
Physiological and Immune Functions of Punicalagin



Abstract
:1. Introduction
2. Methods
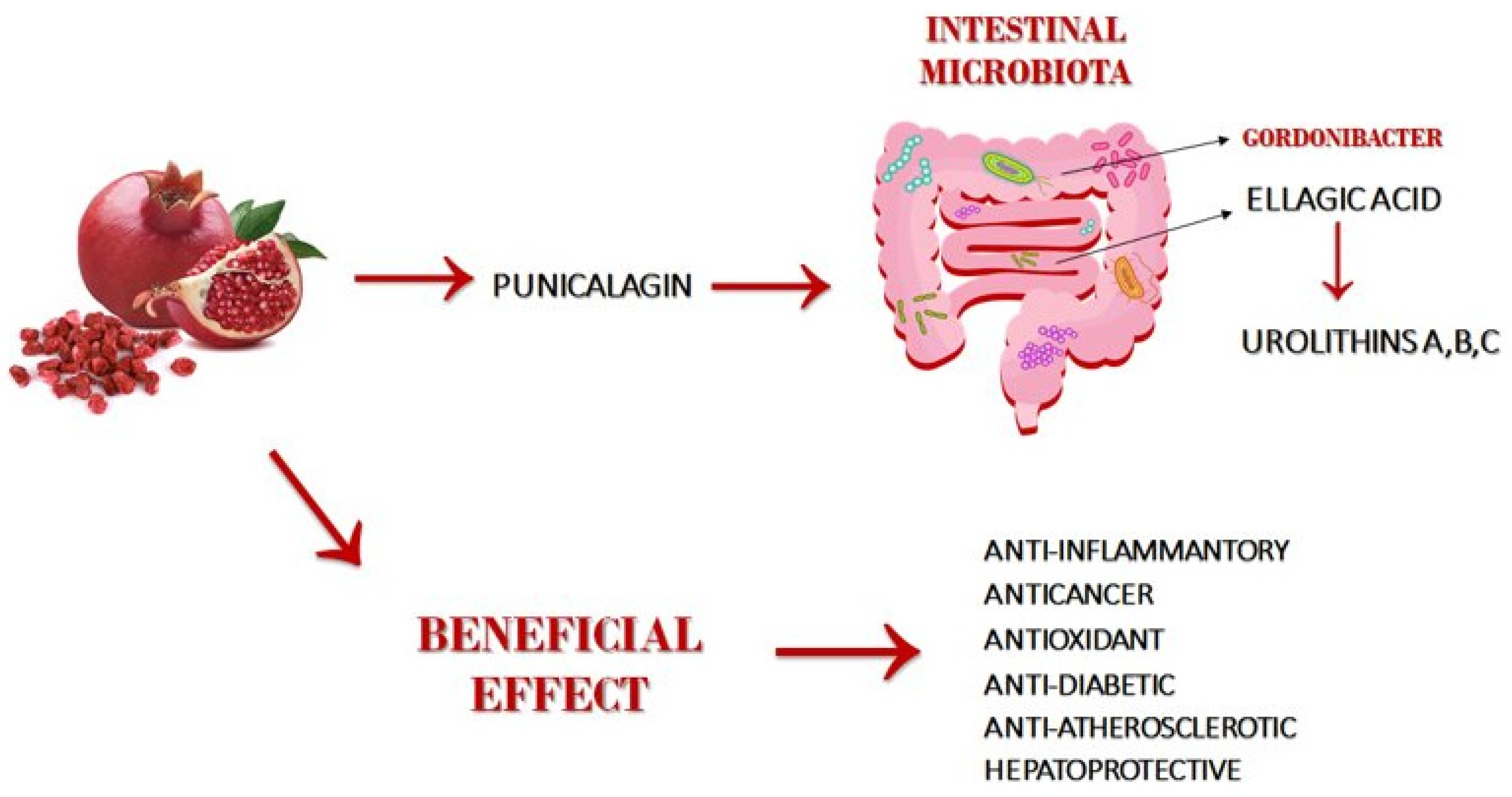
3. Metabolism and Bioavailability of Punicalagin
4. Methods of Determination PUN and EA
5. Anti-Inflammatory and Immunosuppressive Effect
5.1. NF-kB and MAPK Activation Inhibitor
5.2. Nuclear Factor of Activated T-Cells Activation Inhibitor
5.3. Apoptosis, Proliferation and Angiogenesis
5.4. Autophagy
6. Pharmacological Effect of Punicalagin and Metabolites
6.1. Antibacterial Effect
6.2. Antiviral Effect
6.3. Antioxidant Activity—Oxidative Stress
6.4. Hepatoprotective Activity
6.5. Anti-Diabetic and Anti-Obesity Activity
6.6. Anti-Atherosclerotic Activity
7. Toxicological Findings and Genotoxicity
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jurenka, J.S. Therapeutic applications of pomegranate (Punica granatum L.): A review. Altern. Med. Rev. A J. Clin. 2008, 13, 128–144. [Google Scholar]
- Abdollahzadeh, S.; Mashouf, R.; Mortazavi, H.; Moghaddam, M.; Roozbahani, N.; Vahedi, M. Antibacterial and antifungal activ-ities of punica granatum peel extracts against oral pathogens. J. Dent. 2011, 8, 1–6. [Google Scholar]
- Syed, D.N.; Chamcheu, J.-C.; Adhami, V.M.; Mukhtar, H. Pomegranate extracts and cancer prevention: Molecular and cellular activities. Anti-Cancer Agents Med. Chem. 2013, 13, 1149–1161. [Google Scholar] [CrossRef] [Green Version]
- Paller, C.J.; Pantuck, A.; Carducci, M.A. A review of pomegranate in prostate cancer. Prostate Cancer Prostatic Dis. 2017, 20, 265–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic compounds as beneficial phytochemicals in pomegranate (Punica granatum L.) peel: A review. Food Chem. 2018, 261, 75–86. [Google Scholar] [CrossRef]
- Kraszni, M.; Marosi, A.; Larive, C.K. NMR assignments and the acid–base characterization of the pomegranate ellagitannin punicalagin in the acidic pH-range. Anal. Bioanal. Chem. 2013, 405, 5807–5816. [Google Scholar] [CrossRef]
- Gil, M.I.; Tomás-Barberán, F.A.; Hess-Pierce, B.; Holcroft, D.M.; Kader, A.A. Antioxidant Activity of Pomegranate Juice and Its Relationship with Phenolic Composition and Processing. J. Agric. Food Chem. 2000, 48, 4581–4589. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Adams, L.S.; Henning, S.M.; Niu, Y.; Zhang, Y.; Nair, M.G.; Heber, D. In vitro antiproliferative, apoptotic and antioxi-dant activities of punicalagin, ellagic acid and a total pomegranate tannin extract are enhanced in combination with oth-er polyphenols as found in pomegranate juice. J. Nutr. Biochem. 2005, 16, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Oudane, B.; Boudemagh, D.; Bounekhel, M.; Sobhi, W.; Vidal, M.; Broussy, S. Isolation, characterization, antioxidant activity, and protein-precipitating capacity of the hydrolyzable tannin punicalagin from pomegranate yellow peel (Punica granatum). J. Mol. Struct. 2018, 1156, 390–396. [Google Scholar] [CrossRef]
- Moilanen, J.; Karonen, M.; Tähtinen, P.; Jacquet, R.; Quideau, S.; Salminen, J.-P. Biological activity of ellagitannins: Effects as an-ti-oxidants, pro-oxidants and metal chelators. Phytochemistry 2016, 125, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.P.; Mahal, H.; Kapoor, S.; Aradhya, S. In vitro studies on the binding, antioxidant, and cytotoxic actions of puni-calagin. J. Agric. Food Chem. 2007, 55, 1491–1500. [Google Scholar] [CrossRef]
- Tang, J.; Li, B.; Hong, S.; Liu, C.; Min, J.; Hu, M.; Li, Y.; Liu, Y.; Hong, L. Punicalagin suppresses the proliferation and invasion of cervical cancer cells through inhibition of the β-catenin pathway. Mol. Med. Rep. 2017, 16, 1439–1444. [Google Scholar] [CrossRef] [Green Version]
- Bialonska, D.; Ramnani, P.; Kasimsetty, S.G.; Muntha, K.R.; Gibson, G.R.; Ferreira, D. The influence of pomegranate by-product and punicalagins on selected groups of human intestinal microbiota. Int. J. Food Microbiol. 2010, 140, 175–182. [Google Scholar] [CrossRef]
- Lin, L.-T.; Chen, T.-Y.; Lin, S.-C.; Chung, C.-Y.; Lin, T.-C.; Wang, G.-H.; Anderson, R.; Lin, C.-C.; Richardson, C.D. Broad-spectrum antiviral activity of chebulagic acid and punicalagin against viruses that use glycosaminoglycans for entry. BMC Microbiol. 2013, 13, 187. [Google Scholar] [CrossRef] [Green Version]
- Zahin, M.; Ahmad, I.; Gupta, R.C.; Aqil, F. Punicalagin and Ellagic Acid Demonstrate Antimutagenic Activity and Inhibition of Benzo[a]pyrene Induced DNA Adducts. Biomed. Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Heber, D. Pomegranate Ellagitannins. In Herbal Medicine: Biomolecular and Clinical Aspects; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011. [Google Scholar]
- Espín, J.C.; Larrosa, M.; García-Conesa, M.T.; Tomás-Barberán, F. Biological Significance of Urolithins, the Gut Microbial Ellagic Acid-Derived Metabolites: The Evidence So Far. Evid. Based Complementary Altern. Med. 2013, 2013, 270418. [Google Scholar] [CrossRef] [Green Version]
- Silacci, P.; Tretola, M. Pomegranate’s Ellagitannins: Metabolism and Mechanisms of Health Promoting Properties. Nutr. Food Sci. Int. J. 2019, 9, 555766. [Google Scholar] [CrossRef]
- García-Villalba, R.; Vissenaekens, H.; Pitart, J.; Vaquero, M.R.; Espín, J.C.; Grootaert, C.; Selma, M.V.; Raes, K.; Smagghe, G.; Possemiers, S.; et al. Gastrointestinal Simulation Model TWIN-SHIME Shows Differences between Human Urolithin-Metabotypes in Gut Microbiota Composition, Pomegranate Polyphenol Metabolism, and Transport along the Intestinal Tract. J. Agric. Food Chem. 2017, 65, 5480–5493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, Á.M.; Tomé-Carneiro, J.; Zafrilla, P.; Mulero, J.; Tomás-Barberán, F.A.; Espín, J.C. Identifying the limits for ellagic acid bioavailability: A crossover pharmacokinetic study in healthy volunteers after consumption of pomegranate extracts. J. Funct. Foods 2015, 19, 225–235. [Google Scholar] [CrossRef]
- Selma, M.V.; Tomas-Barberan, F.A.; Beltran, D.; García-Villalba, R.; Espín, J.C. Gordonibacter urolithinfaciens sp. nov., a uro-lithin-producing bacterium isolated from the human gut. Int. J. Syst. Evol. Microbiol. 2014, 64, 2346–2352. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Beltrán, D.; García-Villalba, R.; Espín, J.C.; Tomás-Barberán, F.A. Description of urolithin production capacity from ellagic acid of two human intestinal Gordonibacter species. Food Funct. 2014, 5, 1779–1784. [Google Scholar] [CrossRef] [Green Version]
- Vaquero, M.R.; García-Villalba, R.; González-Sarrías, A.; Beltrán, D.; Tomás-Barberán, F.A.; Espín, J.C.; Selma, M.V. Interindividual variability in the human metabolism of ellagic acid: Contribution of Gordonibacter to urolithin production. J. Funct. Foods 2015, 17, 785–791. [Google Scholar] [CrossRef]
- Selma, M.V.; Romo-Vaquero, M.; García-Villalba, R.; González-Sarrías, A.; Tomás-Barberán, F.A.; Espín, J.C. The human gut microbial ecology associated with overweight and obesity determines ellagic acid metabolism. Food Funct. 2015, 7, 1769–1774. [Google Scholar] [CrossRef]
- Qin, G.; Xu, C.; Ming, R.; Tang, H.; Guyot, R.; Kramer, E.M.; Hu, Y.; Yi, X.; Qi, Y.; Xu, X.; et al. The pomegranate (Punica granatum L.) genome and the genomics of punicalagin biosynthesis. Plant J. 2017, 91, 1108–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorach, R.; Cerdá, B.; Cerón, J.J.; Espín, J.C.; Tomás-Barberán, F.A. Evaluation of the bioavailability and metabolism in the rat of punicalagin, an antioxidant polyphenol from pomegranate juice. Eur. J. Nutr. 2003, 42, 18–28. [Google Scholar] [CrossRef]
- Cerdá, B.; Espín, J.C.; Parra, S.; Martínez, P.; Tomás-Barberán, F.A. The potent in vitro antioxidant ellagitannins from pomegranate juice are metabolised into bioavailable but poor antioxidant hydroxy–6H–dibenzopyran–6–one derivatives by the colon-ic microflora of healthy humans. Eur. J. Nutr. 2004, 43, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Zuccari, G.; Baldassari, S.; Ailuno, G.; Turrini, F.; Alfei, S.; Caviglioli, G. Formulation Strategies to Improve Oral Bioavailability of Ellagic Acid. Appl. Sci. 2020, 10, 3353. [Google Scholar] [CrossRef]
- Nyamba, I.; Lechanteur, A.; Semdé, R.; Evrard, B. Physical formulation approaches for improving aqueous solubility and bio-availability of ellagic acid: A review. Eur. J. Pharm. Biopharm. 2020, 159, 198–210. [Google Scholar] [CrossRef]
- Williams, H.D.; Trevaskis, N.L.; Charman, S.; Shanker, R.M.; Charman, W.; Pouton, C.; Porter, C.J.H. Strategies to Address Low Drug Solubility in Discovery and Development. Pharm. Rev. 2013, 65, 315–499. [Google Scholar] [CrossRef]
- Qu, W.; Iii, A.P.B.; Pan, Z.; Ma, H. Quantitative determination of major polyphenol constituents in pomegranate products. Food Chem. 2012, 132, 1585–1591. [Google Scholar] [CrossRef]
- Nathan, C. Points of control in inflammation. Nat. Cell Biol. 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Xu, X.; Yin, P.; Wan, C.; Chong, X.; Liu, M.; Cheng, P.; Chen, J.; Liu, F.; Xu, J. Punicalagin inhibits inflammation in LPS-induced RAW264.7 macrophages via the suppression of TLR4-mediated MAPKs and NF-κB activation. Inflammation 2014, 37, 956–965. [Google Scholar] [CrossRef] [PubMed]
- BenSaad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and puni-calagin A&B isolated from Punica granatum. BMC Complementary Altern. Med. 2017, 17, 47. [Google Scholar]
- Lee, S.-I.; Kim, B.-S.; Kim, K.-S.; Lee, S.; Shin, K.-S.; Lim, J.-S. Immune-suppressive activity of punicalagin via inhibition of NFAT activation. Biochem. Biophys. Res. Commun. 2008, 371, 799–803. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin Prevents Inflammation in LPS-Induced RAW264.7 Macro-phages by Inhibiting FoxO3a/Autophagy Signaling Pathway. Nutrients 2019, 11, 2794. [Google Scholar] [CrossRef] [Green Version]
- Ngkelo, A.; Meja, K.; Yeadon, M.; Adcock, I.; Kirkham, P.A. LPS induced inflammatory responses in human peripheral blood mononuclear cells is mediated through NOX4 and Giα dependent PI-3kinase signalling. J. Inflamm. 2012, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Trevillyan, J.M.; Chiou, X.G.; Chen, Y.-W.; Ballaron, S.J.; Sheets, M.P.; Smith, M.L.; Wiedeman, P.E.; Warrior, U.; Wilkins, J.; Gubbins, E.J.; et al. Potent Inhibition of NFAT Activation and T Cell Cytokine Production by Novel Low Molecular Weight Pyrazole Compounds. J. Biol. Chem. 2001, 276, 48118–48126. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, H.R.; Arastoo, M.; Ostad, S.N. A Comprehensive Review of Punica granatum (Pomegranate) Properties in Toxicolog-ical, Pharmacological, Cellular and Molecular Biology Researches. Iran J. Pharm. Res. 2012, 11, 385–400. [Google Scholar] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chinnathambi, A.; Alharbi, S.A.; Veeraraghavan, V.P.; Mohan, S.K.; Zhang, G. Punicalagin promotes the apoptosis in human cervical cancer (ME-180) cells through mitochondrial pathway and by inhibiting the NF-kB signaling pathway. Saudi J. Biol. Sci. 2020, 27, 1100–1106. [Google Scholar] [CrossRef]
- Carneiro, C.C.; Santos, S.; Lino, R.D.S.; Bara, M.T.F.; Chaibub, B.A.; Reis, P.R.D.M.; Chaves, D.A.; da Silva, A.J.R.; Silva, L.S.; Silva, D.D.M.E.; et al. Chemopreventive effect and angiogenic activity of punicalagin isolated from leaves of Lafoensia pacari A. St.-Hil. Toxicol. Appl. Pharm. 2016, 310, 1–8. [Google Scholar] [CrossRef]
- Stahlhut, C.; Slack, F.J. MicroRNAs and the cancer phenotype: Profiling, signatures and clinical implications. Genome Med. 2013, 5, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Zhang, X.; Wang, H. Punicalagin inhibited proliferation, invasion and angiogenesis of osteosarcoma through suppression of NF-κB signaling. Mol. Med. Rep. 2020, 22, 2386–2394. [Google Scholar] [CrossRef]
- Syed, D.N.; Malik, A.; Hadi, N.; Sarfaraz, S.; Afaq, F.; Mukhtar, H. Photochemopreventive Effect of Pomegranate Fruit Extract on UVA-mediated Activation of Cellular Pathways in Normal Human Epidermal Keratinocytes. Photochem. Photobiol. 2006, 82, 398–405. [Google Scholar] [CrossRef]
- Larrosa, M.; Tomás-Barberán, F.A.; Espín, J.C. The dietary hydrolysable tannin punicalagin releases ellagic acid that induces apoptosis in human colon adenocarcinoma Caco-2 cells by using the mitochondrial pathway. J. Nutr. Biochem. 2006, 17, 611–625. [Google Scholar] [CrossRef]
- Wang, S.G.; Huang, M.H.; Li, J.H.; Lai, F.I.; Lee, H.M.; Hsu, Y.N. Punicalagin induces apoptotic and autophagic cell death in human U87MG glioma cells. Acta Pharm. Sin. 2013, 34, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Shao, S.H.; Xu, Z.-X.; Hennessy, B.; Ding, Z.; Larrea, M.; Kondo, S.; Dumont, D.J.; Gutterman, J.U.; Walker, C.L.; et al. The energy sensing LKB1–AMPK pathway regulates p27kip1 phosphorylation mediating the decision to enter autophagy or apoptosis. Nat. Cell Biol. 2007, 9, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Ammar, O.M.A.; Ilktac, M.; Gülcan, H. Urolithins and their antimicrobial activity: A short review. EMU J. Pharm. Sci. 2019, 3, 117–124. [Google Scholar]
- Gulube, Z.; Patel, M. Effect of Punica granatum on the virulence factors of cariogenic bacteria Streptococcus mutans. Microb. Pathog. 2016, 98, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Tito, A.; Colantuono, A.; Pirone, L.; Pedone, E.; Intartaglia, D.; Giamundo, G.; Conte, I.; Vitaglione, P.; Apone, F. A pomegranate peel extract as inhibitor of SARS-CoV-2 Spike binding to human ACE2 (in vitro): A promising source of novel antiviral drugs. Front. Chem. 2021, 9, 638187. [Google Scholar]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Djedjibegovic, J.; Marjanovic, A.; Panieri, E.; Saso, L. Ellagic Acid-Derived Urolithins as Modulators of Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 5194508. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; DELLA Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.Q.; Xin, T.A.O.; Men, X.M.; Xu, Z.W.; Tian, W.A.N.G. In vitro and in vivo antioxidant activities of three major polyphenolic com-pounds in pomegranate peel: Ellagic acid, punicalin, and punicalagin. J. Integr. Agric. 2017, 16, 1808–1818. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, H.; Liang, H.; Yuan, Q. Purification, antioxidant activity and protein-precipitating capacity of punicalin from pomegranate husk. Food Chem. 2013, 138, 437–443. [Google Scholar] [CrossRef]
- Bialonska, D.; Kasimsetty, S.G.; Khan, S.I.; Ferreira, D. Urolithins, Intestinal Microbial Metabolites of Pomegranate Ellagitannins, Exhibit Potent Antioxidant Activity in a Cell-Based Assay. J. Agric. Food Chem. 2009, 57, 10181–10186. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.A.; Qutub, H.O.; Al-Melhim, W.N. Punicalagin alleviates hepatotoxicity in rats challenged with cyclophosphamide. Environ. Toxicol. Pharmacol. 2016, 45, 158–162. [Google Scholar] [CrossRef]
- Luedde, T.; Schwabe, R.F. NF-κB in the liver—linking injury, fibrosis and hepatocellular carcinoma. Nat. Rev. Gastro-Enterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Foroutanfar, A.; Mehri, S.; Marzyeh, K.; Tandisehpanah, Z.; Hosseinzadeh, H. Protective effect of punicalagin, the main poly-phenol compound of pomegranate, against acrylamide-induced neurotoxicity and hepatotoxicity in rats. Phytother. Res. 2020, 34, 3262–3272. [Google Scholar] [CrossRef]
- Les, F.; Arbonés-Mainar, J.M.; Valero, M.S.; López, V. Pomegranate polyphenols and urolithin A inhibit α-glucosidase, dipeptidyl peptidase-4, lipase, triglyceride accumulation and adipogenesis related genes in 3T3-L1 adipocyte-like cells. J. Ethnopharmacol. 2018, 220, 67–74. [Google Scholar] [CrossRef]
- Wu, D.; Ma, X.; Tian, W. Pomegranate husk extract, punicalagin and ellagic acid inhibit fatty acid synthase and adipogenesis of 3T3-L1 adipocyte. J. Funct. Foods 2013, 5, 633–641. [Google Scholar] [CrossRef]
- Reguero, M.; Gómez de Cedrón, M.; Reglero, G.; Quintela, J.C.; de Molina, A.R. Natural Extracts to Augment Energy Ex-penditure as a Complementary Approach to Tackle Obesity and Associated Metabolic Alterations. Biomolecules 2021, 11, 412. [Google Scholar] [CrossRef]
- Mele, L.; Mena, P.; Piemontese, A.; Marino, V.; López-Gutiérrez, N.; Bernini, F.; Brighenti, F.; Zanotti, I.; Del Rio, D. Antiatherogenic effects of ellagic acid and urolithins in vitro. Arch. Biochem. Biophys. 2016, 599, 42–50. [Google Scholar] [CrossRef]
- Kruth, S.H. Fluid-phase pinocytosis of LDL by macrophages: A novel target to reduce macrophage cholesterol accumula-tion in atherosclerotic lesions. Curr. Pharm. Des. 2013, 19, 5865–5872. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, L.; Haller, V.; Ritsch, A. A Novel Candidate for Prevention and Treatment of Atherosclerosis: Urolithin B Decreases Lipid Plaque Deposition in apoE(-/-) Mice and Increases Early Stages of Reverse Cholesterol Transport in ox-LDL Treated Macrophages Cells. Mol. Nutr. Food Res. 2019, 63, e1800887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, G.-H.; Chen, W.-Q.; Shen, Z.-Y. Urolithin A shows anti-atherosclerotic activity via activation of class B scavenger receptor and activation of Nef2 signaling pathway. Pharm. Rep. 2018, 70, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Cerón, J.J.; Tomás-Barberán, F.A.; Espín, J.C. Repeated oral administration of high doses of the pomegranate ellag-itannin punicalagin to rats for 37 days is not toxic. J. Agric. Food Chem 2003, 51, 3493–3501. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Lamar, A.; Fonseca, G.; Fuentes, J.L.; Cozzi, R.; Cundari, E.; Fiore, M.; Ricordy, R.; Perticone, P.; Degrassi, F.; De Salvia, R. As-sessment of the genotoxic risk of Punica granatum L. (Punicaceae) whole fruit extracts. J. Ethnopharmacol. 2008, 115, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Labieniec, M.; Gabryelak, T. Effects of tannins on Chinese hamster cell line B14. Mutat. Res. Toxicol. Env. Mutagen. 2003, 539, 127–135. [Google Scholar] [CrossRef]


{kind=link}
| In Vitro | Preclinical | Clinical/Interventional | Aim/Mechanism | References |
|---|---|---|---|---|
| KB and CAL27 oral cancer, SW480, SW620, HT29 and HCT116 colon cancer and RWPE-1 prostate cancer cell lines | - | - | Apoptotic and antioxidant activity | [8] |
| Vero (normal African green monkey kidney cell line), Hep-2 (human larynx epithelial cancer cell line), and A-549 (human small cell lung carcinoma cell line) | - | - | ROS elimination and antioxidant activity | [11] |
| HeLa cells | - | - | Antiproliferative activity (β-catenin signaling pathway) | [12] |
| Vero (African green monkey kidney cells, ATCC CCL-81), HEL (human embryonic lung fibroblast, ATCC CCL-137), and A549 (human lung carcinoma, ATCC CCL-185) | - | - | Antiviral activity | [14] |
| - | - | 20 young healthy volunteers (10 men (BMI 21.8 ± 2.5 kg/m2) and 10 women (BMI 23.4 ± 1.6 kg/m), with an average age of 20.5 ± 2.0 and 21.5 ± 1.5 years | Bioavailability of PUN and EA | [20] |
| - | - | Healthy volunteers (n = 49, 32 men and 17 women; BMI > 27 kg/m2) aged between 40 and 65 years | Metabolism EA | [23] |
| - | Rat | - | Bioavailability and metabolism of PUN and EA | [26] |
| - | - | Healthy volunteers (n = 6, 4 men and 2 women) | Bioavailability and metabolism of PUN and EA | [27] |
| RAW264.7 cells | - | - | Anti-inflammatory activity | [32] |
| RAW264.7 cells | - | - | Anti-inflammatory activity | [33] |
| Splenocytes from Balb/c mice and normal splenocytes from C57 Bl/6 mice | Mouse Balb/c | - | Immunosuppressive activity | [34] |
| RAW264.7 cells | - | - | Anti-inflammatory activity | [35] |
| PBMCs from healthy volunteers | - | - | Anti-inflammatory activity | [36] |
| Jurkat E2 cells and PBMC | - | - | Immunosuppressive activity | [37] |
| Buňky (ME-180)—Cell carcinoma of the cervix uteri | - | - | Apoptotic activity through the mitochondrial and NF-kB pathway | [40] |
| - | Mouse (Swiss Webster) | - | Chemoprotective and angiogenic activity | [41] |
| human osteosarcoma cell lines (U2OS, MG63 and SaOS2) and normal osteoblast cell line (hFOB1.19) | - | - | Antiproliferative activity | [43] |
| human colon cell line Caco-2 and the normal colon cells CCD-112CoN | - | - | Apoptotic activity | [45] |
| human U87MG glioma cells | - | - | Apoptotic activity | [46] |
| Streptococcus mutans | - | - | Antibacterial activity | [49] |
| SARS-CoV-2 (spike protein) | - | - | Antiviral activity | [50] |
| - | Mouse Balb/c | - | Antioxidant activity | [54] |
| - | Male Rats (Sprague-Dawley) | - | Antihepatotoxicity | [57] |
| - | Male Wistar rats | - | Antihepatotoxicity and antineurotoxicity | [59] |
| 3T3-L1 murine pre-adipocytes | - | - | Antiobesity activity | [60] |
| 3T3-L1 mouse adipocytes | - | - | Antiobesity activity | [61] |
| Human adipocytes (SGBS), human myocytes (HSMM) | - | - | Antiobesity activity | [62] |
| Human Umbilical Vein Endothelial Cells (HUVECs) | - | - | Antiatherogenic effects | [63] |
| THP-1 cells (Human immortalized cells, ATCC, TIB-202) | Male apoE−/− mice | - | Anti-atherosclerotic activity | [65] |
| - | Wistar rats (n = 48) | - | Anti-atherosclerotic activity | [66] |
| - | Sprague−Dawley rats | - | Toxicity effect | [67] |
| Chinese hamster ovary (CHO) cells | Mouse Balb/c | - | Genotoxicity effect | [68] |
| Chinese hamster cells (B14 cell line) | Genotoxicity and cytotoxicity effect | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venusova, E.; Kolesarova, A.; Horky, P.; Slama, P. Physiological and Immune Functions of Punicalagin. Nutrients 2021, 13, 2150. https://doi.org/10.3390/nu13072150
Venusova E, Kolesarova A, Horky P, Slama P. Physiological and Immune Functions of Punicalagin. Nutrients. 2021; 13(7):2150. https://doi.org/10.3390/nu13072150
Chicago/Turabian StyleVenusova, Eva, Adriana Kolesarova, Pavel Horky, and Petr Slama. 2021. "Physiological and Immune Functions of Punicalagin" Nutrients 13, no. 7: 2150. https://doi.org/10.3390/nu13072150
APA StyleVenusova, E., Kolesarova, A., Horky, P., & Slama, P. (2021). Physiological and Immune Functions of Punicalagin. Nutrients, 13(7), 2150. https://doi.org/10.3390/nu13072150