
Combination-Feeding Causes Differences in Aspects of Systemic and Mucosal Immune Cell Phenotypes and Functions Compared to Exclusive Sow-Rearing or Formula-Feeding in Piglets

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
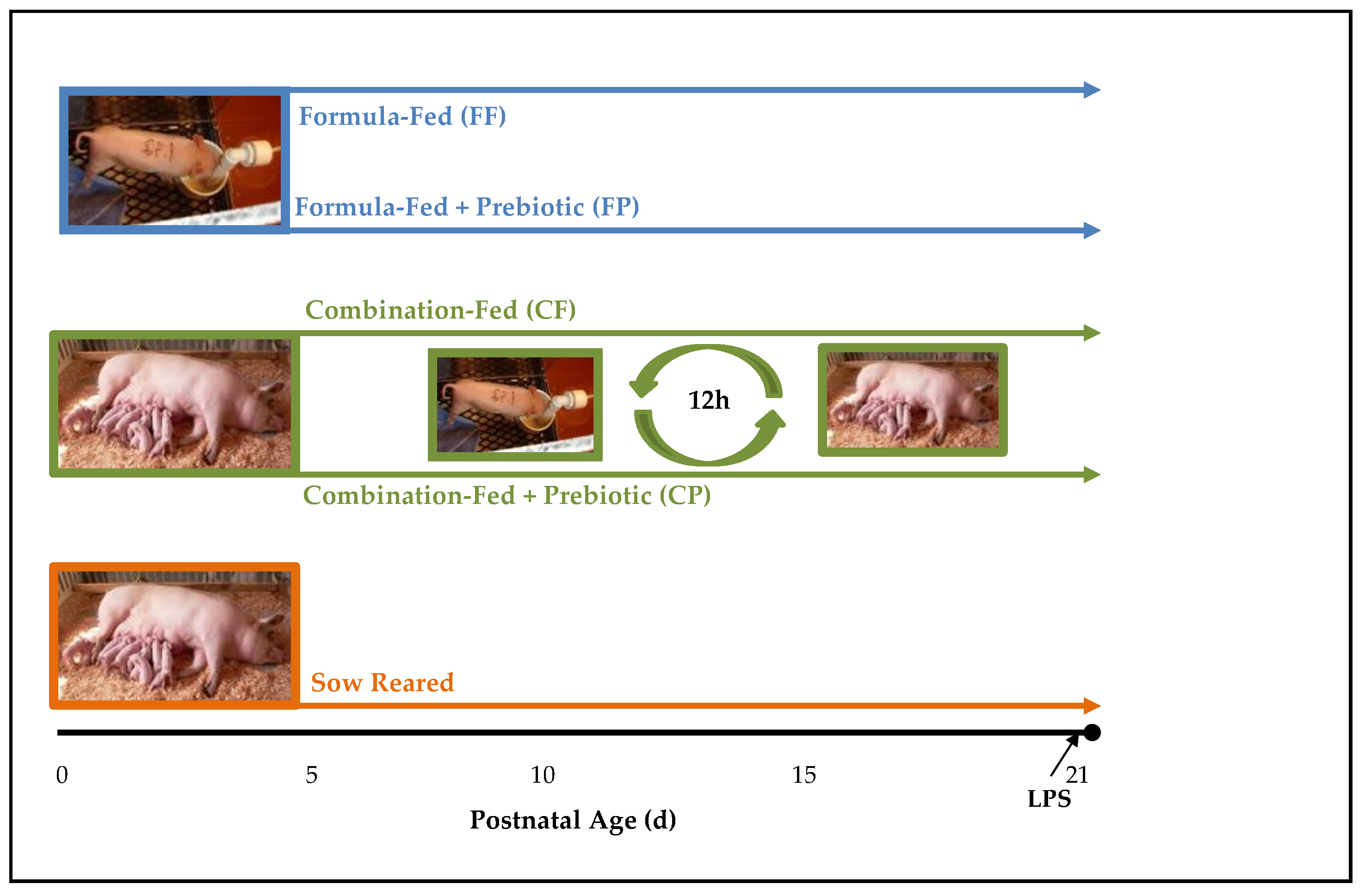
2.1. Experimental Design and Sample Collection
2.2. Isolation of Mononuclear Cells from Blood, Spleen and MLN
2.3. Enrichment of Macrophages
2.4. Phenotypic Identification of Mononuclear Cells by Flow Cytometry
2.5. Macrophage Activation Assays
2.6. Serum IL-6 and TNF-α
2.7. Statistical Analyses
3. Results
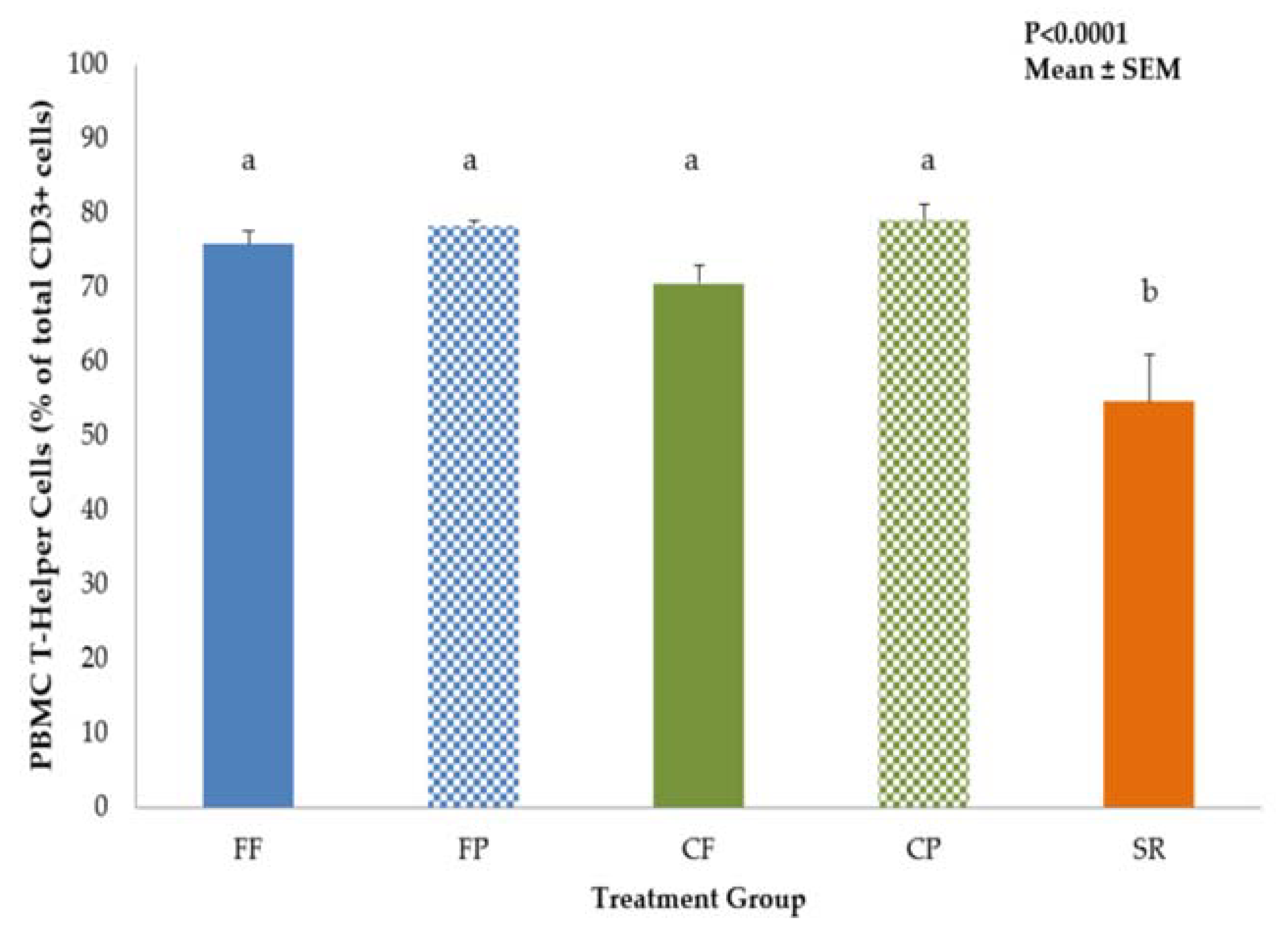
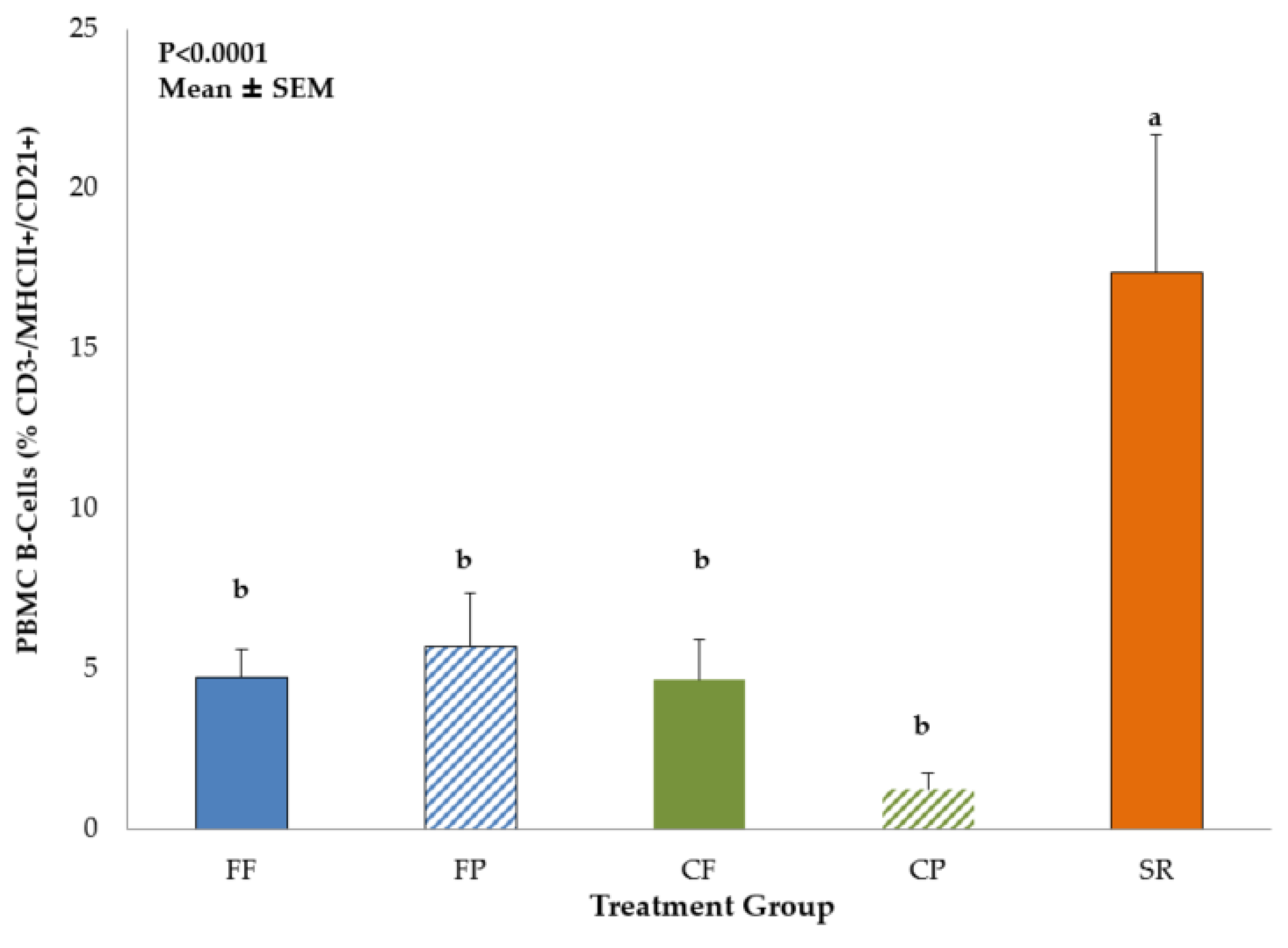
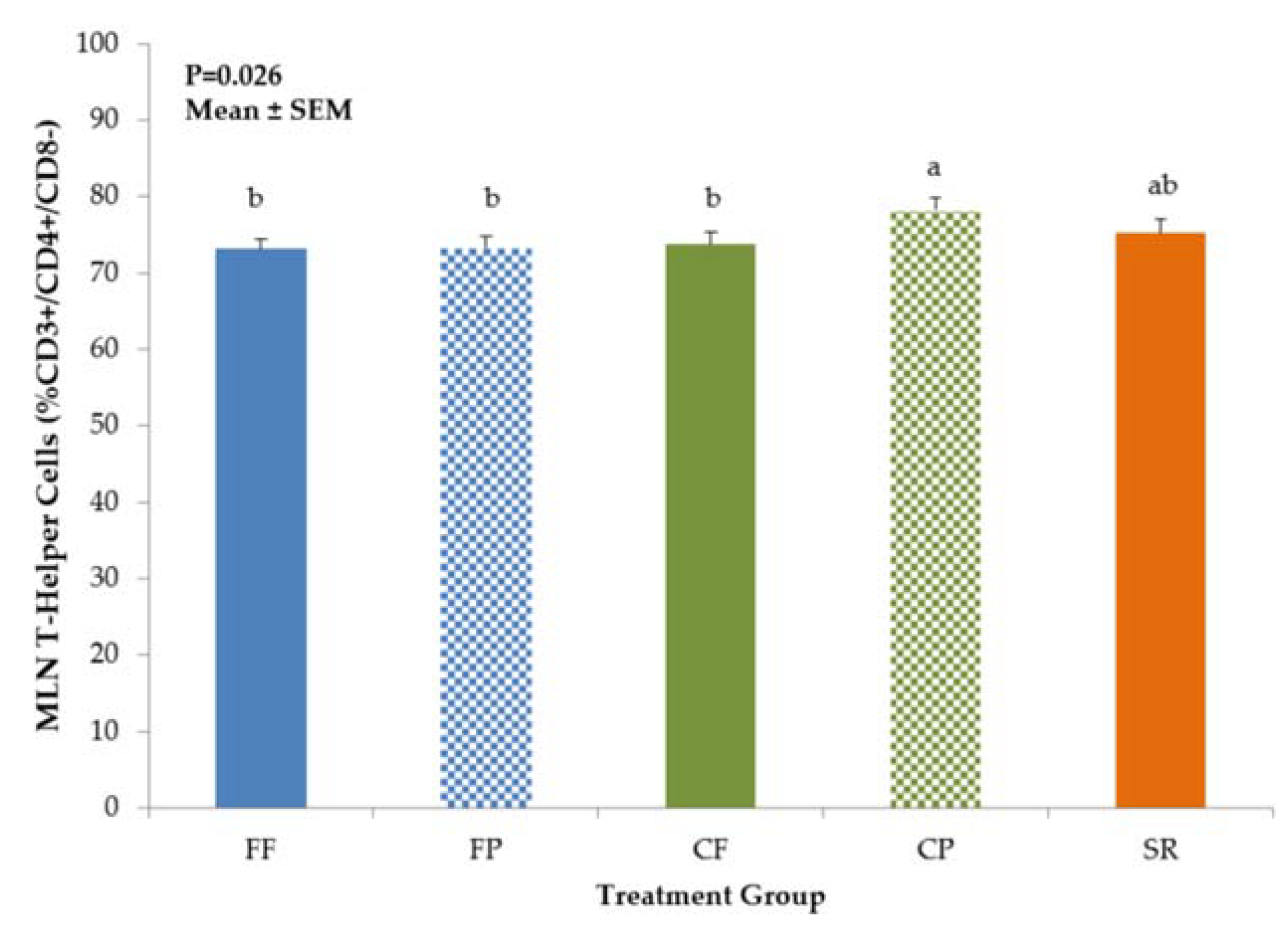
3.1. Phenotypic Identification of Mononuclear Cells by Flow Cytometry
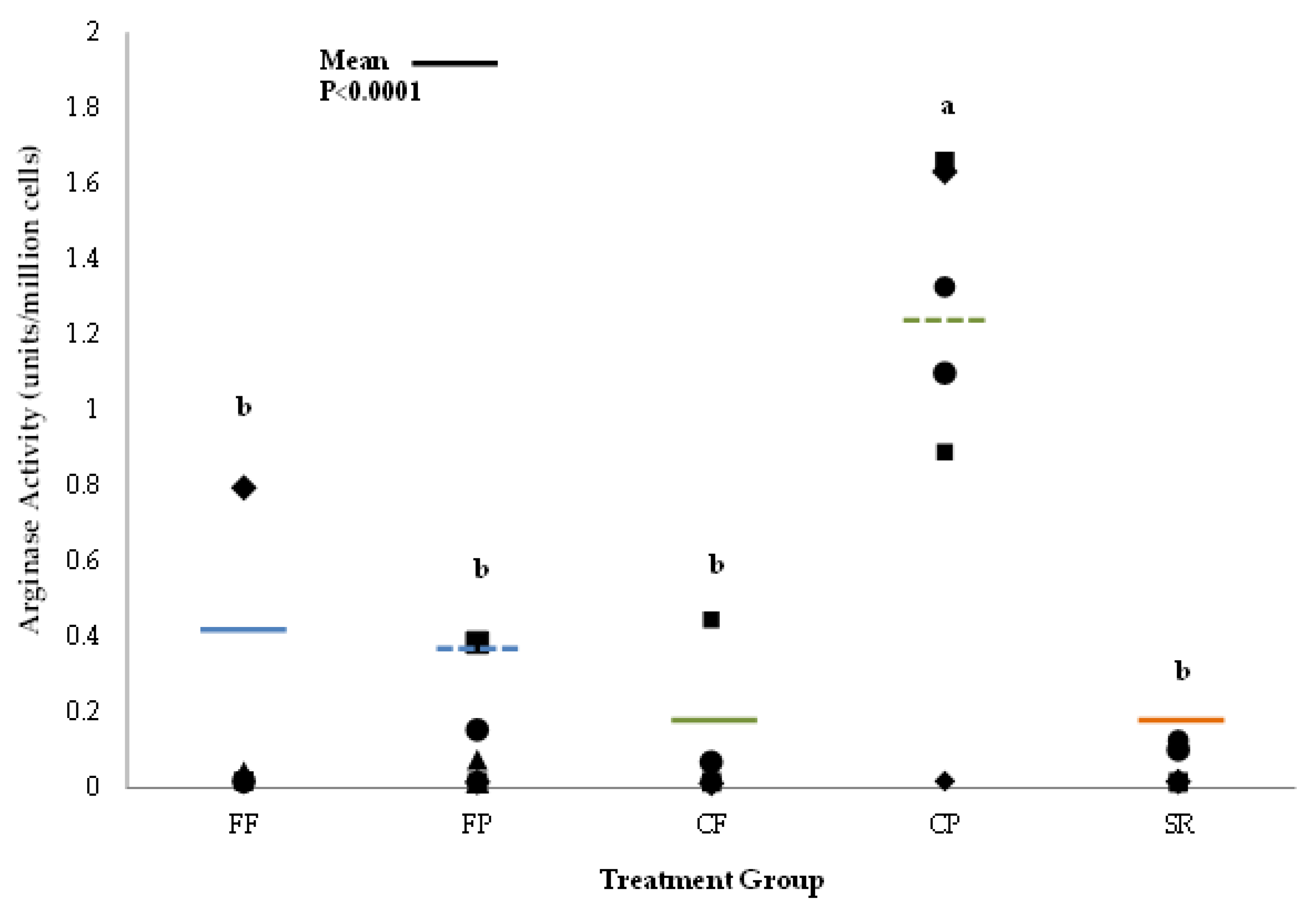
3.2. Macrophage Activation Assays
3.3. Serum IL-6 and TNF-α
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gartner, L.M.; Morton, J.; Lawrence, R.A.; Naylor, A.J.; O’Hare, D.; Schanler, R.J.; Eidelman, A.I.; American Academy of Pediatrics Section on Breastfeeding. Breastfeeding and the use of human milk. Pediatrics 2015, 115, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Victora, C.G.; Bahl, R.; Barros, A.J.D.; França, G.V.A.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C.; et al. Breastfeeding in the 21st century: Epidemiology, mechanisms and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Ip, S.; Chung, M.; Raman, G.; Trikalinos, T.A.; Lau, J. A summary of the Agency for Healthcare Research and Quality’s evidence report on breastfeeding in developed countries. Breastfeed Med. 2009, 4 (Suppl. 1), S17–S30. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Breastfeeding Report Card. 2020. Available online: https://www.cdc.gov/breastfeeding/data/reportcard.htm (accessed on 15 January 2021).
- Dinour, L.M.; Szaro, J.M. Employer-based programs to support breastfeeding among working mothers: A systematic review. Breastfeed. Med. 2017, 12, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.S.; Zhou, W.; Arensberg, M.B. The effect of employment status on breastfeeding in the United States. Womens Health Issues 2006, 16, 243–252. [Google Scholar] [CrossRef]
- Thulier, D.; Mercer, J. Variables associated with breastfeeding duration. J. Obstet. Gynecol. Neonatal Nurs. 2009, 38, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Han, D.H.; Shin, J.-M.; An, S.; Kim, J.S.; Kim, D.-Y.; Moon, S.; Kim, J.-S.; Cho, J.-S.; Kim, S.-W.; Kim, Y.H.; et al. Long-term breastfeeding in the prevention of allergic rhinitis: Allergic rhinitis cohort study for kids (ARCO-Kids study). Clin. Exp. Otorhinolaryngol. 2019, 12, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Ling, P.-R.; Blackburn, G.L. Review of infant feeding: Key features of breastmilk and infant formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkerman, R.; Fass, M.M.; de Vos, P. Non-digestible carbohydrates in infant formula as substitution for human milk oligosaccharide function: Effects on microbiota and gut maturation. Crit. Rev. Food. Sci. Nutr. 2019, 59, 1486–1497. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, M.F.; Jenmalm, M.C.; Garofalo, R.P.; Björkstén, B. Cytokines in breast milk from allergic and nonallergic mothers. Pediatr. Res. 2000, 47, 157–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, S.M.; Comstock, S.S. Human milk oligosaccharides influence neonatal mucosal and systemic immunity. Ann. Nutr. Metab. 2017, 69, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.C.; Wang, M.; Donovan, S.M. The role of early life nutrition in the establishment of gut microbial composition and function. Gut Microbes 2017, 8, 143–171. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, M.; Donovan, S.M. Early development of the gut microbiome and immune-mediated childhood disorders. Semin. Reprod. Med. 2014, 32, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Monaco, M.H.; Donovan, S.M. Impact of early gut microbiota on immune and metabolic development and function. Semin. Fetal Neonatal Med. 2016, 21, 380–387. [Google Scholar] [CrossRef]
- Karmaus, W.; Soto-Ramirez, N.; Zhang, H. Infant feeding pattern in the first six months of age in the USA: A follow-up study. Int. Breastfeed. J. 2017, 12, 48. [Google Scholar] [CrossRef]
- Garrido, D.; Dallas, D.; Mills, D. Consumption of human milk glycoconjugates by infant-associated bifidobacteria: Mechanisms and implications. Microbiology 2013, 159, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Vandenplas, Y.; Berger, B.; Paolo Carnielli, V.; Ksiazyk, J.; Lagström, H.; Sanchez Luna, M.; Migacheva, M.; Mosselmans, J.-M.; Picaud, J.-M.; Possner, M.; et al. Human milk oligosaccharides: 2-fucosyllactose (2-FL) and lacto-N-neotetraose (LNnT) in infant formula. Nutrients 2018, 10, 1161. [Google Scholar] [CrossRef] [Green Version]
- Vandenplas, Y.; De Greef, E.; Veereman, G. Prebiotics in infant formula. Gut Microbes 2014, 5, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Radlowski, E.C.; Li, M.; Monaco, M.H.; Donovan, S.M. Feeding mode, but not prebiotics, affects colonic microbiota composition and volatile fatty acid concentrations in sow-reared, formula-fed and combination-fed piglets. J. Nutr. 2019, 149, 2156–2163. [Google Scholar] [CrossRef]
- Wang, M.; Radlowski, E.C.; Monaco, M.H.; Fahey, G.C.; Gaskins, H.R.; Donovan, S.M. Mode of delivery and early nutrition modulate microbial colonization and fermentation products in the neonatal piglet. J. Nutr. 2013, 143, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Correa-Matos, N.J.; Donovan, S.M.; Isaacson, R.E.; Gaskins, H.R.; White, B.A.; Tappenden, K.A. Fermentable fiber reduces recovery time and improves intestinal function in piglets following Salmonella typhimurium infection. J. Nutr. 2003, 133, 1845–1852. [Google Scholar] [CrossRef] [Green Version]
- Webel, D.M.; Finck, B.N.; Baker, D.H.; Johnson, R.W. Time course of increased plasma cytokines, cortisol, and urea nitrogen in pigs following intraperitoneal injection of lipopolysaccharide. J. Anim. Sci. 1997, 7, 1514–1520. [Google Scholar] [CrossRef] [Green Version]
- Boudry, C.; Buldgen, A.; Portetelle, D.; Collard, A.; Théwis, A.; Dehoux, J.-P. Effects of oral supplementation with bovine colostrum on the immune system of weaned piglets. Res. Vet. Sci. 2007, 83, 91–101. [Google Scholar] [CrossRef]
- Thorum, S.C.; Hester, S.N.; Comstock, S.S.; Monaco, M.H.; Pence, B.D.; Woods, J.A.; Donovan, S.M. Dietary (1, 3/1, 6)-β-D-glucan decreases transforming growth factor β expression in the lung of the neonatal piglet. Nutr. Res. 2013, 33, 322–331. [Google Scholar] [CrossRef]
- Comstock, S.S.; Reznikov, E.A.; Contractor, N.; Donovan, S.M. Dietary bovine lactoferrin alters mucosal and systemic immune cell response in the neonatal piglet. J. Nutr. 2014, 144, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S.; Grisham, M.B. Methods to detect nitric oxide and its metabolites in biological samples. Free Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, R.; Mosser, D. The isolation and characterization of murine macrophages. Curr. Protoc. Immunol. 2015, 111, 14.1.1–14.1.16. [Google Scholar] [CrossRef]
- Monaco, M.H.; Kashtanov, D.O.; Wang, M.; Walker, D.C.; Rai, D.; Jouni, Z.E.; Miller, M.J.; Donovan, S.M. Addition of polydextrose and galactooligosaccharide to formula does not affect bacterial translocation in the neonatal piglet. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 210–216. [Google Scholar] [CrossRef]
- Sierra, C.; Bernal, M.J.; Blasco, J.; Martínez, R.; Dalmau, J.; Ortuño, I.; Espín, B.; Vasallo, M.I.; Gil, D.; Vidal, M.L.; et al. Prebiotic effect during the first year of life in healthy infants fed formula containing GOS as the only prebiotic: A multicentre, randomised, double-blind and placebo-controlled trial. Eur. J. Nutr. 2015, 54, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Ben, X.M.; Li, J.; Feng, Z.T.; Shi, S.Y.; Lu, Y.D.; Chen, R.; Zhou, X.Y. Low level of galacto-oligosaccharide in infant formula stimulates growth of intestinal bifidobacteria and lactobacilli. World J. Gastroenterol. 2008, 14, 6564–6568. [Google Scholar] [CrossRef]
- Dzidic, M.; Boix-Amorós, A.; Selma-Royo, M.; Mira, A.; Carmen Collado, M. Gut microbiota and mucosal immunity in the neonate. Med. Sci. 2018, 56, 56. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Friedberg, I.; Ivanov, I.V.; Davidson, L.A.; Goldsby, J.S.; Dahl, D.B.; Herman, D.; Wang, M.; Donovan, S.M.; Chapkin, R.S. A metagenomic study of diet-dependent interaction between gut microbiota and host in infants reveals differences in immune response. Genome Biol. 2012, 13, r32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenger, S.; Röllinghoff, M. Role of cytokines in the innate immune response to intracellular pathogens. Ann. Rheum. Dis. 2001, 60 (Suppl. 3), 43–46. [Google Scholar] [CrossRef]
- Gordon, S. Macrophage-restricted molecules: Role in differentiation and activation. Immunol. Lett. 1999, 65, 5–8. [Google Scholar] [CrossRef]
- McFadden, J.P.; Thyssen, J.P.; Basketter, D.A.; Puangpet, P.; Kimber, I. T helper cell 2 immune skewing in pregnancy/early life: Chemical exposure and the development of atopic disease and allergy. Br. J. Dermatol. 2015, 172, 584–591. [Google Scholar] [CrossRef]
- Hawkes, J.; Neumann, M.; Gibson, R. The Effect of Breast Feeding on Lymphocyte Subpopulations in Healthy Term Infants at 6 Months of Age. Pediatr. Res. 1999, 45, 648–651. [Google Scholar] [CrossRef] [PubMed]
- Andersson, Y.; Hammarström, M.L.; Lönnerdal, B.; Graverholt, G.; Fält, H.; Hernell, O. Formula feeding skews immune cell composition toward adaptive immunity compared to breastfeeding. J. Immunol. 2009, 183, 4322–4328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, M.I.; Catalan-Dibene, J.; Zlotnik, A. B cells responses and cytokine production are regulated by their immune microenvironment. Cytokine 2015, 74, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.V.; Yuan, L.; Azevedo, M.S.; Jeong, K.I.; Gonzalez, A.M.; Saif, L.J. Transfer of maternal cytokines to suckling piglets: In vivo and in vitro models with implications for immunomodulation of neonatal immunity. Vet. Immunol. Immunopathol. 2007, 117, 236–248. [Google Scholar] [CrossRef] [Green Version]
- Kainonen, E.; Rautava, S.; Isolauri, E. Immunological programming by breast milk creates an anti-inflammatory cytokine milieu in breast-fed infants compared to formula-fed infants. Br. J. Nutr. 2013, 109, 1962–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandtzaeg, P. Mucosal immunity: Integration between mother and the breast-fed infant. Vaccine 2003, 21, 3382–3388. [Google Scholar] [CrossRef]
- Tapping, R. Innate immune sensing and activation of cell surface Toll-like receptors. Semin. Immunol. 2009, 21, 175–184. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhani, S.A.; Bogue, C.W. Toll-like receptor signaling in sepsis. Curr. Opin. Pediatr. 2003, 15, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B.; Kvistgaard, A.S.; Peerson, J.M.; Donovan, S.M.; Peng, Y.M. Growth, nutrition, and cytokine response of breast-fed infants and infants fed formula with added bovine osteopontin. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Bowatte, G.; Tham, R.; Allen, K.J.; Tan, D.J.; Lau, M.; Dai, X.; Lodge, C.J. Breastfeeding and childhood acute otitis media: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Lodge, C.J.; Tan, D.J.; Lau, M.X.; Dai, X.; Tham, R.; Lowe, A.J.; Bowatte, G.; Allen, K.J.; Dharmage, S.C. Breastfeeding and asthma and allergies: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 38–53. [Google Scholar] [CrossRef]
- Sankar, M.J.; Sinha, B.; Chowdhury, R.; Bhandari, N.; Taneja, S.; Martines, J.; Bahl, R. Optimal breastfeeding practices and infant and child mortality: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 3–13. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | FF | FP | CF | CP | SR |
|---|---|---|---|---|---|
| IL-6 | 902 ± 163 a | 548 ± 114 ab | 277 ± 132 b | 542 ± 162 ab | 456 ± 103 b |
| TNF-α | 6.3 ± 0.9 | 5.5 ± 0.9 | 3.8 ± 0.3 | 6.6 ± 1.2 | 4.5 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radlowski, E.C.; Wang, M.; Monaco, M.H.; Comstock, S.S.; Donovan, S.M. Combination-Feeding Causes Differences in Aspects of Systemic and Mucosal Immune Cell Phenotypes and Functions Compared to Exclusive Sow-Rearing or Formula-Feeding in Piglets. Nutrients 2021, 13, 1097. https://doi.org/10.3390/nu13041097
Radlowski EC, Wang M, Monaco MH, Comstock SS, Donovan SM. Combination-Feeding Causes Differences in Aspects of Systemic and Mucosal Immune Cell Phenotypes and Functions Compared to Exclusive Sow-Rearing or Formula-Feeding in Piglets. Nutrients. 2021; 13(4):1097. https://doi.org/10.3390/nu13041097
Chicago/Turabian StyleRadlowski, Emily C., Mei Wang, Marcia H. Monaco, Sarah S. Comstock, and Sharon M. Donovan. 2021. "Combination-Feeding Causes Differences in Aspects of Systemic and Mucosal Immune Cell Phenotypes and Functions Compared to Exclusive Sow-Rearing or Formula-Feeding in Piglets" Nutrients 13, no. 4: 1097. https://doi.org/10.3390/nu13041097