
Human Milk Oligosaccharide 3′-GL Improves Influenza-Specific Vaccination Responsiveness and Immunity after Deoxynivalenol Exposure in Preclinical Models

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vivo Experiments
2.1.1. Study Design
2.1.2. Antigen-Specific Delayed-Type Hypersensitivity Reactions
2.1.3. Vaccine-Specific Immunoglobulins (Igs) in Serum
2.1.4. Splenocyte Isolation and Flow Cytometry of Immune Cells
2.1.5. Re-Stimulation of Splenocytes with Vaccine-Loaded Bone Marrow-Derived Dendritic Cells (BMDCs)
2.1.6. Isolation of RNA from Intestinal Samples for qRT-PCR
2.1.7. Histomorphometric Analysis of Intestinal Specimens
2.1.8. Short-Chain Fatty Acid (SCFA) Concentrations in Cecum
2.2. In Vitro Experiments
2.2.1. Cell Culture
2.2.2. Caco-2 Cell Monolayer Integrity Measurement
2.2.3. Cytokine Production
3. Statistical Analysis
4. Results
4.1. In Vivo Results
4.1.1. Dietary TOS Improved Vaccine-Specific Cellular and Humoral Responses in DON-Exposed Mice
4.1.2. Dietary TOS Increased Th1 Cell Activation and Attenuated DON-Induced Modification in T Cell Populations in the Spleen
4.1.3. Dietary TOS Reversed the Effect of DON on the B Cell Population and Activation in the Spleen
4.1.4. Dietary TOS Increased the Production of Type-1 Cytokine IFN-γ and Regulatory Cytokine IL-10 from Re-Stimulated Splenocytes of DON-Exposed Mice
4.1.5. Dietary TOS Reversed the Effect of DON on mRNA Expression of Junctional Proteins and Chemokine CXCL1 in the Ileum
4.1.6. Dietary TOS Prevented the Adverse Effect of DON on Villus Height of Ileum Sections
4.1.7. Dietary TOS Had No Significant Effect on SCFA, whereas a Higher Concentration of DON Reduced SCFA Levels in the Cecum
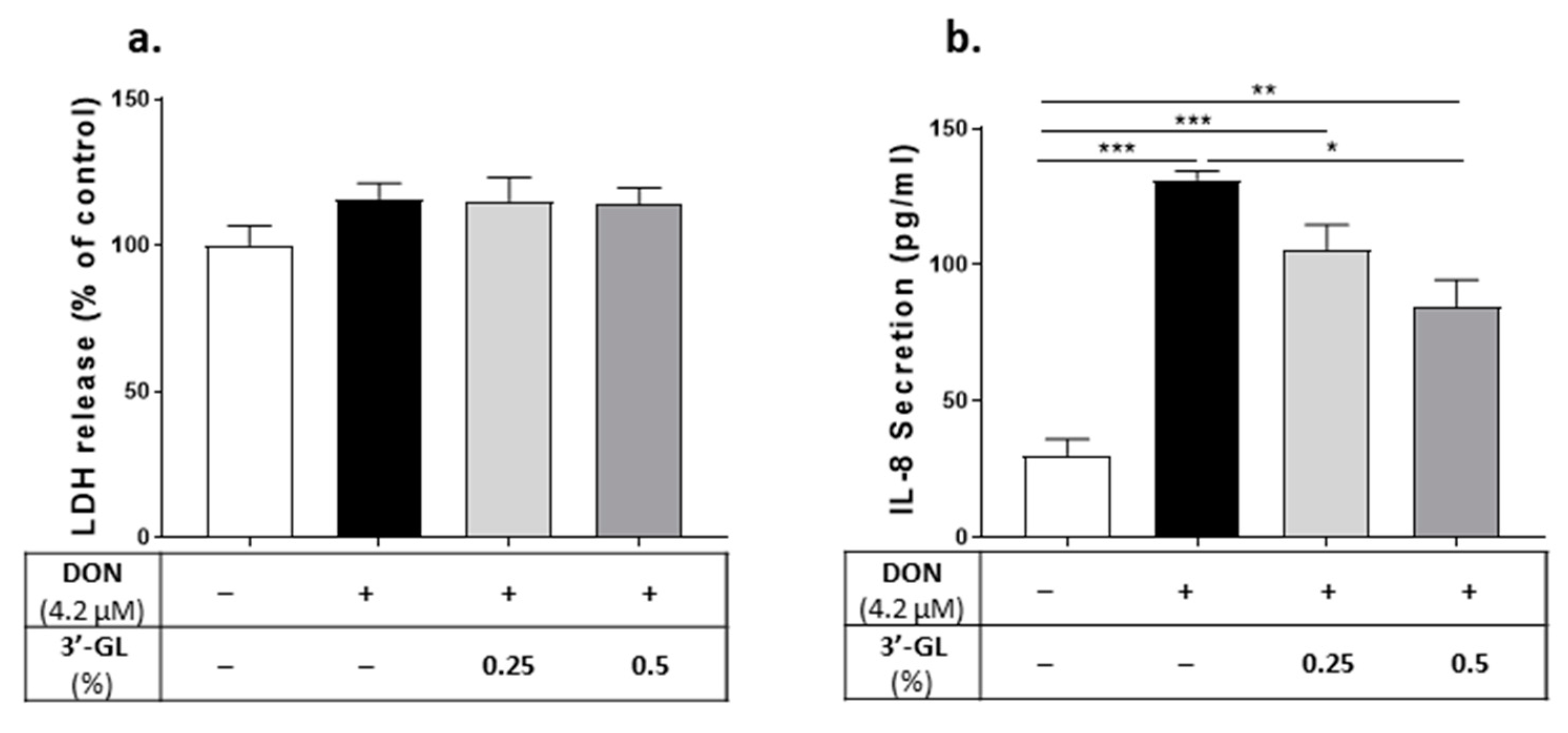
4.2. In Vitro Results: Protective Effect of TOS and Its Main Component, β3′-GL, on DON-Induced Impairment of Caco-2 Cell Monolayer Integrity and IL-8 Production
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murphy, P.A.; Hendrich, S.; Landgren, C.; Bryant, C.M. Food Mycotoxins: An Update. J. Food Sci. 2006, 71, R51–R65. [Google Scholar] [CrossRef]
- Pascari, X.; Marín, S.; Ramos, A.J.; Molino, F.; Sanchis, V. Deoxynivalenol in cereal-based baby food production process. A review. Food Control 2019, 99, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the Gut to the Brain: Journey and Pathophysiological Effects of the Food-Associated Trichothecene Mycotoxin Deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol Impairs Weight Gain and Affects Markers of Gut Health after Low-Dose, Short-Term Exposure of Growing Pigs. Toxins 2015, 7, 2071–2095. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J.; Zhou, H.-R.; Moon, Y.; Chung, Y. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unraveling a paradox. Toxicol. Lett. 2004, 153, 61–73. [Google Scholar] [CrossRef]
- Bimczok, D.; Döll, S.; Rau, H.; Goyarts, T.; Wundrack, N.; Naumann, M.; Dänicke, S.; Rothkötter, H.-J. The Fusarium toxin deoxynivalenol disrupts phenotype and function of monocyte-derived dendritic cells In Vivo and In Vitro. Immunobiology 2007, 212, 655–666. [Google Scholar] [CrossRef]
- Luongo, D.; Severino, L.; Bergamo, P.; D’Arienzo, R.; Rossi, M. Trichothecenes NIV and DON modulate the maturation of murine dendritic cells. Toxicon 2010, 55, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbana-Barnat, S.; Lafarge-Frayssinet, C.; Cohen, H.; Neish, G. Immunosuppressive properties of deoxynivalenol. Toxicology 1988, 48, 155–166. [Google Scholar] [CrossRef]
- Islam, M.R.; Roh, Y.S.; Kim, J.; Lim, C.W.; Kim, B. Differential immune modulation by deoxynivalenol (vomitoxin) in mice. Toxicol. Lett. 2013, 221, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M.; Madsen, K.; Spiller, R.; Meerveld, B.G.-V.; Verne, G.N. Intestinal barrier function in health and gastrointestinal disease. Neurogastroenterol. Motil. 2012, 24, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Akbari, P.; Braber, S.; Gremmels, H.; Koelink, P.J.; Verheijden, K.A.T.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol: A trigger for intestinal integrity breakdown. FASEB J. 2014, 28, 2414–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, P.; Braber, S.; Alizadeh, A.; Verheijden, K.; Schoterman, M.H.C.; Kraneveld, A.D.; Garssen, J.; Fink-Gremmels, J. Galacto-oligosaccharides Protect the Intestinal Barrier by Maintaining the Tight Junction Network and Modulating the Inflammatory Responses after a Challenge with the Mycotoxin Deoxynivalenol in Human Caco-2 Cell Monolayers and B6C3F1 Mice. J. Nutr. 2015, 145, 1604–1613. [Google Scholar] [CrossRef] [Green Version]
- Ramani, S.; Stewart, C.J.; Laucirica, D.R.; Ajami, N.J.; Robertson, B.; Autran, C.A.; Shinge, D.; Rani, S.; Anandan, S.; Hu, L.; et al. Human milk oligosaccharides, milk microbiome and infant gut microbiome modulate neonatal rotavirus infection. Nat. Commun. 2018, 9, 5010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martinez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Coppa, G.V.M.D.; Bruni, S.M.D.; Morelli, L.P.H.D.; Soldi, S.P.H.D.; Gabrielli, O.M.D. The First Prebiotics in Humans: Human Milk Oligosaccharides. Journal of clinical gastroenterology. J. Clin. Gastroenterol. 2004, 38, S80–S83. [Google Scholar] [CrossRef]
- Salminen, S.; Stahl, B.; Vinderola, G.; Szajewska, H. Infant Formula Supplemented with Biotics: Current Knowledge and Future Perspectives. Nutrients 2020, 12, 1952. [Google Scholar] [CrossRef]
- Vazquez, E.; Santos-Fandila, A.; Buck, R.; Rueda, R.; Ramirez, M. Major human milk oligosaccharides are absorbed into the systemic circulation after oral administration in rats. Br. J. Nutr. 2017, 117, 237–247. [Google Scholar] [CrossRef] [Green Version]
- DiFilippo, E.; Bettonvil, M.; Willems, R.; Braber, S.; Fink-Gremmels, J.; Jeurink, P.V.; Schoterman, M.H.C.; Gruppen, H.; Schols, H.A. Oligosaccharides in Urine, Blood, and Feces of Piglets Fed Milk Replacer Containing Galacto-oligosaccharides. J. Agric. Food Chem. 2015, 63, 10862–10872. [Google Scholar] [CrossRef]
- Zuurveld, M.; Van Witzenburg, N.P.; Garssen, J.; Folkerts, G.; Stahl, B.; Land, B.V.; Willemsen, L.E.M. Immunomodulation by Human Milk Oligosaccharides: The Potential Role in Prevention of Allergic Diseases. Front. Immunol. 2020, 11, 801. [Google Scholar] [CrossRef]
- Boehm, G.; Stahl, B.; Jelinek, J.; Knol, J.; Miniello, V.; Moro, G.E. Prebiotic carbohydrates in human milk and formulas. Acta Paediatr. 2007, 94, 18–21. [Google Scholar] [CrossRef]
- Fanaro, S.; Jelinek, J.; Stahl, B.; Boehm, G.; Kock, R.; Vigi, V. Acidic Oligosaccharides from Pectin Hydrolysate as New Component for Infant Formulae: Effect on Intestinal Flora, Stool Characteristics, and pH. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 186–190. [Google Scholar] [CrossRef]
- Boehm, G.; Fanaro, S.; Jelinek, J.; Stahl, B.; Marini, A. Prebiotic concept for infant nutrition. Acta Paediatr. 2003, 92, 64–67. [Google Scholar] [CrossRef]
- Van’t Land, B.; Schijf, M.; van Esch, B.C.A.M.; van Bergenhenegouwen, J.; Bastiaans, J.; Schouten, B.; Boon, L.; Garssen, J. Regulatory T-cells have a prominent role in the immune modulated vaccine response by specific oligosaccharides. Vaccine 2010, 28, 5711–5717. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.P.; M’Rabet, L.; Stahl, B.; Boehm, G.; Garssen, J. Immune-Modulatory Effects and Potential Working Mechanisms of Orally Applied Nondigestible Carbohydrates. Crit. Rev. Immunol. 2007, 27, 97–140. [Google Scholar] [CrossRef] [PubMed]
- Donald, A.S.; Feeney, J. Separation of human milk oligosaccharides by recycling chromatography. First isolation of lacto-N-neo-difucohexaose II and 3′-galactosyllactose from this source. Carbohydr. Res. 1988, 178, 79–91. [Google Scholar] [CrossRef]
- Newburg, D.S.; Ko, J.S.; Leone, S.; Nanthakumar, N.N. Human Milk Oligosaccharides and Synthetic Galactosyloligosaccharides Contain 3′-, 4-, and 6′-Galactosyllactose and Attenuate Inflammation in Human T84, NCM-460, and H4 Cells and Intestinal Tissue Ex Vivo. J. Nutr. 2016, 146, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Kobata, A. Oligosaccharides of human milk: V. Isolation and characterization of a new trisaccharide, 6′-galactosyllactose. Arch. Biochem. Biophys. 1974, 161, 164–170. [Google Scholar] [CrossRef]
- Panesar, P.; Kaur, R.; Singh, R.S.; Kennedy, J.F. Biocatalytic strategies in the production of galacto-oligosaccharides and its global status. Int. J. Biol. Macromol. 2018, 111, 667–679. [Google Scholar] [CrossRef]
- Van de Heijning, B.J.M.; Berton, A.; Bouritius, H.; Goulet, O. GI symptoms in infants are a potential target for fermented infant milk formulae: A review. Nutrients 2014, 6, 3942–3967. [Google Scholar] [CrossRef] [Green Version]
- Vos, A.P.; Knol, J.; Stahl, B.; M’Rabet, L.; Garssen, J. Specific prebiotic oligosaccharides modulate the early phase of a murine vaccination response. Int. Immunopharmacol. 2010, 10, 619–625. [Google Scholar] [CrossRef]
- Vos, A.P.; Haarman, M.; Buco, A.; Govers, M.; Knol, J.; Garssen, J.; Stahl, B.; Boehm, G.; M’Rabet, L. A specific prebiotic oligosaccharide mixture stimulates delayed-type hypersensitivity in a murine influenza vaccination model. Int. Immunopharmacol. 2006, 6, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Bol-Schoenmakers, M.; Braber, S.; Akbari, P.; De Graaff, P.; Van Roest, M.; Kruijssen, L.; Smit, J.; Esch, B.C.A.M.V.; Jeurink, P.V.; Garssen, J.; et al. The mycotoxin deoxynivalenol facilitates allergic sensitization to whey in mice. Mucosal Immunol. 2016, 9, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Van Loveren, H.; Van Amsterdam, J.G.; Vandebriel, R.J.; Kimman, T.G.; Rümke, H.C.; Steerenberg, P.S.; Vos, J.G. Vaccine-induced antibody responses as parameters of the influence of endogenous and environmental factors. Environ. Health Perspect. 2001, 109, 757–764. [Google Scholar] [CrossRef]
- Albers, R.; Antoine, J.-M.; Bourdet-Sicard, R.; Calder, P.C.; Gleeson, M.; LeSourd, B.; Samartín, S.; Sanderson, I.R.; Van Loo, J.; Dias, F.W.V.; et al. Markers to measure immunomodulation in human nutrition intervention studies. Br. J. Nutr. 2005, 94, 452–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef]
- Perrin, V.; Fenet, B.; Praly, J.-P.; Lecroix, F.; Ta, C.D. Identification and synthesis of a trisaccharide produced from lactose by transgalactosylation. Carbohydr. Res. 2000, 325, 202–210. [Google Scholar] [CrossRef]
- Xiao, L.; Leusink-Muis, T.; Kettelarij, N.; Van Ark, I.; Blijenberg, B.; Hesen, N.A.; Stahl, B.; Overbeek, S.A.; Garssen, J.; Folkerts, G.; et al. Human Milk Oligosaccharide 2′-Fucosyllactose Improves Innate and Adaptive Immunity in an Influenza-Specific Murine Vaccination Model. Front. Immunol. 2018, 9, 452. [Google Scholar] [CrossRef] [Green Version]
- Van Vlies, N.; Hogenkamp, A.; Fear, A.L.; Van Esch, B.C.; Oosting, A.; Van De Heijning, B.; Van Der Beek, E.; Calder, P.C.; Garssen, J. Perinatal programming of murine immune responses by polyunsaturated fatty acids. J. Dev. Orig. Health Dis. 2010, 2, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in Susceptibility to Heat Stress along the Chicken Intestine and the Protective Effects of Galacto-Oligosaccharides. PLoS ONE 2015, 10, e0138975. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.; Jacobson, K.A.; Rose, J.; Zeller, R. Hematoxylin and Eosin Staining of Tissue and Cell Sections. Cold Spring Harb. Protoc. 2008, 2008, pdb-prot4986. [Google Scholar] [CrossRef]
- De Theije, C.G.M.; Wopereisac, H.; Ramadan, M.; van Eijndthoven, T.; Lambert, J.; Knol, J.; Garssen, J.; Kraneveld, A.D.; Oozeer, R. Altered gut microbiota and activity in a murine model of autism spectrum disorders. Brain Behav. Immun. 2014, 37, 197–206. [Google Scholar] [CrossRef]
- Varasteh, S.; Braber, S.; Garssen, J.; Fink-Gremmels, J. Galacto-oligosaccharides exert a protective effect against heat stress in a Caco-2 cell model. J. Funct. Foods 2015, 16, 265–277. [Google Scholar] [CrossRef]
- Agematsu, K.; Hokibara, S.; Nagumo, H.; Komiyama, A. CD27: A memory B-cell marker. Immunol. Today 2000, 21, 204–206. [Google Scholar] [CrossRef]
- Hol, J.; Wilhelmsen, L.; Haraldsen, G. The murine IL-8 homologues KC, MIP-2, and LIX are found in endothelial cytoplasmic granules but not in Weibel-Palade bodies. J. Leukoc. Biol. 2009, 87, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.P. Immunotoxicity of Mycotoxins. J. Dairy Sci. 1993, 76, 892–897. [Google Scholar] [CrossRef]
- Xiao, L.; Engen, P.A.; Leusink-Muis, T.; van Ark, I.; Stahl, B.; Overbeek, S.A.; Garssen, J.; Naqib, A.; Green, S.J.; Keshavarzian, A.; et al. The Combination of 2′-Fucosyllactose with Short-Chain Galacto-Oligosaccharides and Long-Chain Fructo-Oligosaccharides that Enhance Influenza Vaccine Responses Is Associated with Mucosal Immune Regulation in Mice. J. Nutr. 2019, 149, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.; van Esch, B.; Stahl, B.; M’Rabet, L.; Folkerts, G.; Nijkamp, F.; Garssen, J. Dietary supplementation with specific oligosaccharide mixtures decreases parameters of allergic asthma in mice. Int. Immunopharmacol. 2007, 7, 1582–1587. [Google Scholar] [CrossRef]
- Courtade, L.C.; Han, S.; Lee, S.-H.; Mian, F.M.; Buck, R.; Forsythe, P. Attenuation of food allergy symptoms following treatment with human milk oligosaccharides in a mouse model. Allergy 2015, 70, 1091–1102. [Google Scholar] [CrossRef]
- Li, M.; Cuff, C.F.; Pestka, J. Modulation of Murine Host Response to Enteric Reovirus Infection by the Trichothecene Deoxynivalenol. Toxicol. Sci. 2005, 87, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Harkema, J.R.; Cuff, C.F.; Pestka, J.J. Deoxynivalenol Exacerbates Viral Bronchopneumonia Induced by Respiratory Reovirus Infection. Toxicol. Sci. 2006, 95, 412–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schijf, M.A.; Kerperien, J.; Bastiaans, J.; Szklany, K.; Meerding, J.; Hofman, G.; Boon, L.; Van Wijk, F.; Garssen, J.; Land, B.V. Alterations in Regulatory T Cells Induced by Specific Oligosaccharides Improve Vaccine Responsiveness in Mice. PLoS ONE 2013, 8, e75148. [Google Scholar] [CrossRef]
- Elsen, L.V.D.; Tims, S.; Jones, A.; Stewart, A.; Stahl, B.; Garssen, J.; Knol, J.; Forbes-Blom, E.; Land, B.V. Prebiotic oligosaccharides in early life alter gut microbiome development in male mice while supporting influenza vaccination responses. Benef. Microbes 2019, 10, 279–291. [Google Scholar] [CrossRef]
- Bridgman, S.L.; Azad, M.B.; Field, C.; Haqq, A.M.; Becker, A.B.; Mandhane, P.J.; Subbarao, P.; Turvey, S.; Sears, M.R.; Scott, J.A.; et al. Fecal Short-Chain Fatty Acid Variations by Breastfeeding Status in Infants at 4 Months: Differences in Relative versus Absolute Concentrations. Front. Nutr. 2017, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.H. Immune regulation by microbiome metabolites. Immunology 2018, 154, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Wrzosek, L.; Miquel, S.; Noordine, M.L.; Bouet, S.; Chevalier-Curt, M.J.; Robert, V.; Philippe, C.; Bridonneau, C.; Cherbuy, C.; Robbe-Masselot, C.; et al. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 2013, 11, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotoya, H.; Shigehisa, A.; Hara, T.; Matsumoto, H.; Hatano, H.; Matsuki, T. Identification of genes involved in galactooligosaccharide utilization in Bifidobacterium breve strain YIT 4014T. Microbiology 2017, 163, 1420–1428. [Google Scholar] [CrossRef]
- Toutounchi, N.S.; Hogenkamp, A.; Varasteh, S.; van’t Land, B.; Garssen, J.; Kraneveld, A.D.; Folkerts, G.; Braber, S. Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line. Toxins 2019, 11, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blikslager, A.T.; Moeser, A.; Gookin, J.; Jones, S.; Odle, J. Restoration of Barrier Function in Injured Intestinal Mucosa. Physiol. Rev. 2007, 87, 545–564. [Google Scholar] [CrossRef]
- Azagra-Boronat, I.; Massot-Cladera, M.; Mayneris-Perxachs, J.; Knipping, K.; Land, B.V.; Tims, S.; Stahl, B.; Garssen, J.; Àngels, F.; Castell, M.; et al. Immunomodulatory and Prebiotic Effects of 2′-Fucosyllactose in Suckling Rats. Front. Immunol. 2019, 10, 1773. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.; Bode, L.; Tappenden, K. Human Milk Oligosaccharides Influence Intestinal Epithelial Cell Maturation In Vitro. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 296–301. [Google Scholar] [CrossRef]
- Triantis, V.; Bode, L.; van Neerven, R. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef]
- Ayechu-Muruzabal, V.; Van Stigt, A.H.; Mank, M.; Willemsen, L.E.M.; Stahl, B.; Garssen, J.; Land, B.V. Diversity of Human Milk Oligosaccharides and Effects on Early Life Immune Development. Front. Pediatr. 2018, 6, 239. [Google Scholar] [CrossRef] [Green Version]
- De Filippo, K.; Dudeck, A.; Hasenberg, M.; Nye, E.; Van Rooijen, N.; Hartmann, K.; Gunzer, M.; Roers, A.; Hogg, N. Mast cell and macrophage chemokines CXCL1/CXCL2 control the early stage of neutrophil recruitment during tissue inflammation. Blood 2013, 121, 4930–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega-González, M.; Ocón, B.; Romero-Calvo, I.; Anzola, A.; Guadix, E.; Zarzuelo, A.; Guadix, E.; Zarzuelo, A.; Suárez, M.D.; de Medina, F.S.; et al. Nondigestible oligosaccharides exert nonprebiotic effects on intestinal epithelial cells enhancing the immune response via activation of TLR4-NFκB. Mol. Nutr. Food Res. 2014, 58, 384–393. [Google Scholar] [CrossRef]
- Press, B.; Di Grandi, D. Permeability for Intestinal Absorption: Caco-2 Assay and Related Issues. Curr. Drug Metab. 2008, 9, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.Y.; Chan, C.H.; Spicer, J.D.; Rousseau, M.C.; Giannias, B.; Rousseau, S.; Ferri, L.E. LPS-Induced TLR4 Signaling in Human Colorectal Cancer Cells Increases β1 Integrin-Mediated Cell Adhesion and Liver Metastasis. Cancer Res. 2011, 71, 1989–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Groups | Vaccination | n |
|---|---|---|
| Control (AIN93G) | - | 3 |
| Control (AIN93G) | + | 9 |
| 0.5% TOS (wt/wt) | + | 9 |
| 1% TOS (wt/wt) | + | 9 |
| DON (6.25 mg/kg of diet) | + | 9 |
| DON (12.5 mg/kg of diet) | + | 9 |
| DON (6.25 mg/kg) + 0.5% TOS | + | 9 |
| DON (12.5 mg/kg) + 0.5% TOS | + | 9 |
| DON (6.25 mg/kg) + 1% TOS | + | 9 |
| DON (12.5 mg/kg) + 1% TOS | + | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toutounchi, N.S.; Braber, S.; Hogenkamp, A.; Varasteh, S.; Cai, Y.; Wehkamp, T.; Tims, S.; Leusink-Muis, T.; van Ark, I.; Wiertsema, S.; et al. Human Milk Oligosaccharide 3′-GL Improves Influenza-Specific Vaccination Responsiveness and Immunity after Deoxynivalenol Exposure in Preclinical Models. Nutrients 2021, 13, 3190. https://doi.org/10.3390/nu13093190
Toutounchi NS, Braber S, Hogenkamp A, Varasteh S, Cai Y, Wehkamp T, Tims S, Leusink-Muis T, van Ark I, Wiertsema S, et al. Human Milk Oligosaccharide 3′-GL Improves Influenza-Specific Vaccination Responsiveness and Immunity after Deoxynivalenol Exposure in Preclinical Models. Nutrients. 2021; 13(9):3190. https://doi.org/10.3390/nu13093190
Chicago/Turabian StyleToutounchi, Negisa Seyed, Saskia Braber, Astrid Hogenkamp, Soheil Varasteh, Yang Cai, Tjalling Wehkamp, Sebastian Tims, Thea Leusink-Muis, Ingrid van Ark, Selma Wiertsema, and et al. 2021. "Human Milk Oligosaccharide 3′-GL Improves Influenza-Specific Vaccination Responsiveness and Immunity after Deoxynivalenol Exposure in Preclinical Models" Nutrients 13, no. 9: 3190. https://doi.org/10.3390/nu13093190
APA StyleToutounchi, N. S., Braber, S., Hogenkamp, A., Varasteh, S., Cai, Y., Wehkamp, T., Tims, S., Leusink-Muis, T., van Ark, I., Wiertsema, S., Stahl, B., Kraneveld, A. D., Garssen, J., Folkerts, G., & van’t Land, B. (2021). Human Milk Oligosaccharide 3′-GL Improves Influenza-Specific Vaccination Responsiveness and Immunity after Deoxynivalenol Exposure in Preclinical Models. Nutrients, 13(9), 3190. https://doi.org/10.3390/nu13093190