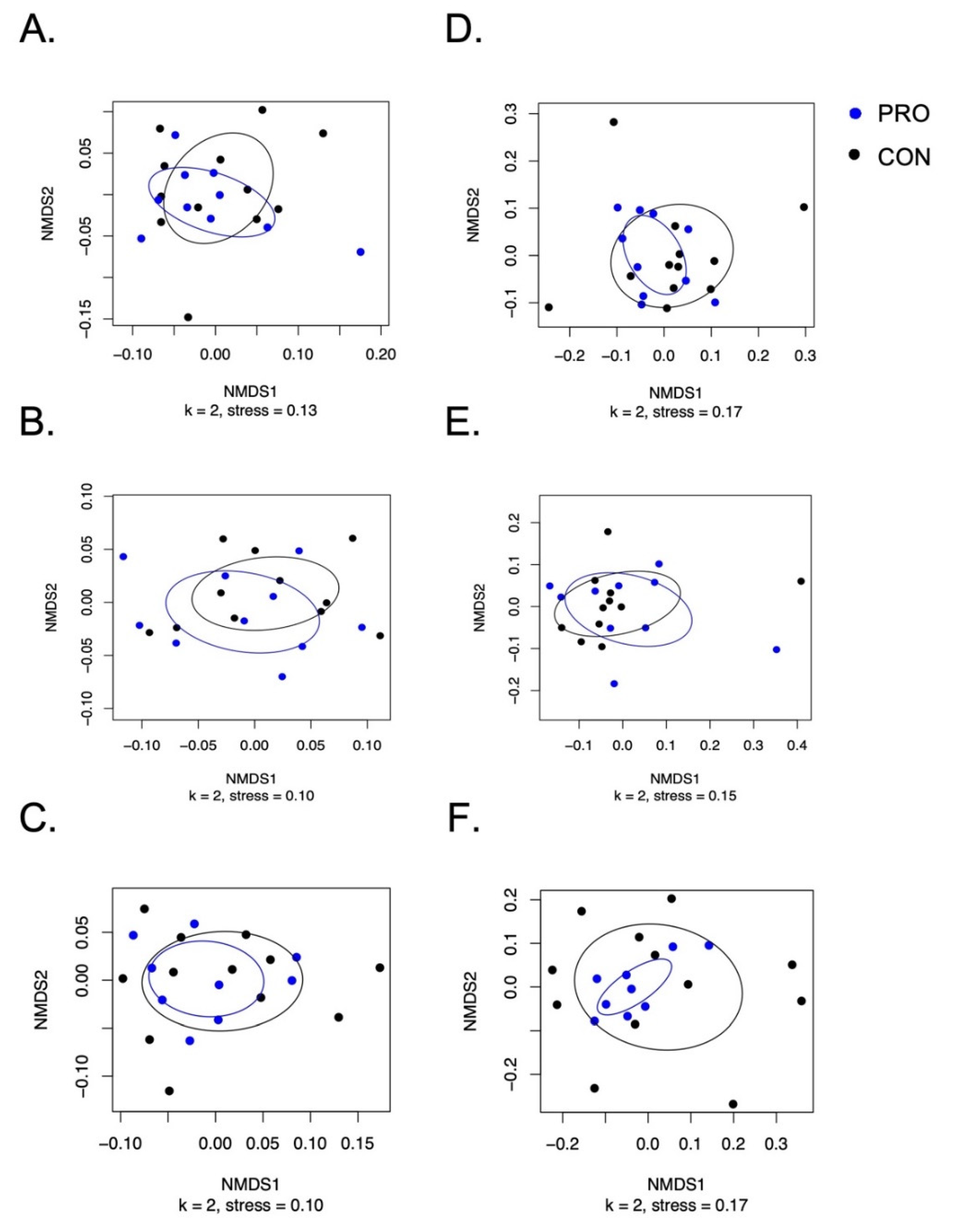
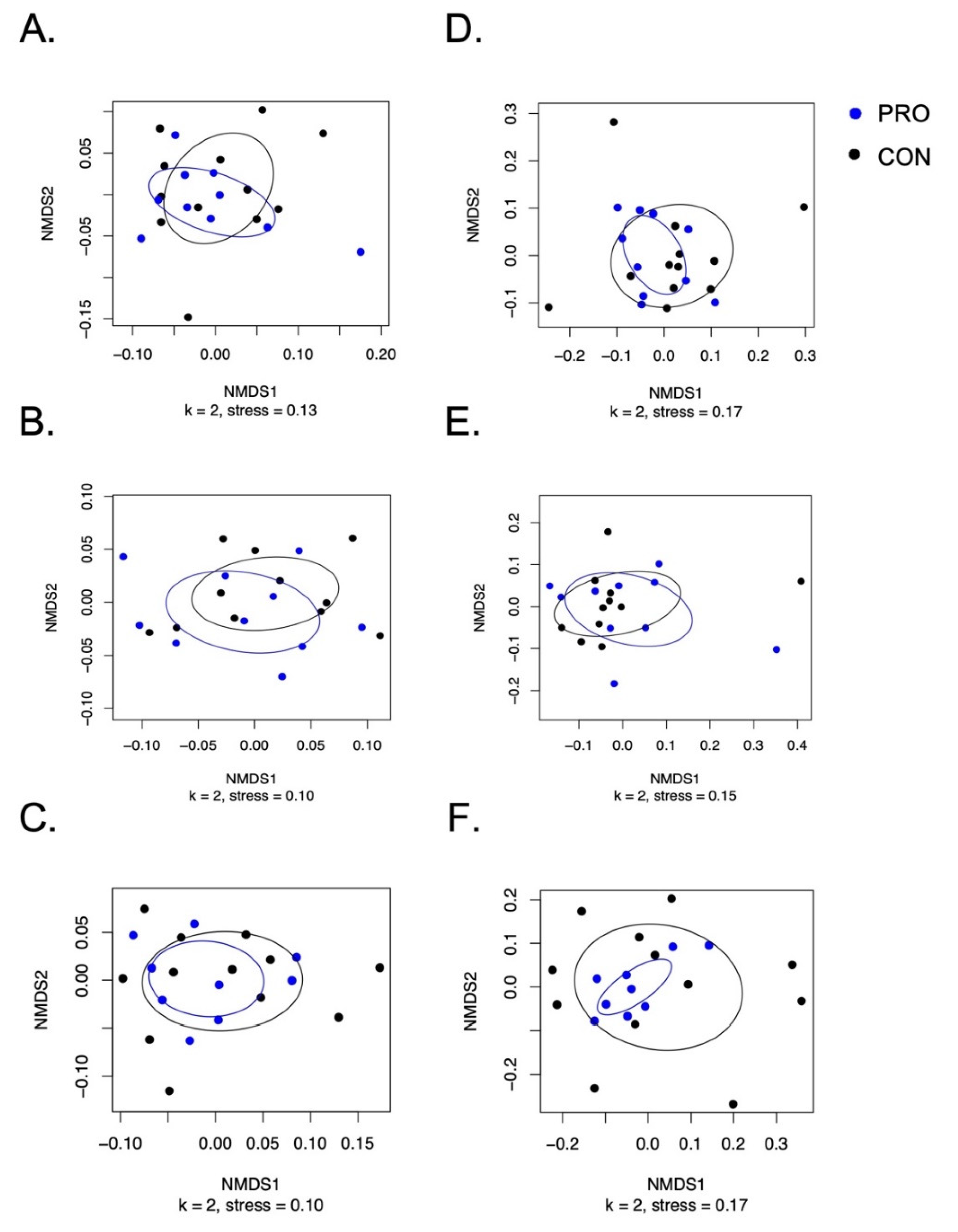
3.3. Salivary Microbiome Measured by 16S rRNA Amplicon Sequencing
3.4. Participant Health and Gastrointestinal Symptoms
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, J.P.; Wescombe, P.A.; Moore, C.J.; Chilcott, C.N.; Tagg, J.R. Safety assessment of the oral cavity probiotic Streptococcus salivarius K12. Appl. Environ. Microbiol. 2006, 72, 3050–3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horz, H.P.; Meinelt, A.; Houben, B.; Conrads, G. Distribution and persistence of probiotic Streptococcus salivarius K12 in the human oral cavity as determined by real-time quantitative polymerase chain reaction. Oral Microbiol. Immunol. 2007, 22, 126–130. [Google Scholar] [CrossRef]
- Burton, J.P.; Chilcott, C.N.; Tagg, J.R. The rationale and potential for the reduction of oral malodour using Streptococcus salivarius probiotics. Oral Dis. 2005, 11, 29–31. [Google Scholar] [CrossRef]
- Upton, M.; Tagg, J.R.; Wescombe, P.; Jenkinson, H.F. Intra- and interspecies signaling between Streptococcus salivarius and Streptococcus pyogenes mediated by SalA and SalA1 lantibiotic peptides. J. Bacteriol. 2001, 183, 3931–3938. [Google Scholar] [CrossRef] [Green Version]
- Ross, K.F.; Ronson, C.W.; Tagg, J.R. Isolation and characterization of the lantibiotic salivaricin A and its structural gene salA from Streptococcus salivarius 20P3. Appl. Environ. Microbiol. 1993, 59, 2014–2021. [Google Scholar] [CrossRef] [Green Version]
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The commensal Streptococcus salivarius K12 downregulates the innate immune responses of human epithelial cells and promotes host-microbe homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef] [Green Version]
- Zupancic, K.; Kriksic, V.; Kovacevic, I.; Kovacevic, D. Influence of Oral Probiotic Streptococcus salivarius K12 on Ear and Oral Cavity Health in Humans: Systematic Review. Probiot. Antimicrob. Proteins 2017, 9, 102–110. [Google Scholar] [CrossRef]
- Hartman, A.L.; Lough, D.M.; Barupal, D.K.; Fiehn, O.; Fishbein, T.; Zasloff, M.; Eisen, J.A. Human gut microbiome adopts an alternative state following small bowel transplantation. Proc. Natl. Acad. Sci. USA 2009, 106, 17187–17192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Joshi, N. Sabre: A Barcode Demultiplexing and Trimming Tool for FastQ Files; GitHub: San Francisco, CA, USA, 2011. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pẽa, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. Fasttree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H.; et al. Multivariable Association Discovery in Population-scale Meta-Omics Studies. Biorxiv 2021. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarlin, S.; Tejesvi, M.V.; Turunen, J.; Vänni, P.; Pokka, T.; Renko, M.; Tapiainen, T. Impact of Streptococcus salivarius K12 on Nasopharyngeal and Saliva Microbiome: A Randomized Controlled Trial. Pediatr. Infect. Dis. J. 2021, 40, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Khalesi, S.; Bellissimo, N.; Vandelanotte, C.; Williams, S.; Stanley, D.; Irwin, C. A review of probiotic supplementation in healthy adults: Helpful or hype? Eur. J. Clin. Nutr. 2019, 73, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, N.B.; Bryrup, T.; Allin, K.H.; Nielsen, T.; Hansen, T.H.; Pedersen, O. Alterations in fecal microbiota composition by probiotic supplementation in healthy adults: A systematic review of randomized controlled trials. Genome Med. 2016, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; Philippart de Foy, J.M.; Dequenne, I.; de Timary, P.; Cani, P.D. How Probiotics Affect the Microbiota. Front. Cell. Infect. Microbiol. 2020, 9, 454. [Google Scholar] [CrossRef] [Green Version]
- Frese, S.A.; Hutkins, R.W.; Walter, J.; Frese, S.A.; Hutkins, R.W.; Walter, J. Comparison of the Colonization Ability of Autochthonous and Allochthonous Strains of Lactobacilli in the Human Gastrointestinal Tract. Adv. Microbiol. 2012, 2, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Burton, J.P.; Chilcott, C.N.; Wescombe, P.A.; Tagg, J.R. Extended Safety Data for the Oral Cavity Probiotic Streptococcus salivarius K12. Probiot. Antimicrob. Proteins 2010, 2, 135–144. [Google Scholar] [CrossRef]
- Huang, F.; Teng, K.; Liu, Y.; Cao, Y.; Wang, T.; Ma, C.; Zhang, J.; Zhong, J. Bacteriocins: Potential for Human Health. Oxid. Med. Cell. Longev. 2021, 2021, 5518825. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PRO | CON | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (n = 12) | (n = 12) | |||||||||||
| Baseline | Intervention | Post-Intervention | Baseline | Intervention | Post-Intervention | |||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Number of Stools (# per day) | 1.5 | 0.55 | 1.3 | 0.51 | 1.3 | 0.56 | 1.8 | 0.82 | 1.8 | 0.85 | 1.7 | 0.78 |
| Stool Firmness (Rating 1–10) | 5.2 | 0.94 | 4.9 | 0.52 | 4.9 | 0.59 | 5.0 | 1.1 | 4.8 | 0.84 | 4.7 | 0.92 |
| Stool Consistency (Rating Type 1–7), (% Rating): | ||||||||||||
| Type 1 | 9.5% | 23.1% | 6.0% | 11.3% | 7.2% | 25.0% | 4.8% | 12.7% | 1.2% | 2.8% | 1.7% | 3.0% |
| Type 2 | 6.0% | 11.3% | 4.8% | 8.8% | 2.2% | 7.7% | 4.8% | 9.3% | 12.5% | 26.7% | 3.3% | 5.3% |
| Type 3 | 17.9% | 16.3% | 15.5% | 16.1% | 10.0% | 13.5% | 29.8% | 27.6% | 20.8% | 21.8% | 16.1% | 14.6% |
| Type 4 | 33.3% | 23.1% | 32.1% | 19.4% | 53.3% | 29.3% | 33.3% | 30.0% | 45.2% | 27.1% | 58.9% | 23.4% |
| Type 5 | 9.5% | 14.1% | 19.6% | 17.3% | 11.7% | 17.1% | 15.5% | 21.5% | 11.3% | 15.7% | 13.3% | 18.2% |
| Type 6 | 13.1% | 22.3% | 8.9% | 16.7% | 4.4% | 8.2% | 4.8% | 11.1% | 4.8% | 10.3% | 2.8% | 7.8% |
| Type 7 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.6% | 2.1% | 0.0% | 0.0% |
| NA (No stool to rate) | 10.7% | 16.3% | 11.3% | 16.5% | 11.1% | 21.3% | 7.1% | 12.9% | 3.6% | 4.8% | 3.3% | 8.3% |
| PRO | CON | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (n = 12) | (n = 12) | |||||||||||
| Baseline | Intervention | Post-Intervention | Baseline | Intervention | Post-Intervention | |||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Antibiotic Use, (% of # days) | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Illness, (% # days) | 2.4% | 5.6% | 2.4% | 6.3% | 3.3% | 8.3% | 3.6% | 8.9% | 2.4% | 6.3% | 12.8% | 22.3% |
| Illness, (# of participants) | 2 | 2 | 2 | 2 | 2 | 4 | ||||||
| PRO (n = 12) | CON (n = 12) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Baseline | Intervention | Post-Intervention | Baseline | Intervention | Post-Intervention | |||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Brush Teeth (# times/day) | 1.9 | 0.4 | 1.9 | 0.4 | 1.9 | 0.4 | 1.9 | 0.3 | 2.0 | 0.4 | 1.9 | 0.4 |
| Study Toothpaste Used, (% # days) | 98.8% | 4.1% | 99.4% | 2.1% | 92.8% | 21.0% | 98.8% | 4.1% | 100.0% | 0.0% | 100.0% | 0.0% |
| Non-Study Oral Care Products Use, (% # of days) | 0.0% | 0.0% | 0.0% | 0.0% | 6.7% | 21.1% | 1.2% | 4.1% | 0.0% | 0.0% | 0.0% | 0.0% |
| Oral Inhaler Use, (% # of days) | 8.3% | 28.9% | 5.4% | 18.6% | 5.0% | 17.3% | 1.2% | 4.1% | 0.0% | 0.0% | 0.0% | 0.0% |
| PRO (n = 12) | CON (n = 12) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Baseline | Intervention | Post-Intervention | Baseline | Intervention | Post-Intervention | |||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Non-Study Probiotics, (% # of days) | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Yogurt, Kombucha, or Kefir Use, (% # of days) | 0.0% | 0.0% | 0.6% | 2.1% | 1.7% | 4.1% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Raw Onion or Garlic, (% # of days) | 4.8% | 9.3% | 7.1% | 15.8% | 6.7% | 9.0% | 6.0% | 7.4% | 2.4% | 6.3% | 6.1% | 10.4% |
| Gum, (% # of days) | 16.7% | 33.8% | 15.5% | 26.2% | 15.6% | 24.3% | 4.8% | 12.7% | 3.6% | 10.3% | 2.8% | 7.8% |
| Sugary Candy, (% # of days) | 10.7% | 13.8% | 14.9% | 12.4% | 12.2% | 14.2% | 15.5% | 24.7% | 18.5% | 20.3% | 8.3% | 15.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cernioglo, K.; Kalanetra, K.M.; Meier, A.; Lewis, Z.T.; Underwood, M.A.; Mills, D.A.; Smilowitz, J.T. Multi-Strain Probiotic Supplementation with a Product Containing Human-Native S. salivarius K12 in Healthy Adults Increases Oral S. salivarius. Nutrients 2021, 13, 4392. https://doi.org/10.3390/nu13124392
Cernioglo K, Kalanetra KM, Meier A, Lewis ZT, Underwood MA, Mills DA, Smilowitz JT. Multi-Strain Probiotic Supplementation with a Product Containing Human-Native S. salivarius K12 in Healthy Adults Increases Oral S. salivarius. Nutrients. 2021; 13(12):4392. https://doi.org/10.3390/nu13124392
Chicago/Turabian StyleCernioglo, Karina, Karen M. Kalanetra, Anna Meier, Zachery T. Lewis, Mark A. Underwood, David A. Mills, and Jennifer T. Smilowitz. 2021. "Multi-Strain Probiotic Supplementation with a Product Containing Human-Native S. salivarius K12 in Healthy Adults Increases Oral S. salivarius" Nutrients 13, no. 12: 4392. https://doi.org/10.3390/nu13124392
APA StyleCernioglo, K., Kalanetra, K. M., Meier, A., Lewis, Z. T., Underwood, M. A., Mills, D. A., & Smilowitz, J. T. (2021). Multi-Strain Probiotic Supplementation with a Product Containing Human-Native S. salivarius K12 in Healthy Adults Increases Oral S. salivarius. Nutrients, 13(12), 4392. https://doi.org/10.3390/nu13124392