
Milk Osteopontin for Gut, Immunity and Brain Development in Preterm Pigs

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Antibacterial Assay
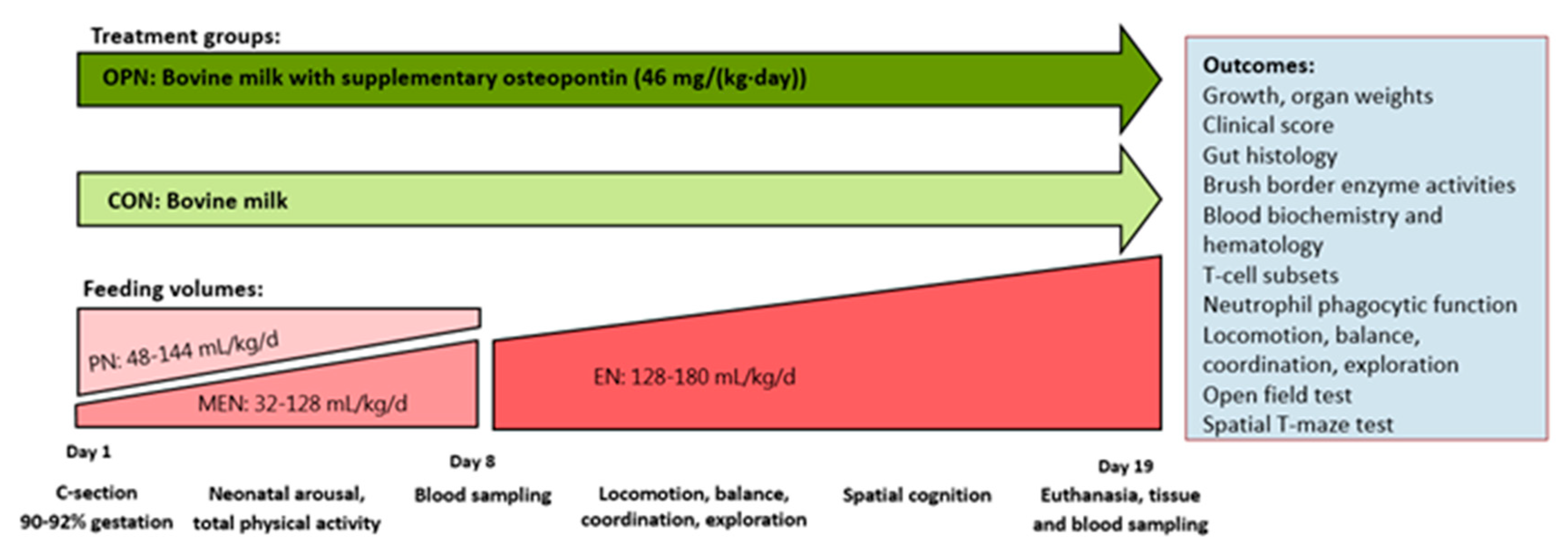
2.2. Animals and Experimental Design
2.3. Nutrition and Feeding
2.4. Clinical Evaluation and Tissue Collection
2.5. Intestinal Mucosa Structure and Brush Border Enzyme Activities
2.6. Blood Biochemistry, Hematology and Systemic Immunity
2.7. Early Motor Development, Behavior and Cognition
2.8. Statistical Analysis
3. Results
3.1. In Vitro Antibacterial Assay
3.2. Clinical and Growth Outcomes
3.3. Gut Structure and Function
3.4. Blood Biochemistry, Hematology and Systemic Immunity
3.5. Brain Structural and Functional Outcomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lönnerdal, B. Infant formula and infant nutrition: Bioactive proteins of human milk and implications for composition of infant formulas. Am. J. Clin. Nutr. 2014, 99, 712–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ip, S.; Chung, M.; Raman, G.; Chew, P.; Magula, N.; DeVine, D.; Trikalinos, T.; Lau, J. Breastfeeding and maternal and infant health outcomes in developed countries. Evid. Rep. Technol. Assess. 2007, 153, 1–186. [Google Scholar]
- Cortez, J.; Makker, K.; Kraemer, D.F.; Neu, J.; Sharma, R.; Hudak, M.L. Maternal milk feedings reduce sepsis, necrotizing enterocolitis and improve outcomes of premature infants. J. Perinatol. 2018, 38, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Schack, L.; Lange, A.; Kelsen, J.; Agnholt, J.; Christensen, B.; Petersen, T.E.; Sørensen, E.S. Considerable variation in the concentration of osteopontin in human milk, bovine milk, and infant formulas. J. Dairy Sci. 2009, 92, 5378–5385. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Lönnerdal, B. Osteopontin in human milk and infant formula affects infant plasma osteopontin concentrations. Pediatr. Res. 2019, 85, 502–505. [Google Scholar] [CrossRef]
- Christensen, B.; Sørensen, E.S. Structure, function and nutritional potential of milk osteopontin. Int. Dairy J. 2016, 57, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bruun, S.; Jacobsen, L.N.; Ze, X.; Husby, S.; Ueno, H.M.; Nojiri, K.; Sørensen, E.S. Osteopontin levels in human milk vary across countries and within lactation period: Data from a multicenter study. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Denhardt, D.T.; Guo, X. Osteopontin: A protein with diverse functions. FASEB J. 1993, 7, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.; Karlsen, N.J.; Jørgensen, S.D.S.; Jacobsen, L.N.; Ostenfeld, M.S.; Petersen, S.V.; Müllertz, A.; Sørensen, E.S. Milk osteopontin retains integrin-binding activity after in vitro gastrointestinal transit. J. Dairy Sci. 2020, 103, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Jiang, R.; Lönnerdal, B. Assessment of bioactivities of the human milk lactoferrin–osteopontin complex in vitro. J. Nutr. Biochem. 2019, 69, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jiang, R.; Liu, J.; Lönnerdal, B. The bovine Lactoferrin-Osteopontin complex increases proliferation of human intestinal epithelial cells by activating the PI3K/Akt signaling pathway. Food Chem. 2019, 310, 125919. [Google Scholar] [CrossRef]
- Donovan, S.M.; Monaco, M.H.; Drnevich, J.; Kvistgaard, A.S.; Hernell, O.; Lönnerdal, B. Bovine Osteopontin Modifies the Intestinal Transcriptome of Formula-Fed Infant Rhesus Monkeys to Be More Similar to Those That Were Breastfed. J. Nutr. 2014, 144, 1910–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, A.P.B.; Ellen, R.P.; Sørensen, E.S.; Goldberg, H.A.; Zohar, R.; Sodek, J. Osteopontin attenuation of dextran sulfate sodium-induced colitis in mice. Lab. Investig. 2009, 89, 1169–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.; Prell, C.; Lönnerdal, B. Milk osteopontin promotes brain development by up-regulating osteopontin in the brain in early life. FASEB J. 2019, 33, 1681–1694. [Google Scholar] [CrossRef] [PubMed]
- Giachelli, C.M.; Steitz, S. Osteopontin: A versatile regulator of inflammation and biomineralization. Matrix Biol. 2000, 19, 615–622. [Google Scholar] [CrossRef]
- Schack, L.; Stapulionis, R.; Christensen, B.; Kofod-Olsen, E.; Sørensen, U.B.S.; Vorup-Jensen, T.; Sørensen, E.S.; Höllsberg, P. Osteopontin Enhances Phagocytosis through a Novel Osteopontin Receptor, the αXβ2 Integrin. J. Immunol. 2009, 182, 6943–6950. [Google Scholar] [CrossRef] [Green Version]
- Holt, P.G.; Jones, C.A. The development of the immune system during pregnancy and early life. Allergy Eur. J. Allergy Clin. Immunol. 2000, 55, 688–697. [Google Scholar] [CrossRef]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (osteopontin): An early component of type-1 (cell-mediated) immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef]
- Anderson, J.W.; Johnstone, B.M.; Remley, D.T. Breast-feeding and cognitive development: A meta-analysis. Am. J. Clin. Nutr. 1999, 70, 525–535. [Google Scholar] [CrossRef]
- Horta, B.L.; Loret De Mola, C.; Victora, C.G. Breastfeeding and intelligence: A systematic review and meta-analysis. Acta Paediatr. Int. J. Paediatr. 2015, 104, 14–19. [Google Scholar] [CrossRef]
- Kramer, M.S.; Aboud, F.; Mironova, E.; Vanilovich, I.; Platt, R.W. Breastfeeding and child cognitive development. Arch Gen Psychiatry 2008, 65, 578–584. [Google Scholar] [CrossRef]
- Jiang, R.; Lönnerdal, B. Evaluation of Bioactivities of Bovine Milk Osteopontin Using a Knockout Mouse Model. J. Pediatr. Gastroenterol. Nutr. 2020, 71, 125–131. [Google Scholar] [CrossRef]
- Selvaraju, R.; Bernasconi, L.; Losberger, C.; Graber, P.; Kadi, L.; Avellana-Adalid, V.; Picard-Riera, N.; Van Evercooren, A.B.; Cirillo, R.; Kosco-Vilbois, M.; et al. Osteopontin is upregulated during in vivo demyelination and remyelination and enhances myelin formation in vitro. Mol. Cell. Neurosci. 2004, 25, 707–721. [Google Scholar] [CrossRef]
- Joung, S.; Fil, J.E.; Heckmann, A.B.; Kvistgaard, A.S.; Dilger, R.N. Early-Life Supplementation of Bovine Milk Osteopontin Supports Neurodevelopment and Influences Exploratory Behavior. Nutrients 2020, 12, 2206. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.H.; Thomas, P.; Peabody, J. Extrauterine growth restriction remains a serious problem in prematurely born neonates. Pediatrics 2003, 111, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.J.; Hansen, N. Infections in VLBW infants: Studies from the NICHD Neonatal Research Network. Semin. Perinatol. 2003, 27, 293–301. [Google Scholar] [CrossRef]
- Lin, P.W.; Nasr, T.R.; Stoll, B.J. Necrotizing Enterocolitis: Recent Scientific Advances in Pathophysiology and Prevention. Semin. Perinatol. 2008, 32, 70–82. [Google Scholar] [CrossRef]
- Saigal, S.; Doyle, L.W. An overview of mortality and sequelae of preterm birth from infancy to adulthood. Lancet 2008, 371, 261–269. [Google Scholar] [CrossRef]
- Møller, H.K.; Thymann, T.; Fink, L.N.; Frokiaer, H.; Kvistgaard, A.S.; Sangild, P.T. Bovine colostrum is superior to enriched formulas in stimulating intestinal function and necrotising enterocolitis resistance in preterm pigs. Br. J. Nutr. 2011, 105, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Obelitz-Ryom, K.; Bering, S.B.; Overgaard, S.H.; Eskildsen, S.F.; Ringgaard, S.; Olesen, J.L.; Skovgaard, K.; Pankratova, S.; Wang, B.; Brunse, A.; et al. Bovine milk oligosaccharides with sialyllactose improves cognition in preterm pigs. Nutrients 2019, 11, 1335. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.D.; Nguyen, D.N.; Langhorn, L.; Renes, I.B.; van Elburg, R.M.; Hartog, A.; Tims, S.; van de Looij, Y.; Sangild, P.T.; Thymann, T. Synbiotics Combined with Glutamine Stimulate Brain Development and the Immune System in Preterm Pigs. J. Nutr. 2018, 148, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sangild, P.T.; Thymann, T.; Schmidt, M.; Stoll, B.; Burrin, D.G.; Buddington, R.K. Invited Review: The preterm pig as a model in pediatric gastroenterology. J. Anim. Sci. 2013, 91, 4713–4729. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.D.; Sangild, P.T.; Munch, S.L.; van der Beek, E.M.; Renes, I.B.; van Ginneken, C.; Greisen, G.O.; Thymann, T. Delayed growth, motor function and learning in preterm pigs during early postnatal life. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2016, 310, R481–R492. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.N.; Jiang, P.; Frøkiær, H.; Heegaard, P.M.H.; Thymann, T.; Sangild, P.T. Delayed development of systemic immunity in preterm pigs as a model for preterm infants. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.N.; Currie, A.J.; Ren, S.; Bering, S.B.; Sangild, P.T. Heat treatment and irradiation reduce anti-bacterial and immune-modulatory properties of bovine colostrum. J. Funct. Foods 2019, 57, 182–189. [Google Scholar] [CrossRef]
- Trend, S.; Strunk, T.; Hibbert, J.; Kok, C.H.; Zhang, G.; Doherty, D.A.; Richmond, P.; Burgner, D.; Simmer, K.; Davidson, D.J.; et al. Antimicrobial protein and peptide concentrations and activity in human breast milk consumed by preterm infants at risk of late-onset neonatal sepsis. PLoS ONE 2015, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ogundele, M.O. Effects of storage on the physicochemical and antibacterial properties of human milk. Br. J. Biomed. Sci. 2002, 59, 205–211. [Google Scholar] [CrossRef]
- Galac, M.R.; Stam, J.; Maybank, R.; Hinkle, M.; Mack, D.; Rohde, H.; Roth, A.L.; Fey, P.D. Complete genome sequence of Staphylococcus epidermidis 1457. Genome Announc. 2017, 5, 15–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Nguyen, D.N.; de Waard, M.; Christensen, L.; Zhou, P.; Jiang, P.; Sun, J.; Bojesen, A.M.; Lauridsen, C.; Lykkesfeldt, J.; et al. Pasteurization Procedures for Donor Human Milk Affect Body Growth, Intestinal Structure, and Resistance against Bacterial Infections in Preterm Pigs. J. Nutr. 2017, 147, 1121–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Jensen, M.L.; Chatterton, D.E.W.; Jensen, B.B.; Thymann, T.; Kvistgaard, A.S.; Sangild, P.T. Raw bovine milk improves gut responses to feeding relative to infant formula in preterm piglets. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G81–G90. [Google Scholar] [CrossRef] [PubMed]
- Vegge, A.; Thymann, T.; Lauritzen, L.; Bering, S.B.; Wiinberg, B.; Sangild, P.T. Parenteral lipids and partial enteral nutrition affect hepatic lipid composition but have limited short term effects on formula-induced necrotizing enterocolitis in preterm piglets. Clin. Nutr. 2014. [Google Scholar] [CrossRef]
- Shen, R.L.; Thymann, T.; Østergaard, M.V.; Støy, A.C.F.; Krych, L.; Nielsen, D.S.; Lauridsen, C.; Hartmann, B.; Holst, J.J.; Burrin, D.G.; et al. Early gradual feeding with bovine colostrum improves gut function and NEC resistance relative to infant formula in preterm pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G310–G323. [Google Scholar] [CrossRef]
- Ren, S.; Hui, Y.; Goericke-Pesch, S.; Pankratova, S.; Kot, W.; Pan, X.; Thymann, T.; Sangild, P.T.; Nguyen, D.N. Gut and immune effects of bioactive milk factors in preterm pigs exposed to prenatal inflammation. Am. J. Physiol.-Gastrointest. Liver Physiol. 2019, 317, G67–G77. [Google Scholar] [CrossRef]
- Elmore, M.R.P.; Dilger, R.N.; Johnson, R.W. Place and direction learning in a spatial T-maze task by neonatal piglets. Anim. Cogn. 2012, 15, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Jiang, R.; Lönnerdal, B. Biological roles of milk osteopontin. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 214–219. [Google Scholar] [CrossRef]
- Soeorg, H.; Huik, K.; Parm, U.; Ilmoja, M.-L.; Metelskaja, N.; Metsvaht, T.; Lutsaret, I. Genetic relatedness of coagulase-negative staphylococci from gastrointestinal tract and blood of preterm neonates with late-onset sepsis. Pediatr. Infect. Dis. J. 2013, 32, 389–393. [Google Scholar] [CrossRef] [Green Version]
- Golińska, E.; Strus, M.; Tomusiak-Plebanek, A.; Więcek, G.; Kozień, L.; Lauterbach, R.; Pawlik, D.; Rzepecka-Węglarz, B.; Kędzierska, J.; Dorycka, M.; et al. Coagulase-Negative Staphylococci Contained in Gut Microbiota as a Primary Source of Sepsis in Low- and Very Low Birth Weight Neonates. J. Clin. Med. 2020, 9, 2517. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.; Zachariae, E.D.; Poulsen, N.A.; Buitenhuis, A.; Larsen, L.; Sørensen, E. Factors influencing milk osteopontin concentration based on measurements from Danish Holstein cows. J. Dairy Res. 2021, 88, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Obelitz-Ryom, K.; Rendboe, A.K.; Nguyen, D.N.; Rudloff, S.; Brandt, A.B.; Nielsen, D.S.; Heckmann, A.B.; Chichlowski, M.; Sangild, P.T.; Thymann, T.; et al. Bovine milk oligosaccharides with sialyllactose for preterm piglets. Nutrients 2018, 10, 1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, C.H.; Hui, Y.; Nguyen, D.N.; Ahnfeldt, A.M.; Burrin, D.G.; Hartmann, B.; Heckmann, A.B.; Sangild, P.T.; Thymann, T.; Bering, S.B. Alpha-lactalbumin enriched whey protein concentrate to improve gut, immunity and brain development in preterm pigs. Nutrients 2020, 12, 245. [Google Scholar] [CrossRef] [Green Version]
- Ahnfeldt, A.M.; Bæk, O.; Hui, Y.; Nielsen, C.H.; Obelitz-Ryom, K.; Busk-Anderson, T.; Ruge, A.; Holst, J.J.; Rudloff, S.; Burrin, D.; et al. Nutrient Restriction has Limited Short-Term Effects on Gut, Immunity, and Brain Development in Preterm Pigs. J. Nutr. 2020, 150, 1196–1207. [Google Scholar] [CrossRef] [PubMed]
- Butler, W.T. Structural and Functional Domains of Osteopontin. Ann. N. Y. Acad. Sci. 1995, 760, 6–11. [Google Scholar] [CrossRef]
- Burkey, T.E.; Skjolaas, K.A.; Dritz, S.S.; Minton, J.E. Expression of Toll-like receptors, interleukin 8, macrophage migration inhibitory factor, and osteopontin in tissues from pigs challenged with Salmonella enterica serovar Typhimurium or serovar Choleraesuis. Vet. Immunol. Immunopathol. 2007, 115, 309–319. [Google Scholar] [CrossRef]
- Li, Y.; Østergaard, M.V.; Jiang, P.; Chatterton, D.E.W.; Thymann, T.; Kvistgaard, A.S.; Sangild, P.T. Whey Protein Processing Influences Formula-Induced Gut Maturation in Preterm Pigs. J. Nutr. 2013, 143, 1934–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, C.E.; Kvistgaard, A.S.; Peerson, J.M.; Donovan, S.M.; Peng, Y.M.; Lönnerdal, B. Effects of osteopontin-enriched formula on lymphocyte subsets in the first 6 months of life: A randomized controlled trial. Pediatr. Res. 2017, 82, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Fleming, S.A.; Dilger, R.N. Young pigs exhibit differential exploratory behavior during novelty preference tasks in response to age, sex, and delay. Behav. Brain Res. 2017, 321, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Manaenko, A.; Fan, R.; Huang, L.; Enkhjargal, B.; McBride, D.W.; Ding, Y.; Tang, J.; Xiao, X.; Zhang, J.H. Osteopontin attenuates inflammation via JAK2/STAT1 pathway in hyperglycemic rats after intracerebral hemorrhage. Neuropharmacology 2018, 138, 160–169. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | OPN | CON | p-Value |
|---|---|---|---|---|
| Birth weight | g | 1051 ± 46.6 | 1035 ± 49.0 | NS |
| Weight at euthanasia, Day 19 | g | 1789 ± 115 | 1861 ± 101 | NS |
| Relative organ dimensions 1 | ||||
| Small intestine length | cm/kg | 279 ± 10.4 | 287 ± 8.19 | NS |
| Prox weight | g/kg | 14.2 ± 0.28 | 14.1 ± 0.39 | NS |
| Mid weight | g/kg | 12.4 ± 0.33 | 12.3 ± 0.36 | NS |
| Dist weight | g/kg | 12.3 ± 0.29 | 12.1 ± 0.34 | NS |
| Stomach weight | g/kg | 6.24 ± 0.18 | 5.94 ± 0.15 | <0.01 |
| Colon weight | g/kg | 32.1 ± 2.38 | 35.7 ± 3.63 | NS |
| Liver weight | g/kg | 25.9 ± 0.96 | 26.3 ± 0.49 | NS |
| Spleen weight | g/kg | 3.22 ± 0.18 | 3.50 ± 0.13 | NS |
| Heart weight | g/kg | 7.76 ± 0.30 | 8.14 ± 0.23 | NS |
| Lungs weight | g/kg | 18.6 ± 1.03 | 21.3 ± 0.96 | NS |
| Kidney weight | g/kg | 7.33 ± 0.20 | 7.09 ± 0.19 | NS |
| Parameter | Unit | OPN | CON | p-Value |
|---|---|---|---|---|
| Prox, villus height | µm | 604 ± 20.4 | 576 ± 18.1 | NS |
| Prox, crypt depth | µm | 92.7 ± 2.74 | 96.8 ± 1.89 | <0.01 |
| Mid, villus height | µm | 590 ± 22.7 | 546 ± 17.6 | NS |
| Mid, crypt depth | µm | 92.2 ± 2.50 | 93.9 ± 2.16 | NS |
| Dist, villus height | µm | 490 ± 19.2 | 457 ± 14.5 | NS |
| Dist, crypt depth | µm | 88.5 ± 2.84 | 88.4 ± 1.82 | NS |
| Gastric content 1 | g/kg | 17.3 ± 1.90 | 17.1 ± 0.96 | NS |
| Brush border enzyme activities (Mid) | ||||
| Lactase | U/g tissue | 34.7 ± 3.15 | 43.1 ± 3.98 | 0.08 |
| Maltase | U/g tissue | 7.80 ± 1.83 | 7.31 ± 1.06 | NS |
| Sucrase | U/g tissue | 1.80 ± 0.29 | 1.79 ± 0.18 | NS |
| ApA | U/g tissue | 7.00 ± 0.50 | 6.35 ± 0.44 | NS |
| ApN | U/g tissue | 11.2 ± 0.44 | 10.2 ± 0.60 | NS |
| DPPIV | U/g tissue | 3.65 ± 0.27 | 3.40 ± 0.20 | NS |
| Parameter | Unit | Day | OPN | CON | p-Value |
|---|---|---|---|---|---|
| Hematology | |||||
| Total erythrocytes | 1012/L | 8 19 | 3.25 ± 0.18 3.56 ± 0.15 | 3.20 ± 0.09 3.52 ± 0.11 | NS NS |
| Hemoglobin | mmol/L | 8 19 | 4.32 ± 0.23 4.12 ± 0.15 | 4.22 ± 0.11 3.98 ± 0.12 | NS NS |
| Hematocrit | % | 8 19 | 22.9 ± 0.01 21.5 ± 0.01 | 22.3 ± 0.01 21.0 ± 0.01 | NS NS |
| Thrombocytes | 109/L | 8 19 | 369 ± 40.9 432 ± 79.2 | 351 ± 27.3 545 ± 26.0 | NS NS |
| Immune cell counts | |||||
| Total leukocytes | 109/L | 8 19 | 5.38 ± 0.59 14.2 ± 2.12 | 4.69 ± 0.31 14.1 ± 1.76 | NS NS |
| Lymphocytes | 109/L | 8 19 | 1.91 ± 0.16 2.52 ± 0.16 | 1.51 ± 0.10 2.39 ± 0.13 | 0.01 * NS |
| Monocytes | 109/L | 8 19 | 0.18 ± 0.04 0.53 ± 0.14 | 0.08 ± 0.01 0.48 ± 0.05 | 0.02 * NS |
| Neutrophils | 109/L | 8 19 | 3.13 ± 0.57 10.9 ± 1.96 | 3.01 ± 0.31 11.0 ± 1.71 | NS NS |
| Eosinophils | 109/L | 8 19 | 0.08 ± 0.04 0.04 ± 0.01 | 0.03 ± 0.01 0.04 ± 0.01 | NS NS |
| Basophils | 109/L | 8 19 | 0.01 ± 0.00 0.01 ± 0.00 | 0.01 ± 0.00 0.01 ± 0.01 | NS NS |
| Large unstained cells | 109/L | 8 19 | 0.06 ± 0.01 0.25 ± 0.08 | 0.06 ± 0.01 0.17 ± 0.03 | NS NS |
| T-cell subsets | |||||
| T-cell frequency (CD3+ lymphocytes) | % | 8 19 | 59.5 ± 2.34 65.4 ± 2.09 | 59.9 ± 1.96 65.2 ± 1.62 | NS NS |
| Th-cell frequency (CD3+CD4+CD8-) | % | 8 19 | 55.0 ± 0.02 49.4 ± 3.08 | 58.3 ± 0.01 53.2 ± 1.97 | 0.09 NS |
| Tc-cells frequency (CD3+CD4-CD8+) | % | 8 19 | 8.38 ± 0.01 10.1 ± 0.94 | 8.71 ± 0.01 11.7 ± 0.60 | NS NS |
| Treg-cells frequency (CD3+CD4+Foxp3+) | % | 8 19 | 6.82 ± 0.00 4.86 ± 0.30 | 7.66 ± 0.00 4.94 ± 0.40 | NS NS |
| Neutrophil phagocytic function | |||||
| Neutrophil phagocytic rate | % | 8 | 92.4 ± 1.51 | 91.7 ± 1.58 | NS |
| Neutrophil phagocytic capacity, MFI | 8 | 15,488 ± 1304 | 15,440 ± 802 | NS |
| Parameter | Unit | OPN | CON | p-Value |
|---|---|---|---|---|
| Absolute values | ||||
| Total brain weight | g | 37.0 ± 0.58 | 37.3 ± 0.49 | NS |
| Cerebrum weight | g | 29.2 ± 0.47 | 29.6 ± 0.48 | NS |
| Cerebellum weight | g | 3.81 ± 0.07 | 3.88 ± 0.05 | NS |
| Brain stem weight | g | 3.54 ± 0.06 | 3.62 ± 0.04 | NS |
| Left hippocampus weight | g | 0.65 ± 0.03 | 0.67 ± 0.02 | NS |
| Left caudate nucleus weight | g | 0.33 ± 0.01 | 0.35 ± 0.01 | NS |
| Cerebral water fraction | % | 82.7 ± 0.00 | 82.9 ± 0.00 | NS |
| Relative values | ||||
| Total brain weight | g/kg | 18.7 ± 0.91 | 20.0 ± 1.05 | NS |
| Cerebrum weight | % | 79.3 ± 0.17 | 79.4 ± 0.51 | NS |
| Cerebellum weight | % | 10.4 ± 0.13 | 10.4 ± 0.13 | NS |
| Brain stem weight | % | 9.65 ± 0.12 | 9.71 ± 0.09 | NS |
| Left hippocampus weight | % | 1.76 ± 0.07 | 1.78 ± 0.04 | NS |
| Left caudate nucleus weight | % | 0.91 ± 0.03 | 0.95 ± 0.03 | NS |
| Open field | ||||
| Overall score | point | 5.75 ± 0.23 | 5.79 ± 0.21 | NS |
| Total distance moved | m | 17.1 ± 2.06 | 17.6 ± 1.34 | NS |
| Velocity | cm/sec | 11.4 ± 1.38 | 11.7 ± 0.90 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aasmul-Olsen, K.; Henriksen, N.L.; Nguyen, D.N.; Heckmann, A.B.; Thymann, T.; Sangild, P.T.; Bering, S.B. Milk Osteopontin for Gut, Immunity and Brain Development in Preterm Pigs. Nutrients 2021, 13, 2675. https://doi.org/10.3390/nu13082675
Aasmul-Olsen K, Henriksen NL, Nguyen DN, Heckmann AB, Thymann T, Sangild PT, Bering SB. Milk Osteopontin for Gut, Immunity and Brain Development in Preterm Pigs. Nutrients. 2021; 13(8):2675. https://doi.org/10.3390/nu13082675
Chicago/Turabian StyleAasmul-Olsen, Karoline, Nicole Lind Henriksen, Duc Ninh Nguyen, Anne Birgitte Heckmann, Thomas Thymann, Per Torp Sangild, and Stine Brandt Bering. 2021. "Milk Osteopontin for Gut, Immunity and Brain Development in Preterm Pigs" Nutrients 13, no. 8: 2675. https://doi.org/10.3390/nu13082675