
Plasmodium chabaudi Infection Alters Intestinal Morphology and Mucosal Innate Immunity in Moderately Malnourished Mice

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice and Parasite
2.2. Malnutrition and Infection
2.3. Physiology of Malaria and Malnutrition
2.4. Preparation of Gut Tissues for Flow Cytometry
2.5. Flow Cytometry (Surface Staining)
2.6. Flow Cytometry (Intracellular Cytokine Staining)
2.7. Assessment of Gut Pathology and Leakage
2.8. Cytokine Determination by ELISA
2.9. Data Analysis
3. Results
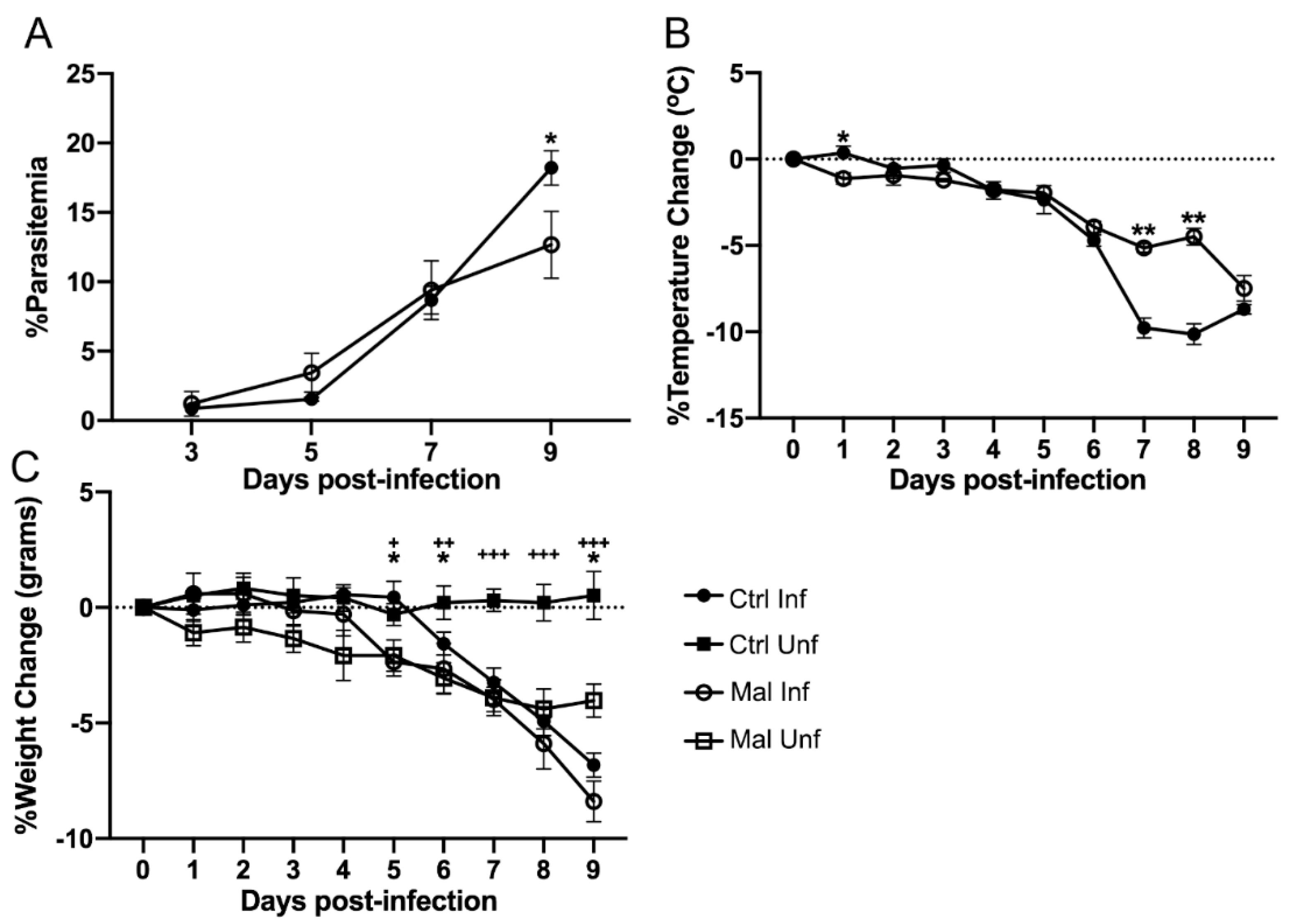
3.1. Moderate Malnutrition Does Not Exacerbate Malaria Induced Pathology
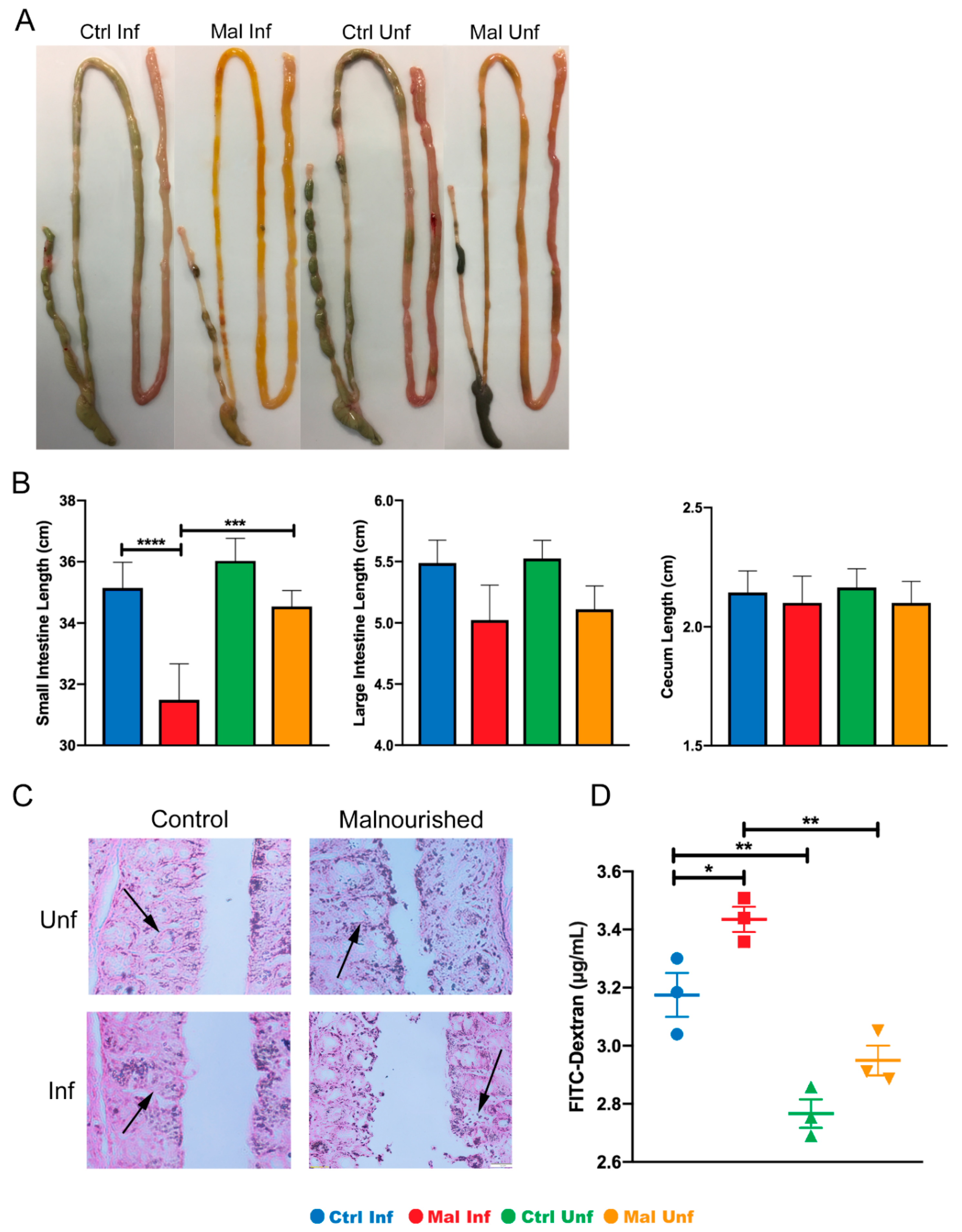
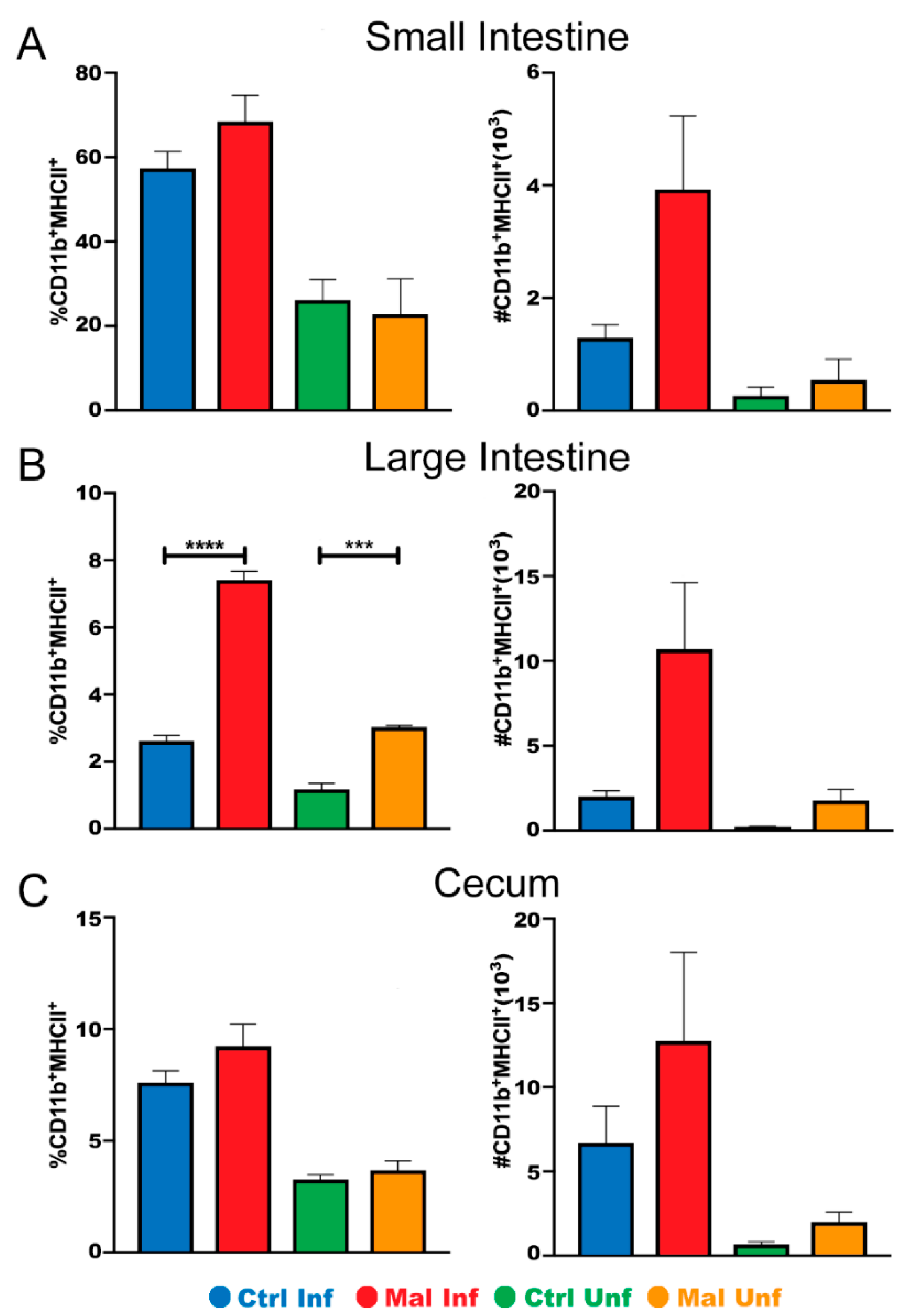
3.2. Changes in Gut Phenotype
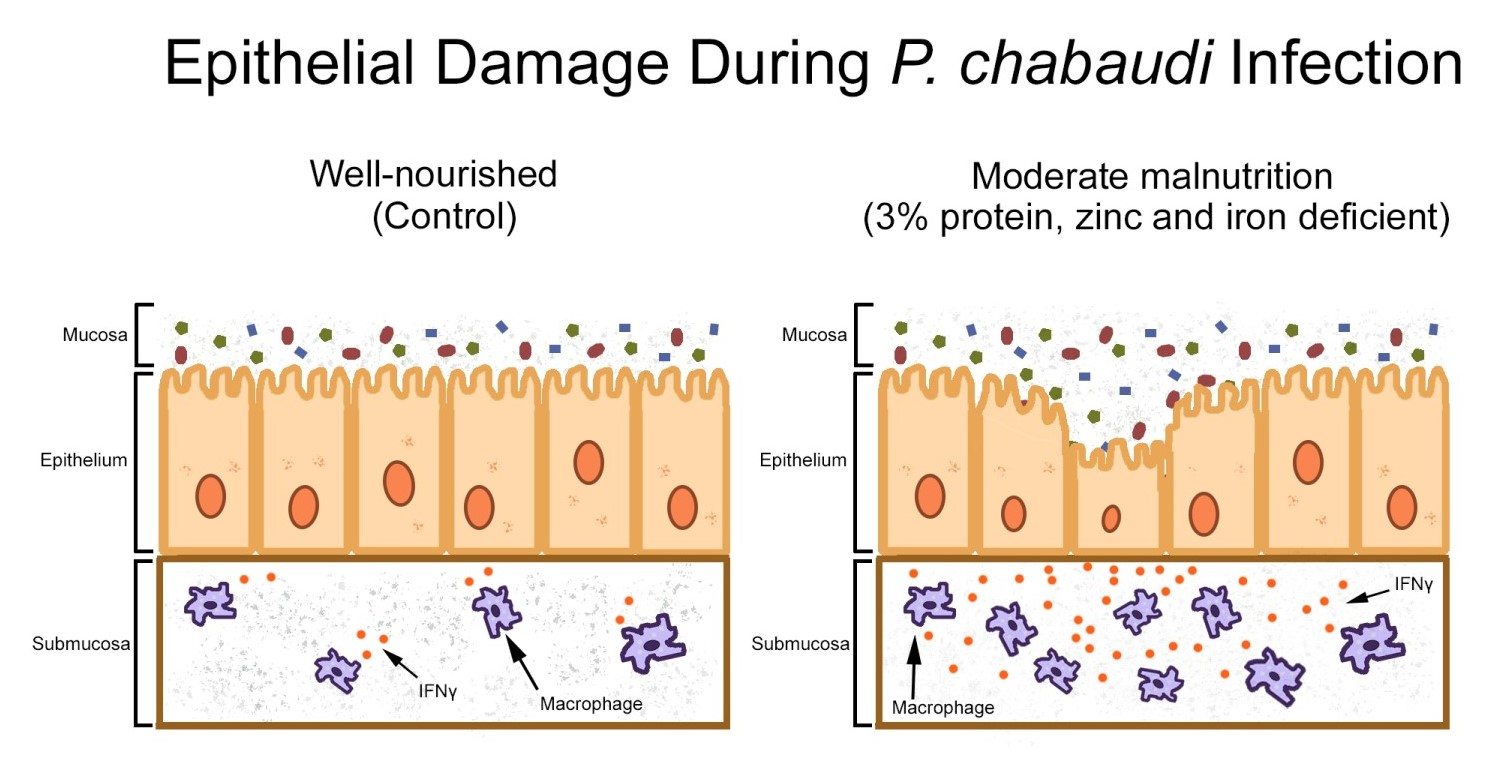
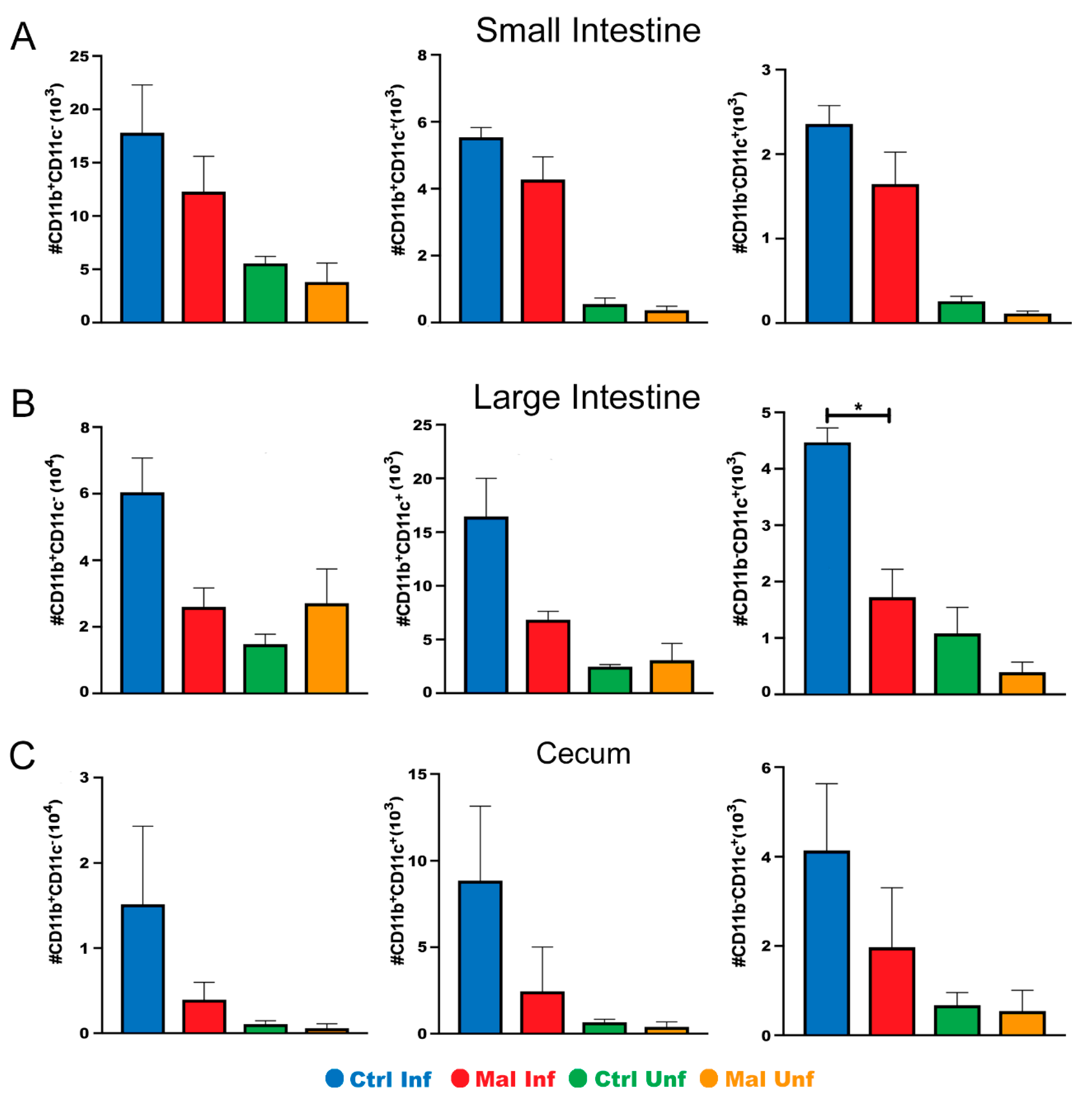
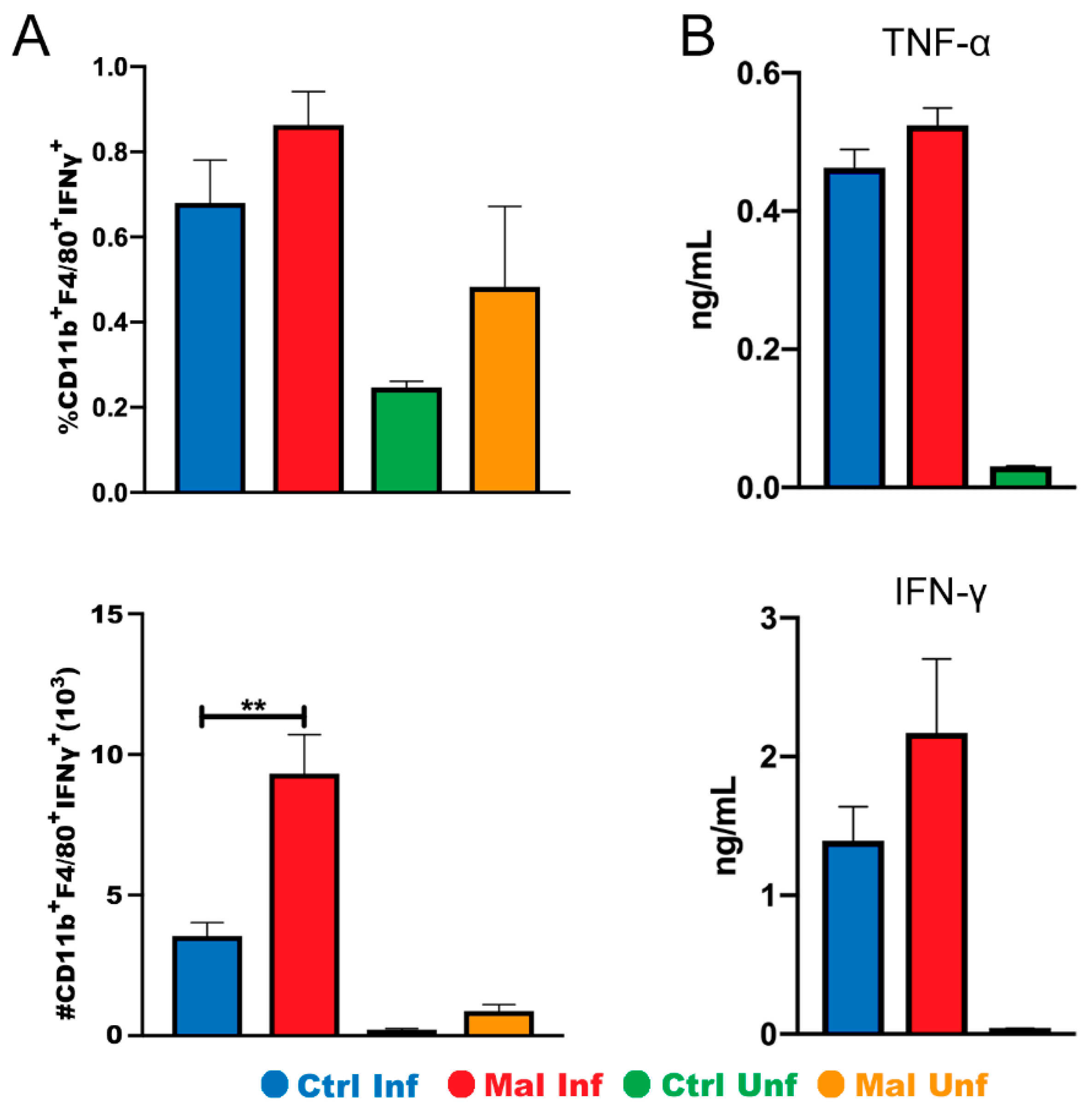
3.3. Moderate Malnutrition Alters Mucosal Innate Immunity
3.4. Moderate Malnutrition Increases Cytokine Production by Macrophages in the Small Intestine during Malaria Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Ehrhardt, S.; Burchard, G.D.; Mantel, C.; Cramer, J.P.; Kaiser, S.; Kubo, M.; Otchwemah, R.N.; Bienzle, U.; Mockenhaupt, F.P. Malaria, Anemia, and Malnutrition in African Children—Defining Intervention Priorities. J. Infect. Dis. 2006, 194, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Wajcman, H.; Galactéros, F. Glucose 6-phosphate dehydrogenase deficiency: A protection against malaria and a risk for hemolytic accidents. Comptes Rendus Biol. 2004, 327, 711–720. [Google Scholar] [CrossRef]
- Fevang, P.; Havemann, K.; Fevang, B.; Høstmark, A.T. Malaria and Malnutrition: Kwashiorkor Associated with Low Levels of Parasitaemia. Malar. Res. Treat. 2018, 2018, 7153173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snow, R.W.; Guerra, C.A.; Noor, A.M.; Myint, H.Y.; Hay, S.I. The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 2005, 434, 214–217. [Google Scholar] [CrossRef]
- Akombi, B.J.; Agho, K.E.; Merom, D.; Renzaho, A.M.; Hall, J.J. Child malnutrition in sub-Saharan Africa: A meta-analysis of demographic and health surveys (2006–2016). PLoS ONE 2017, 12, e0177338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoloni, A.; Zammarchi, L. Clinical aspects of uncomplicated and severe malaria. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, O.; Becher, H.; van Zweeden, A.B.; Ye, Y.; Diallo, D.A.; Konate, A.T.; Gbangou, A.; Kouyate, B.; Garenne, M. Effect of zinc supplementation on malaria and other causes of morbidity in west African children: Randomised double blind placebo controlled trial. BMJ 2001, 322, 1567. [Google Scholar] [CrossRef] [Green Version]
- Shankar, A.H. Nutritional Modulation of Malaria Morbidity and Mortality. J. Infect. Dis. 2000, 182, S37–S53. [Google Scholar] [CrossRef]
- Owusu-Agyei, S.; Newton, S.; Mahama, E.; Febir, L.G.; Ali, M.; Adjei, K.; Tchum, K.; Alhassan, L.; Moleah, T.; Tanumihardjo, S.A. Impact of vitamin A with zinc supplementation on malaria morbidity in Ghana. Nutr. J. 2013, 12, 131. [Google Scholar] [CrossRef] [Green Version]
- Petry, N.; Olofin, I.; Hurrell, R.F.; Boy, E.; Wirth, J.P.; Moursi, M.; Donahue Angel, M.; Rohner, F. The Proportion of Anemia Associated with Iron Deficiency in Low, Medium, and High Human Development Index Countries: A Systematic Analysis of National Surveys. Nutrients 2016, 8, 693. [Google Scholar] [CrossRef] [PubMed]
- White, N.J. Anaemia and malaria. Malar. J. 2018, 17, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barffour, M.A.; Schulze, K.J.; Coles, C.L.; Chileshe, J.; Kalungwana, N.; Arguello, M.; Siamusantu, W.; Moss, W.J.; West, K.P., Jr.; Palmer, A.C. High Iron Stores in the Low Malaria Season Increase Malaria Risk in the High Transmission Season in a Prospective Cohort of Rural Zambian Children. J. Nutr. 2017, 147, 1531–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Guideline: Daily Iron Supplementation in Infants and Children; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Neuberger, A.; Okebe, J.; Yahav, D.; Paul, M. Oral iron supplements for children in malaria-endemic areas. Cochrane Database Syst. Rev. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, M.; Farrell, A.; Biggs, B.-A.; Pasricha, S.-R. Effects of daily iron supplementation in primary-school-aged children: Systematic review and meta-analysis of randomized controlled trials. CMAJ Can. Med Assoc. J. 2013, 185, E791–E802. [Google Scholar] [CrossRef] [Green Version]
- Muriuki, J.M.; Mentzer, A.J.; Kimita, W.; Ndungu, F.M.; Macharia, A.W.; Webb, E.L.; Lule, S.A.; Morovat, A.; Hill, A.V.S.; Bejon, P.; et al. Iron Status and Associated Malaria Risk Among African Children. Clin. Infect. Dis. 2018, 68, 1807–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorat, S.; Daly, T.M.; Bergman, L.W.; Burns, J.M. Elevated Levels of the Plasmodium yoelii Homologue of Macrophage Migration Inhibitory Factor Attenuate Blood-Stage Malaria. Infect. Immun. 2010, 78, 5151–5162. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, M.M.; Riley, E.M. Innate immunity to malaria. Nat. Rev. Immunol. 2004, 4, 169–180. [Google Scholar] [CrossRef]
- Gowda, D.C.; Wu, X. Parasite Recognition and Signaling Mechanisms in Innate Immune Responses to Malaria. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.Q.; Nhi, D.M.; Huy, N.T.; Kikuchi, M.; Yanagi, T.; Hamano, S.; Hirayama, K. Splenic CD11c+ cells derived from semi-immune mice protect naïve mice against experimental cerebral malaria. Malar. J. 2015, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Oakley, M.S.; Chorazeczewski, J.K.; Aleshnick, M.; Anantharaman, V.; Majam, V.; Chawla, B.; Myers, T.G.; Su, Q.; Okoth, W.A.; Takeda, K.; et al. TCRβ-expressing macrophages induced by a pathogenic murine malaria correlate with parasite burden and enhanced phagocytic activity. PLoS ONE 2018, 13, e0201043. [Google Scholar] [CrossRef]
- Tamura, T.; Kimura, K.; Yuda, M.; Yui, K. Prevention of Experimental Cerebral Malaria by Flt3 Ligand during Infection with Plasmodium ANKA. Infect. Immun. 2011, 79, 3947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldenburg, C.E.; Guerin, P.J.; Berthé, F.; Grais, R.F.; Isanaka, S. Malaria and Nutritional Status Among Children With Severe Acute Malnutrition in Niger: A Prospective Cohort Study. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2018, 67, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812. [Google Scholar] [CrossRef]
- Esmaillzadeh, A.; Kimiagar, M.; Mehrabi, Y.; Azadbakht, L.; Hu, F.B.; Willett, W.C. Dietary patterns and markers of systemic inflammation among Iranian women. J. Nutr. 2007, 137, 992–998. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.Q.; Bretin, A.; Adeshirlarijaney, A.; Yeoh, B.S.; Vijay-Kumar, M.; Zou, J.; Denning, T.L.; Chassaing, B.; Gewirtz, A.T. “Western Diet”-Induced Adipose Inflammation Requires a Complex Gut Microbiota. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Blander, J.M.; Longman, R.S.; Iliev, I.D.; Sonnenberg, G.F.; Artis, D. Regulation of inflammation by microbiota interactions with the host. Nat. Immunol. 2017, 18, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S.; et al. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef]
- Deen, J.L.; Walraven, G.E.; von Seidlein, L. Increased risk for malaria in chronically malnourished children under 5 years of age in rural Gambia. J. Trop. Pediatr. 2002, 48, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, H.; West, C.E.; Veenemans, J.; Beguin, Y.; Kok, F.J. Stunting may determine the severity of malaria-associated anemia in African children. Pediatrics 2002, 110, e48. [Google Scholar] [CrossRef] [Green Version]
- Genton, B.; Al-Yaman, F.; Ginny, M.; Taraika, J.; Alpers, M.P. Relation of anthropometry to malaria morbidity and immunity in Papua New Guinean children. Am. J. Clin. Nutr. 1998, 68, 734–741. [Google Scholar] [CrossRef] [Green Version]
- Snow, R.W.; Byass, P.; Shenton, F.C.; Greenwood, B.M. The relationship between anthropometric measurements and measurements of iron status and susceptibility to malaria in Gambian children. Trans. R. Soc. Trop. Med. Hyg. 1991, 85, 584–589. [Google Scholar] [CrossRef]
- Taniguchi, T.; Miyauchi, E.; Nakamura, S.; Hirai, M.; Suzue, K.; Imai, T.; Nomura, T.; Handa, T.; Okada, H.; Shimokawa, C.; et al. Plasmodium berghei ANKA causes intestinal malaria associated with dysbiosis. Sci. Rep. 2015, 5, 15699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anstead, G.M.; Chandrasekar, B.; Zhao, W.; Yang, J.; Perez, L.E.; Melby, P.C. Malnutrition alters the innate immune response and increases early visceralization following Leishmania donovani infection. Infect. Immun. 2001, 69, 4709–4718. [Google Scholar] [CrossRef] [Green Version]
- Villarino, N.F.; LeCleir, G.R.; Denny, J.E.; Dearth, S.P.; Harding, C.L.; Sloan, S.S.; Gribble, J.L.; Campagna, S.R.; Wilhelm, S.W.; Schmidt, N.W. Composition of the gut microbiota modulates the severity of malaria. Proc. Natl. Acad. Sci. USA 2016, 113, 2235–2240. [Google Scholar] [CrossRef] [Green Version]
- Denning, T.L.; Norris, B.A.; Medina-Contreras, O.; Manicassamy, S.; Geem, D.; Madan, R.; Karp, C.L.; Pulendran, B. Functional Specializations of Intestinal Dendritic Cell and Macrophage Subsets That Control Th17 and Regulatory T Cell Responses Are Dependent on the T Cell/APC Ratio, Source of Mouse Strain, and Regional Localization. J. Immunol. 2011, 187, 733–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, S.; Misharin, A.; Perlman, H. A novel Ly6C/Ly6G-based strategy to analyze the mouse splenic myeloid compartment. Cytom. A 2012, 81, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Pabst, O.; Bernhardt, G. The puzzle of intestinal lamina propria dendritic cells and macrophages. Eur. J. Immunol. 2010, 40, 2107–2111. [Google Scholar] [CrossRef]
- Cerovic, V.; Houston, S.A.; Scott, C.L.; Aumeunier, A.; Yrlid, U.; Mowat, A.M.; Milling, S.W.F. Intestinal CD103− dendritic cells migrate in lymph and prime effector T cells. Mucosal Immunol. 2013, 6, 104–113. [Google Scholar] [CrossRef]
- Arnold, I.C.; Mathisen, S.; Schulthess, J.; Danne, C.; Hegazy, A.N.; Powrie, F. CD11c+ monocyte/macrophages promote chronic Helicobacter hepaticus-induced intestinal inflammation through the production of IL-23. Mucosal Immunol. 2016, 9, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Bourke, C.D.; Berkley, J.A.; Prendergast, A.J. Immune Dysfunction as a Cause and Consequence of Malnutrition. Trends Immunol. 2016, 37, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Bwakura-Dangarembizi, M.; Amadi, B.; Bourke, C.D.; Robertson, R.C.; Mwapenya, B.; Chandwe, K.; Kapoma, C.; Chifunda, K.; Majo, F.; Ngosa, D.; et al. Health Outcomes, Pathogenesis and Epidemiology of Severe Acute Malnutrition (HOPE-SAM): Rationale and methods of a longitudinal observational study. BMJ Open 2019, 9, e023077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, O.; Krawinkel, M. Malnutrition and health in developing countries. CMAJ 2005, 173, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snow, R.W.; Omumbo, J.A. Malaria. In Disease and Mortality in Sub-Saharan Africa, 2nd ed.; Jamison, D.T., Feachem, R.G., Makgoba, M.W., Bos, E.R., Baingana, F.K., Hofman, K.J., Rogo, K.O., Eds.; The International Bank of Reconstruction & Development/The World Bank: Washington, DC, USA, 2006. [Google Scholar]
- Lamikanra, A.A.; Brown, D.; Potocnik, A.; Casals-Pascual, C.; Langhorne, J.; Roberts, D.J. Malarial anemia: Of mice and men. Blood 2007, 110, 18–28. [Google Scholar] [CrossRef]
- Leopold, S.J.; Ghose, A.; Allman, E.L.; Kingston, H.W.F.; Hossain, A.; Dutta, A.K.; Plewes, K.; Chotivanich, K.; Day, N.P.J.; Tarning, J.; et al. Identifying the Components of Acidosis in Patients With Severe Plasmodium falciparum Malaria Using Metabolomics. J. Infect. Dis. 2018, 219, 1766–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.K.; Barnes, J.L.; Osorio, E.Y.; Anstead, G.M.; Jimenez, F.; Osterholzer, J.J.; Travi, B.L.; Ahuja, S.S.; White, A.C.; Melby, P.C. Deficiency of Lymph Node-Resident Dendritic Cells (DCs) and Dysregulation of DC Chemoattractants in a Malnourished Mouse Model of Leishmania donovani Infection. Infect. Immun. 2014, 82, 3098. [Google Scholar] [CrossRef] [Green Version]
- Fuss, I.J.; Strober, W.; Cuccherini, B.A.; Pearlstein, G.R.; Bossuyt, X.; Brown, M.; Fleisher, T.A.; Horgan, K. Intestinal lymphangiectasia, a disease characterized by selective loss of naive CD45RA+ lymphocytes into the gastrointestinal tract. Eur. J. Immunol. 1998, 28, 4275–4285. [Google Scholar] [CrossRef]
- Spencer, S.P.; Wilhelm, C.; Yang, Q.; Hall, J.A.; Bouladoux, N.; Boyd, A.; Nutman, T.B.; Urban, J.F.; Wang, J.; Ramalingam, T.R.; et al. Adaptation of Innate Lymphoid Cells to a Micronutrient Deficiency Promotes Type 2 Barrier Immunity. Science 2014, 343, 432–437. [Google Scholar] [CrossRef] [Green Version]
- Schuster, G.U.; Kenyon, N.J.; Stephensen, C.B. Vitamin A Deficiency Decreases and High Dietary Vitamin A Increases Disease Severity in the Mouse Model of Asthma. J. Immunol. 2008, 180, 1834–1842. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Thaiss, C.A.; Zeevi, D.; Dohnalová, L.; Zilberman-Schapira, G.; Mahdi, J.A.; David, E.; Savidor, A.; Korem, T.; Herzig, Y.; et al. Microbiota-Modulated Metabolites Shape the Intestinal Microenvironment by Regulating NLRP6 Inflammasome Signaling. Cell 2015, 163, 1428–1443. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, E.D.; Zheng, H.; Joglekar, P.; Higginbottom, S.K.; Firbank, S.J.; Bolam, D.N.; Sonnenburg, J.L. Specificity of polysaccharide use in intestinal bacteroides species determines diet-induced microbiota alterations. Cell 2010, 141, 1241–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo Hornero, R.; Hamad, I.; Côrte-Real, B.; Kleinewietfeld, M. The Impact of Dietary Components on Regulatory T Cells and Disease. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-G.; Udayanga, K.G.S.; Totsuka, N.; Weinberg, J.B.; Núñez, G.; Shibuya, A. Gut dysbiosis promotes M2 macrophage polarization and allergic airway inflammation via fungi-induced PGE2. Cell Host Microbe 2014, 15, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Ji, B.; Babaei, P.; Das, P.; Lappa, D.; Ramakrishnan, G.; Fox, T.E.; Haque, R.; Petri, W.A.; Bäckhed, F.; et al. Gut microbiota dysbiosis is associated with malnutrition and reduced plasma amino acid levels: Lessons from genome-scale metabolic modeling. Metab. Eng. 2018, 49, 128–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stough, J.M.A.; Dearth, S.P.; Denny, J.E.; LeCleir, G.R.; Schmidt, N.W.; Campagna, S.R.; Wilhelm, S.W. Functional Characteristics of the Gut Microbiome in C57BL/6 Mice Differentially Susceptible to Plasmodium yoelii. Front. Microbiol. 2016, 7, 1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, G.J.; Contijoch, E.J.; Mogno, I.; Vennaro, O.H.; Llewellyn, S.R.; Ng, R.; Li, Z.; Mortha, A.; Merad, M.; Das, A.; et al. Microbiotas from Humans with Inflammatory Bowel Disease Alter the Balance of Gut Th17 and RORγt(+) Regulatory T Cells and Exacerbate Colitis in Mice. Immunity 2019, 50, 212–224.e214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lissner, D.; Schumann, M.; Batra, A.; Kredel, L.-I.; Kühl, A.A.; Erben, U.; May, C.; Schulzke, J.-D.; Siegmund, B. Monocyte and M1 Macrophage-induced Barrier Defect Contributes to Chronic Intestinal Inflammation in IBD. Inflamm. Bowel Dis. 2015, 21, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- de Morais, M.B.; Lifschitz, C.H. Intestinal permeability to lactulose and mannitol in growing rats with iron-deficiency anemia. Biol. Trace Elem. Res. 2004, 99, 233–240. [Google Scholar] [CrossRef]
- Finamore, A.; Massimi, M.; Conti Devirgiliis, L.; Mengheri, E. Zinc Deficiency Induces Membrane Barrier Damage and Increases Neutrophil Transmigration in Caco-2 Cells. J. Nutr. 2008, 138, 1664–1670. [Google Scholar] [CrossRef] [Green Version]
- Barekatain, R.; Chrystal, P.V.; Howarth, G.S.; McLaughlan, C.J.; Gilani, S.; Nattrass, G.S. Performance, intestinal permeability, and gene expression of selected tight junction proteins in broiler chickens fed reduced protein diets supplemented with arginine, glutamine, and glycine subjected to a leaky gut model. Poult. Sci. 2019, 98, 6761–6771. [Google Scholar] [CrossRef]
- Hoopes, S.L.; Willcockson, H.H.; Caron, K.M. Characteristics of multi-organ lymphangiectasia resulting from temporal deletion of calcitonin receptor-like receptor in adult mice. PLoS ONE 2012, 7, e45261. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Cabantous, S.; Poudiougou, B.; Traore, A.; Keita, M.; Cisse, M.B.; Doumbo, O.; Dessein, A.J.; Marquet, S. Evidence That Interferon-γ Plays a Protective Role during Cerebral Malaria. J. Infect. Dis. 2005, 192, 854–860. [Google Scholar] [CrossRef] [Green Version]
- Bruewer, M.; Utech, M.; Ivanov, A.I.; Hopkins, A.M.; Parkos, C.A.; Nusrat, A. Interferon-γ induces internalization of epithelial tight junction proteins via a macropinocytosis-like process. FASEB J. 2005, 19, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Youakim, A.; Ahdieh, M. Interferon-γ decreases barrier function in T84 cells by reducing ZO-1 levels and disrupting apical actin. Am. J. Physiol. Gastrointest. Liver Physiol. 1999, 276, G1279–G1288. [Google Scholar] [CrossRef]
- McKay, D.M.; Watson, J.L.; Wang, A.; Caldwell, J.; Prescott, D.; Ceponis, P.M.J.; Di Leo, V.; Lu, J. Phosphatidylinositol 3′-Kinase Is a Critical Mediator of Interferon-γ-Induced Increases in Enteric Epithelial Permeability. J. Pharmacol. Exp. Ther. 2007, 320, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Woting, A.; Blaut, M. Small Intestinal Permeability and Gut-Transit Time Determined with Low and High Molecular Weight Fluorescein Isothiocyanate-Dextrans in C3H Mice. Nutrients 2018, 10, 685. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.M.; Wlodarska, M.; Willing, B.P.; Vonaesch, P.; Han, J.; Reynolds, L.A.; Arrieta, M.-C.; Uhrig, M.; Scholz, R.; Partida, O.; et al. Diet and specific microbial exposure trigger features of environmental enteropathy in a novel murine model. Nat. Commun. 2015, 6, 7806. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, J.P.; Mulsow, J.J.; O’Keane, C.; Docherty, N.G.; Watson, R.W.; O’Connell, P.R. Fibrogenesis in Crohn’s disease. Am. J. Gastroenterol. 2007, 102, 439–448. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15.25.11–15.25.14. [Google Scholar] [CrossRef] [PubMed]
- Cernetich, A.; Garver, L.S.; Jedlicka, A.E.; Klein, P.W.; Kumar, N.; Scott, A.L.; Klein, S.L. Involvement of gonadal steroids and gamma interferon in sex differences in response to blood-stage malaria infection. Infect. Immun. 2006, 74, 3190–3203. [Google Scholar] [CrossRef] [Green Version]
- Pathak, S.; Rege, M.; Gogtay, N.J.; Aigal, U.; Sharma, S.K.; Valecha, N.; Bhanot, G.; Kshirsagar, N.A.; Sharma, S. Age-dependent sex bias in clinical malarial disease in hypoendemic regions. PLoS ONE 2012, 7, e35592. [Google Scholar] [CrossRef] [PubMed]
- Ferruzza, S.; Scacchi, M.; Scarino, M.L.; Sambuy, Y. Iron and copper alter tight junction permeability in human intestinal Caco-2 cells by distinct mechanisms. Toxicol. Vitro 2002, 16, 399–404. [Google Scholar] [CrossRef]
- Silva, G.B.d.J.; Pinto, J.R.; Barros, E.J.G.; Farias, G.M.N.; Daher, E.D.F. Kidney involvement in malaria: An update. Rev. Inst. Med. Trop. Sao Paulo 2017, 59, e53. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murr, N.J.; Olender, T.B.; Smith, M.R.; Smith, A.S.; Pilotos, J.; Richard, L.B.; Mowa, C.N.; Opata, M.M. Plasmodium chabaudi Infection Alters Intestinal Morphology and Mucosal Innate Immunity in Moderately Malnourished Mice. Nutrients 2021, 13, 913. https://doi.org/10.3390/nu13030913
Murr NJ, Olender TB, Smith MR, Smith AS, Pilotos J, Richard LB, Mowa CN, Opata MM. Plasmodium chabaudi Infection Alters Intestinal Morphology and Mucosal Innate Immunity in Moderately Malnourished Mice. Nutrients. 2021; 13(3):913. https://doi.org/10.3390/nu13030913
Chicago/Turabian StyleMurr, Noah Joseph, Tyler B. Olender, Margaret R. Smith, Amari S. Smith, Jennifer Pilotos, Lyndsay B. Richard, Chishimba Nathan Mowa, and Michael Makokha Opata. 2021. "Plasmodium chabaudi Infection Alters Intestinal Morphology and Mucosal Innate Immunity in Moderately Malnourished Mice" Nutrients 13, no. 3: 913. https://doi.org/10.3390/nu13030913
APA StyleMurr, N. J., Olender, T. B., Smith, M. R., Smith, A. S., Pilotos, J., Richard, L. B., Mowa, C. N., & Opata, M. M. (2021). Plasmodium chabaudi Infection Alters Intestinal Morphology and Mucosal Innate Immunity in Moderately Malnourished Mice. Nutrients, 13(3), 913. https://doi.org/10.3390/nu13030913