
Endothelium-Independent Relaxation of Vascular Smooth Muscle Induced by Persimmon-Derived Polyphenol Phytocomplex in Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of PJ and PLs Powder
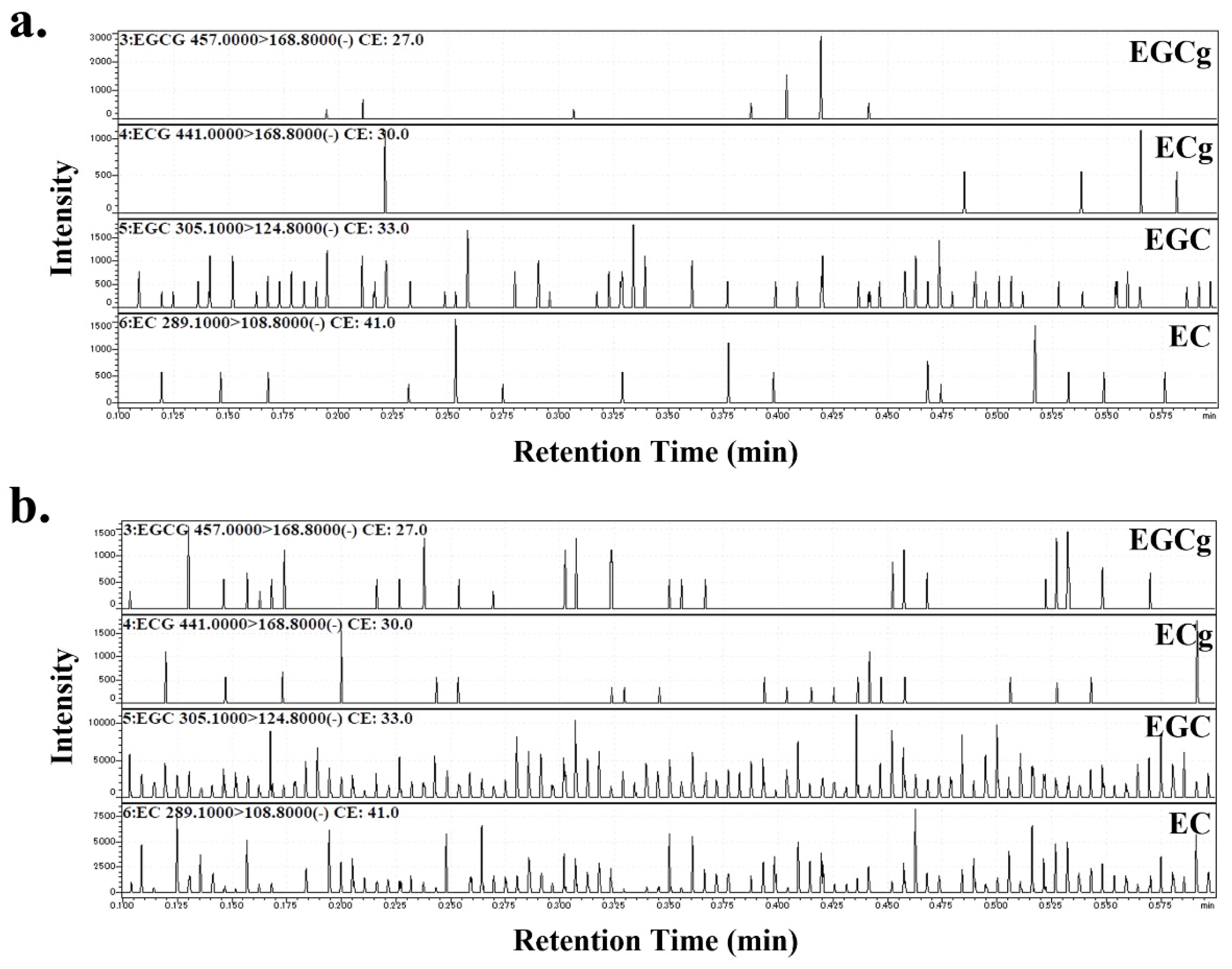
2.2. Estimation of Catechin Components in PJ and PLs
2.3. Animal Procedures
2.4. Preparation of Rings of Isolated Superior Mesenteric Artery
2.5. Tension Measurement
2.6. PJ- and PLs-Induced Vasorelaxation
2.7. Effect of PJ and PLs on Potassium Chloride- and Phenylephrine-Induced Vasocontraction
2.8. Statistical Analyses
2.9. Chemicals and Drugs
3. Results
3.1. Catechins Contained in PJ and PLs
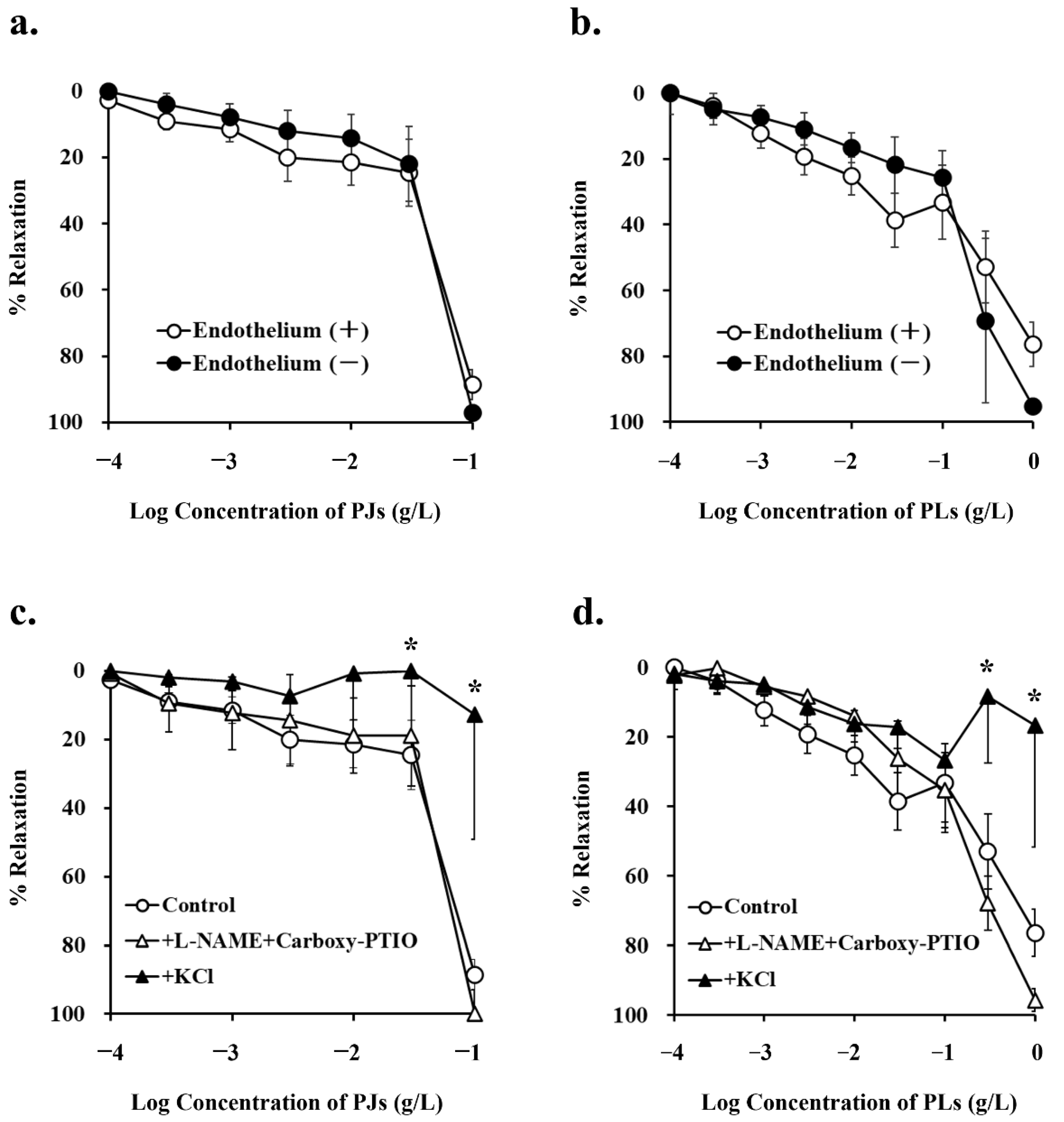
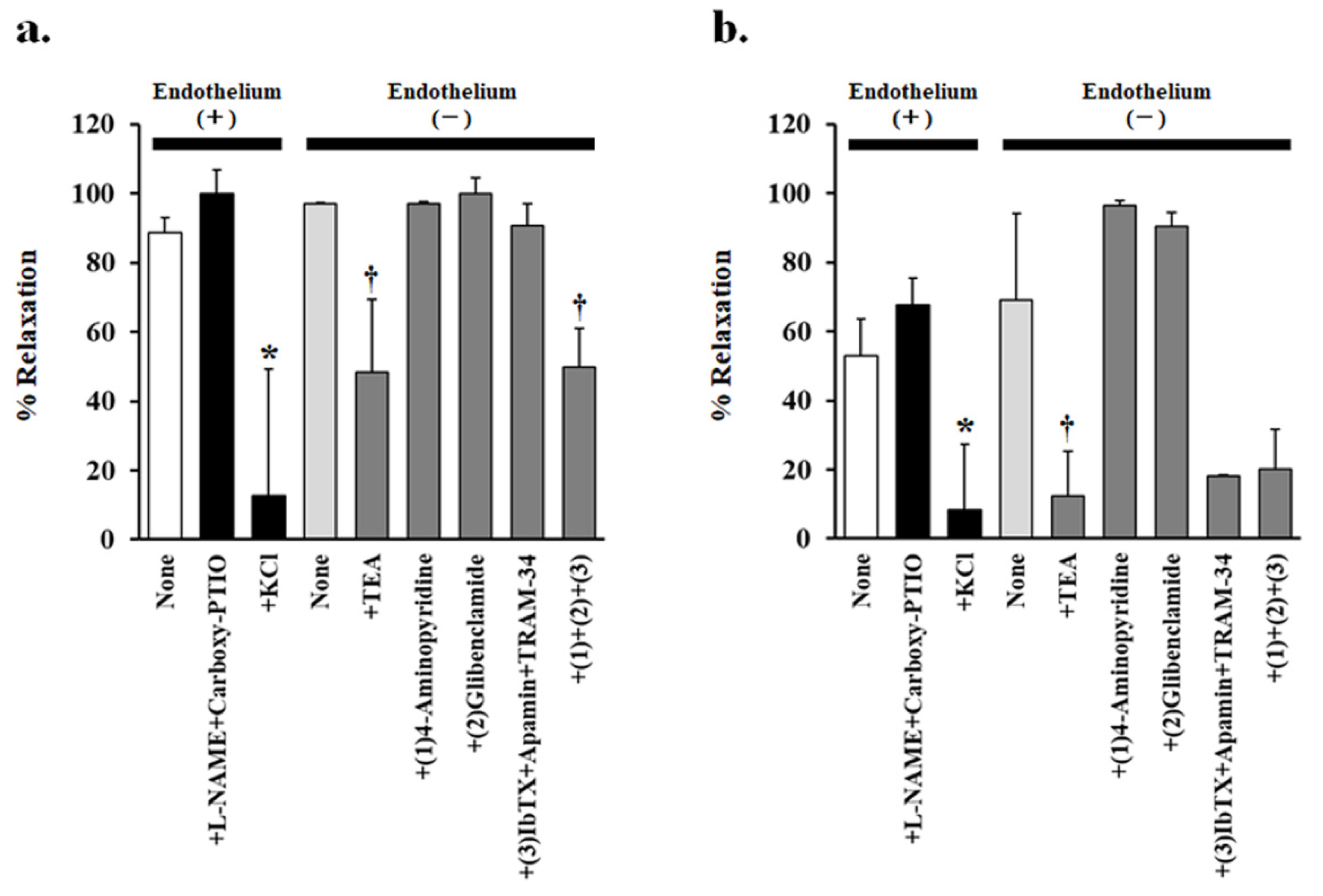
3.2. Endothelium-Dependent Vasorelaxation Induced by PJ and PLs
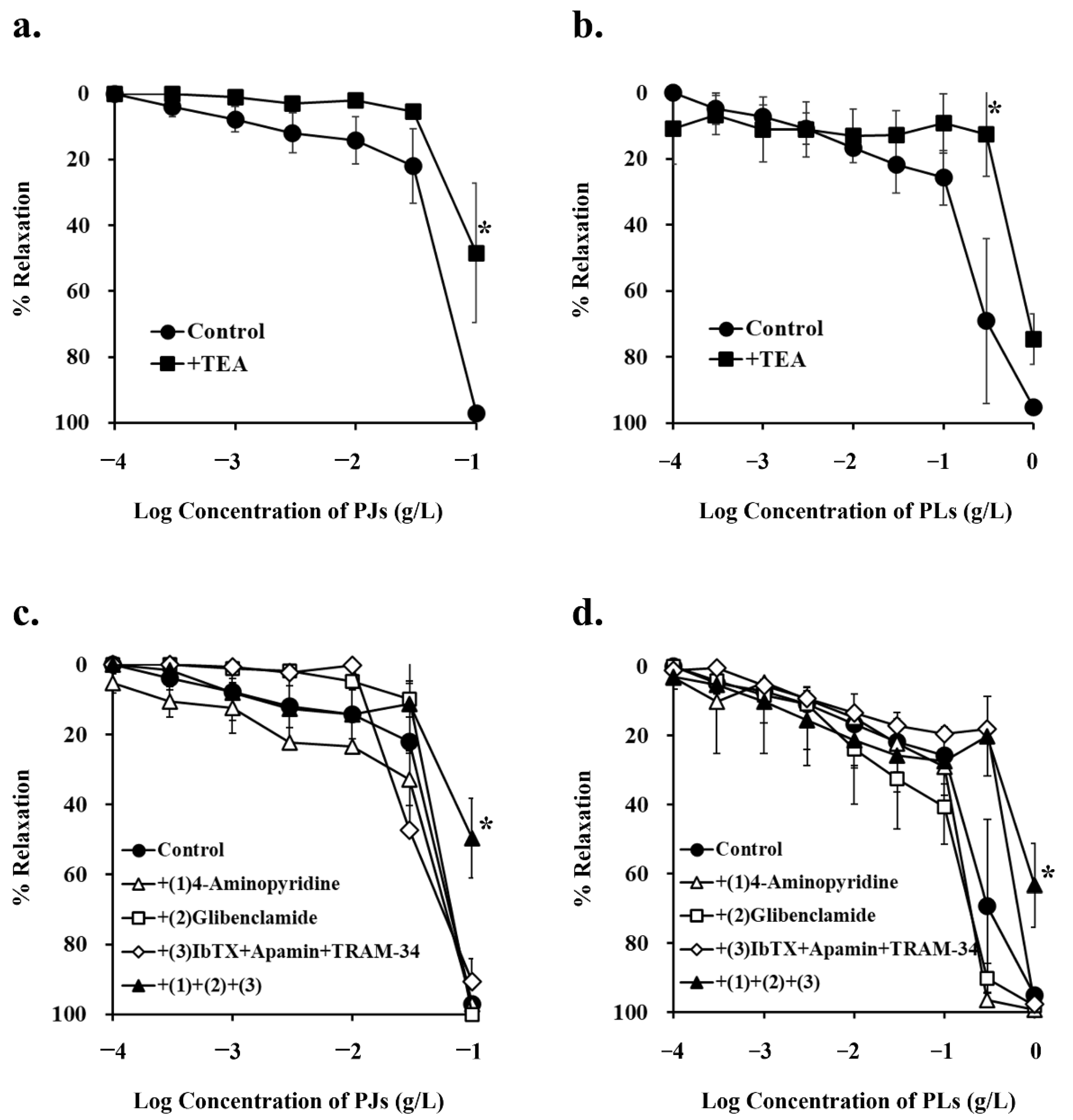
3.3. Effect of Potassium-Channel Inhibitors on PJ- and PLs-Induced Vasorelaxation
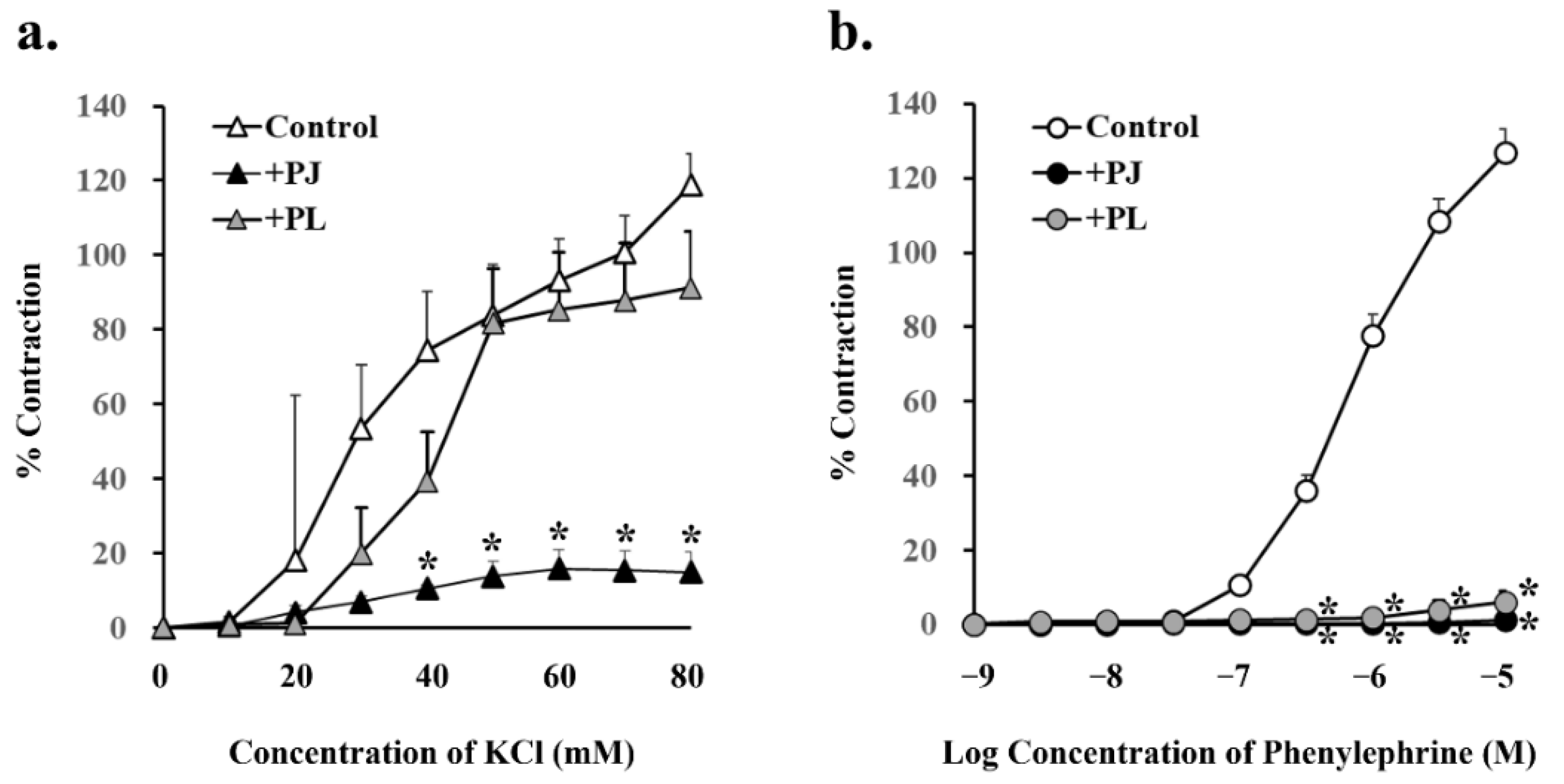
3.4. Effect of PJ and PLs on Potassium Chloride- and Phenylephrine-Induced Vasocontraction
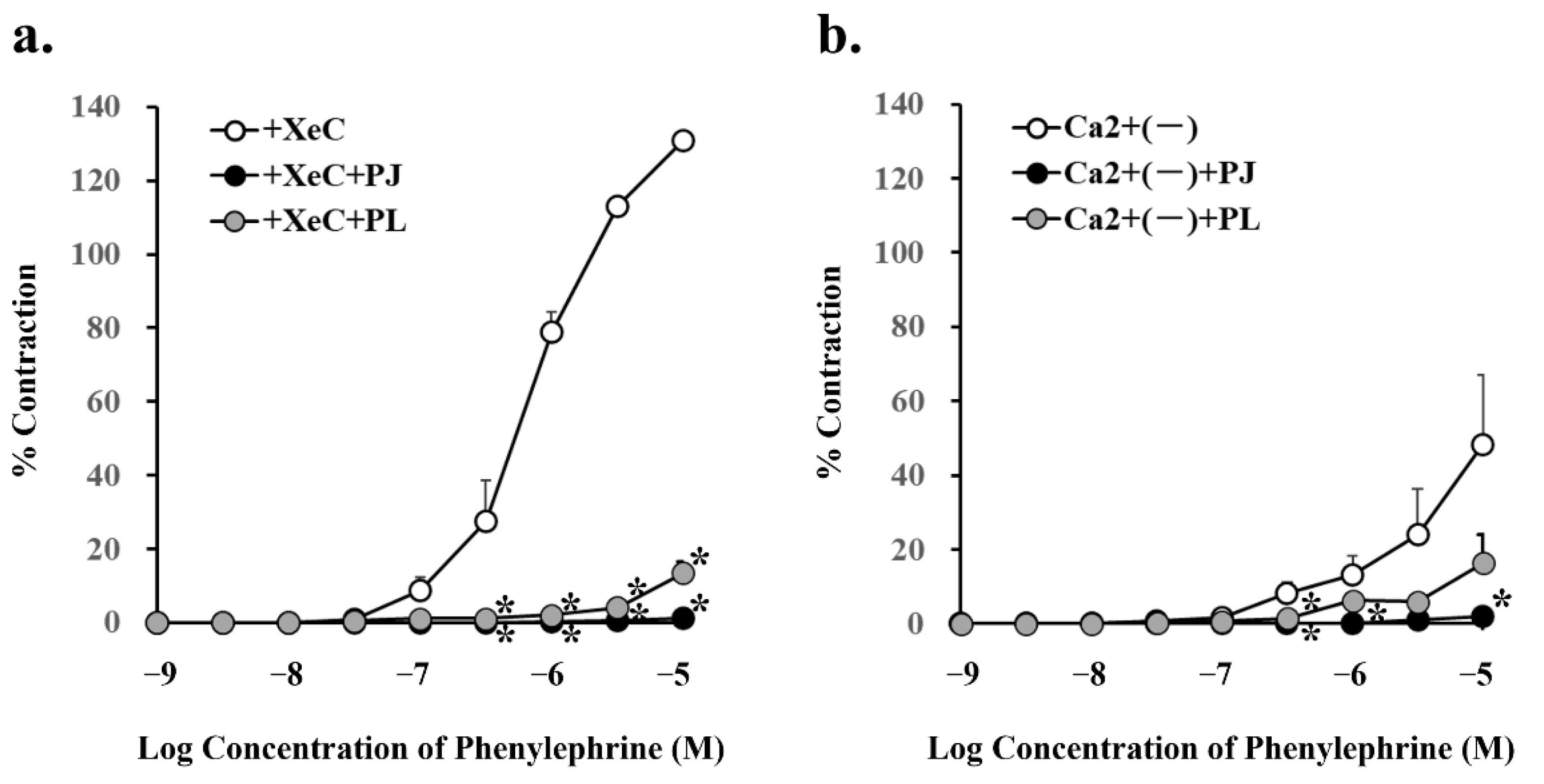
3.5. Effect of PJ and PLs on Phenylephrine-Induced Vasocontraction in the Presence of an Inhibitor of Intracellular Triphosphate-Mediated Ca2+ Release and in the Absence of Ca2+
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, N.; Mukhtar, H. Tea Polyphenols in Promotion of Human Health. Nutrients 2018, 11, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Hida, M.; Matsumoto, T.; Ikeuchi-Takahashi, Y.; Onishi, H.; Kobayashi, T. Effect of short-term polyphenol treatment on endothelial dysfunction and thromboxane A2 levels in streptozotocin-induced diabetic mice. Biol. Pharm. Bull. 2014, 37, 1056–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munir, K.M.; Chandrasekaran, S.; Gao, F.; Quon, M.J. Mechanisms for food polyphenols to ameliorate insulin resistance and endothelial dysfunction: Therapeutic implications for diabetes and its cardiovascular complications. Am. J. Physiol. Endocrinol. Metab. 2013, 305, 679–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dam, R.M.; Naidoo, N.; Landberg, R. Dietary flavonoids and the development of type 2 diabetes and cardiovascular diseases: Review of recent findings. Curr. Opin. Lipidol. 2013, 24, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, M.; Miguel, M.; Aleixandre, A. Beneficial effects of polyphenols on cardiovascular disease. Pharmacol. Res. 2013, 68, 125–131. [Google Scholar] [CrossRef]
- Sies, H. Polyphenols and health: Update and perspectives. Arch. Biochem. Biophys. 2010, 501, 2–5. [Google Scholar] [CrossRef]
- Andrade, A.C.; Cesena, F.H.; Consolim-Colombo, F.M.; Coimbra, S.R.; Benjó, A.M.; Krieger, E.M.; Luz, P.L. Short-term red wine consumption promotes differential effects on plasma levels of high-density lipoprotein cholesterol, sympathetic activity, and endothelial function in hypercholesterolemic, hypertensive, and healthy subjects. Clinics 2009, 64, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Chaves, A.A.; Joshi, M.S.; Coyle, C.M.; Brady, J.E.; Dech, S.J.; Schanbacher, B.L.; Baliga, R.; Basuray, A.; Bauer, J.A. Vasoprotective endothelial effects of a standardized grape product in humans. Vascul. Pharmacol. 2009, 50, 20–26. [Google Scholar] [CrossRef]
- Coimbra, S.R.; Lage, S.H.; Brandizzi, L.; Yoshida, V.; Da Luz, P.L. The action of red wine and purple grape juice on vascular reactivity is independent of plasma lipids in hypercholesterolemic patients. Braz. J. Med. Biol. Res. 2005, 38, 1339–1347. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yuan, H.; Zhang, C.; Guan, Y.; Wu, Y.; Ling, F.; Niu, Y.; Li, Y. Epigallocatechin gallate improves insulin resistance in HepG2 cells through alleviating inflammation and lipotoxicity. Diabetes Res. Clin. Pract. 2018, 142, 363–373. [Google Scholar] [CrossRef]
- Mi, Y.; Liu, X.; Tian, H.; Liu, H.; Li, J.; Qi, G.; Liu, X. EGCG stimulates the recruitment of brite adipocytes, suppresses adipogenesis and counteracts TNF-α-triggered insulin resistance in adipocytes. Food Funct. 2018, 9, 3374–3386. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K. Polyphenols Regulate Endothelial Functions and Reduce the Risk of Cardiovascular Disease. Curr. Pharm. Des. 2019, 25, 2443–2458. [Google Scholar] [CrossRef]
- Li, H.; Xia, N.; Hasselwander, S.; Daiber, A. Resveratrol and Vascular Function. Int. J. Mol. Sci. 2019, 20, 2155. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Watanabe, S.; Kawamura, R.; Taguchi, K.; Kobayashi, T. Epigallocatechin gallate attenuates ET-1-induced contraction in carotid artery from type 2 diabetic OLETF rat at chronic stage of disease. Life Sci. 2014, 118, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo, R.; Gil, D.; Miranda-Merchak, A.; Kalantzidis, G. Antihypertensive role of polyphenols. Adv. Clin. Chem. 2012, 58, 225–254. [Google Scholar] [CrossRef]
- Dal-Ros, S.; Bronner, C.; Schott, C.; Kane, M.O.; Chataigneau, M.; Schini-Kerth, V.B.; Chataigneau, T. Angiotensin II-induced hypertension is associated with a selective inhibition of endothelium-derived hyperpolarizing factor-mediated responses in the rat mesenteric artery. J. Pharmacol. Exp. Ther. 2009, 328, 478–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, C.J.; Kubo, E.; Fujii, K.; Nishimura, Y.; Kobuchi, S.; Ohkita, M.; Yoshimura, M.; Kiso, Y.; Matsumura, Y. Antihypertensive effect of French maritime pine bark extract (Flavangenol): Possible involvement of endothelial nitric oxide-dependent vasorelaxation. J. Hypertens. 2009, 27, 92–101. [Google Scholar] [CrossRef]
- Anselm, E.; Socorro, V.F.; Dal-Ros, S.; Schott, C.; Bronner, C.; Schini-Kerth, V.B. Crataegus special extract WS 1442 causes endothelium-dependent relaxation via a redox-sensitive Src- and Akt-dependent activation of endothelial NO synthase but not via activation of estrogen receptors. J. Cardiovasc. Pharmacol. 2009, 53, 253–260. [Google Scholar] [CrossRef]
- Andriambeloson, E.; Kleschyov, A.L.; Muller, B.; Beretz, A.; Stoclet, J.C.; Andriantsitohaina, R. Nitric oxide production and endothelium-dependent vasorelaxation induced by wine polyphenols in rat aorta. Br. J. Pharmacol. 1997, 120, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Rocha, A.P.; Carvalho, L.C.; Sousa, M.A.; Madeira, S.V.; Sousa, P.J.; Tano, T.; Schini-Kerth, V.B.; Resende, A.C.; Soares de Moura, R. Endothelium-dependent vasodilator effect of Euterpe oleracea Mart. (Açaí) extracts in mesenteric vascular bed of the rat. Vascul. Pharmacol. 2007, 46, 97–104. [Google Scholar] [CrossRef]
- Anselm, E.; Chataigneau, M.; Ndiaye, M.; Chataigneau, T.; Schini-Kerth, V.B. Grape juice causes endothelium-dependent relaxation via a redox-sensitive Src- and Akt-dependent activation of eNOS. Cardiovasc. Res. 2007, 73, 404–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndiaye, M.; Chataigneau, T.; Chataigneau, M.; Schini-Kerth, V.B. Red wine polyphenols induce EDHF-mediated relaxations in porcine coronary arteries through the redox-sensitive activation of the PI3-kinase/Akt pathway. Br. J. Pharmacol. 2004, 142, 1131–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, M.; Zhang, X.; Bai, M.; Wang, L. Persimmon leaf flavonoid promotes brain ischemic tolerance. Neural Regen. Res. 2013, 8, 2625–2632. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.F.; Gao, Y.S.; Xin, X.M. Hypoglycemic and hypocholesterolaemia effect of total flavonoids from persimmon leaf in alloxan-induced diabetic mice and its mechanism. Taishan Yixueyuan Xuebao 2009, 30, 245–247. [Google Scholar]
- Xin, N.; Feng, J.; Yao, B. Extraction, separation and pharmacological effects of total flavonoids from persimmon leaf. Zhongyiyao Xuebao 2007, 2, 49–51. [Google Scholar]
- Kawakami, K.; Aketa, S.; Sakai, H.; Watanabe, Y.; Nishida, H.; Hirayama, M. Antihypertensive and vasorelaxant effects of water-soluble proanthocyanidins from persimmon leaf tea in spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2011, 75, 1435–1439. [Google Scholar] [CrossRef] [Green Version]
- Iwase, K.; Torii, H. A colorimetric determination of tea tannin with ferrous tartrate. Tea Res. J. (Chagyo Kenkyu Hokoku) 1962, 19, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Usui, K.; Murata, T.; Fujita, Y.; Kamijo, Y.; Hanazawa, T.; Yoshizawa, T.; Funayama, M. Direct detection of the psychoactive substance MT-45 in human tissue samples by probe electrospray ionization-tandem mass spectrometry. Drug Test. Anal. 2018, 10, 1033–1038. [Google Scholar] [CrossRef]
- Wang, C.Y.; Li, Q.S.; Han, G.Z.; Zou, L.L.; Lv, L.; Zhou, Q.; Li, N. LC-MS/MS for simultaneous determination of four major active catechins of tea polyphenols in rat plasma and its application to pharmacokinetics. Chin. Herb. Med. 2010, 2, 289–296. [Google Scholar]
- Yuui, K.; Kudo, R.; Kasuda, S.; Hatake, K. Ethanol attenuates vasorelaxation via inhibition of inducible nitric oxide synthase in rat artery exposed to interleukin-1β. Hum. Exp. Toxicol. 2016, 35, 938–945. [Google Scholar] [CrossRef]
- Kudo, R.; Yuui, K.; Kasuda, S.; Nakata, M.; Imai, H.; Nakanishi, M.; Hatake, K. Effect of ethanol on capsaicin-induced nerve-mediated vasorelaxation in rat arteries. Rom. J. Legal Med. 2017, 25, 1–7. [Google Scholar] [CrossRef]
- Shimokawa, H.; Yasutake, H.; Fujii, K.; Owada, M.K.; Nakaike, R.; Fukumoto, Y.; Takayanagi, T.; Nagao, T.; Egashira, K.; Fujishima, M.; et al. The importance of the hyperpolarizing mechanism increases as the vessel size decreases in endothelium-dependent relaxations in rat mesenteric circulation. J. Cardiovasc. Pharmacol. 1996, 28, 703–711. [Google Scholar] [CrossRef]
- Nagao, T.; Illiano, S.; Vanhoutte, P.M. Heterogeneous distribution of endothelium-dependent relaxations resistant to NG-nitro-L-arginine in rats. Am. J. Physiol. 1992, 263, H1090–H1094. [Google Scholar] [CrossRef]
- Schini-Kerth, V.B.; Auger, C.; Kim, J.H.; Etienne-Selloum, N.; Chataigneau, T. Nutritional improvement of the endothelial control of vascular tone by polyphenols: Role of NO and EDHF. Pflugers. Arch. 2010, 459, 853–862. [Google Scholar] [CrossRef]
- Schini-Kerth, V.B.; Auger, C.; Etienne-Selloum, N.; Chataigneau, T. Polyphenol-induced endothelium-dependent relaxations role of NO and EDHF. Adv. Pharmacol. 2010, 60, 133–175. [Google Scholar] [CrossRef] [PubMed]
- López-Sepúlveda, R.; Gómez-Guzmán, M.; Zarzuelo, M.J.; Romero, M.; Sánchez, M.; Quintela, A.M.; Galindo, P.; O’Valle, F.; Tamargo, J.; Pérez-Vizcaíno, F.; et al. Red wine polyphenols prevent endothelial dysfunction induced by endothelin-1 in rat aorta: Role of NADPH oxidase. Clin. Sci. 2011, 120, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Reiter, C.N.; Kim, J.; Quon, M.J. Green tea polyphenol epigallocatechin gallate reduces endothelin-1 expression and secretion in vascular endothelial cells: Roles for AMP-activated protein kinase, Akt, and FOXO1. Endocrinology 2010, 151, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Kane, M.O.; Etienne-Selloum, N.; Madeira, S.F.; Sarr, M.; Walter, A.; Dal-Ros, S.; Schott, C.; Chataigneau, T.; Schini-Kerth, V.B. Endothelium-derived contracting factors mediate the Ang II-induced endothelial dysfunction in the rat aorta: Preventive effect of red wine polyphenols. Pflug. Arch. 2010, 459, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Kometani, T.; Takemori, K. Polyphenols from Persimmon Fruits as a Functional Foods Material. Nippon Shokuhin Kagaku Kogaku Kaishi 2016, 63, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, T.; Ito, S. The chemical structure of kaki-tannin from immature fruit of the persimmon (Diospyros kaki L.). Agric. Biol. Chem. 1978, 42, 1637–1643. [Google Scholar]
- Kawakami, K.; Aketa, S.; Nakanami, M.; Iizuka, S.; Hirayama, M. Major water-soluble polyphenols, proanthocyanidins, in leaves of persimmon (Diospyros kaki) and their alpha-amylase inhibitory activity. Biosci. Biotechnol. Biochem. 2010, 74, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.; Wessler, S.; Follmann, E.; Michaelis, W.; Düsterhöft, T.; Baumann, G.; Stangl, K.; Stangl, V. A constituent of green tea, epigallocatechin-3-gallate, activates endothelial nitric oxide synthase by a phosphatidylinositol-3-OH-kinase-, cAMP-dependent protein kinase-, and Akt-dependent pathway and leads to endothelial-dependent vasorelaxation. J. Biol. Chem. 2004, 279, 6190–6195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinko, M.; Jankovic, G.; Nenezic, D.; Milojevic, P.; Stojanovic, I.; Kanjuh, V.; Novakovic, A. (−)-Epicatechin-induced relaxation of isolated human saphenous vein: Roles of K+ and Ca2+ channels. Phytother. Res. 2018, 32, 267–275. [Google Scholar] [CrossRef]
- Luo, D.; Chen, X.; Zhu, X.; Liu, S.; Li, J.; Xu, J.; Zhao, J.; Ji, X. Pu-Erh Tea Relaxes the Thoracic Aorta of Rats by Reducing Intracellular Calcium. Front. Pharmacol. 2019, 10, 1430. [Google Scholar] [CrossRef] [PubMed]
- Desch, S.; Schmidt, J.; Kobler, D.; Sonnabend, M.; Eitel, I.; Sareban, M.; Rahimi, K.; Schuler, G.; Thiele, H. Effect of cocoa products on blood pressure: Systematic review and meta-analysis. Am. J. Hypertens. 2010, 23, 97–103. [Google Scholar] [CrossRef]
- Balzer, J.; Rassaf, T.; Heiss, C.; Kleinbongard, P.; Lauer, T.; Merx, M.; Heussen, N.; Gross, H.B.; Keen, C.L.; Schroeter, H.; et al. Sustained benefits in vascular function through flavanol-containing cocoa in medicated diabetic patients a double-masked, randomized, controlled trial. J. Am. Coll. Cardiol. 2008, 51, 2141–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassi, D.; Desideri, G.; Necozione, S.; Lippi, C.; Casale, R.; Properzi, G.; Blumberg, J.B.; Ferri, C. Blood pressure is reduced and insulin sensitivity increased in glucose-intolerant, hypertensive subjects after 15 days of consuming high-polyphenol dark chocolate. J. Nutr. 2008, 138, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cohn, J.S.; Harvey, I.; Le Cornu, K.A.; Ryder, J.J.; Hall, W.L.; Cassidy, A. Flavonoids, flavonoid-rich foods, and cardiovascular risk: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2008, 88, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Scheepens, A. Vascular action of polyphenols. Mol. Nutr. Food. Res. 2009, 53, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.; Urban, J.; Engelhardt, U.; Baumann, G.; Stangl, K.; Stangl, V. Green and black tea are equally potent stimuli of NO production and vasodilation: New insights into tea ingredients involved. Basic Res. Cardiol. 2009, 104, 100–110. [Google Scholar] [CrossRef]
- Duffy, S.J.; Keaney, J.F., Jr.; Holbrook, M.; Gokce, N.; Swerdloff, P.L.; Frei, B.; Vita, J.A. Short- and long-term black tea consumption reverses endothelial dysfunction in patients with coronary artery disease. Circulation 2001, 104, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, Y.; Yanagisawa, T.; Taira, N. BRL 38227 (levcromakalim)-induced hyperpolarization reduces the sensitivity to Ca2+ of contractile elements in canine coronary artery. Naunyn Schmiedebergs Arch. Pharmacol. 1993, 347, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Yanagisawa, T.; Yamagishi, T.; Taira, N. K+ channel-opening action and KRN2391-induced reduction of Ca2+ sensitivity of arterial smooth muscle. Arch. Int. Pharmacodyn. Ther. 1993, 326, 33–51. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitors Added | PJ (0.1 g/L) | PLs (0.3 g/L) | ||
|---|---|---|---|---|
| % Relaxation | p Value | % Relaxation | p Value | |
| Endothelium (+) | ||||
| none | 88.6 ± 4.39 | 58.4 ± 7.79 | ||
| +L-NAME + carboxy-PTIO | 100.0 ± 6.99 | 0.97 | 72.6 ± 6.58 | 0.93 |
| +KCl | 12.7 ± 36.5 * | 0.0002 | 8.39 ± 19.0 * | 0.05 |
| Endothelium (-) | ||||
| none | 97.1 ± 0.28 | 0.95 1 | 69.2 ± 25.0 | 0.91 1 |
| +TEA | 48.4 ± 21.1 † | 0.03 | 12.5 ± 12.9 † | 0.04 |
| +(1) 4-Aminopyridine | 97.0 ± 0.46 | 0.85 | 96.6 ± 1.44 | 0.99 |
| +(2) Glibenclamide | 100.0 ± 4.54 | 0.89 | 90.3 ± 4.35 | 0.98 |
| +(3) IbTX + apamin + TRAM-34 | 90.7 ± 6.48 | 0.74 | 18.2 ± 0.11 | 0.09 |
| +(1) + (2) + (3) | 49.7 ± 11.4 † | 0.03 | 20.2 ± 11.5 | 0.07 |
| Pretreatment | Potassium Chloride (80 mM) | Phenylephrine (10 µM) | ||
|---|---|---|---|---|
| % Contraction | p Value | % Contraction | p Value | |
| Control | 118.8 ± 8.46 | 126.9 ± 6.32 | ||
| +PJ (1 g/L) | 15.0 ± 5.28 * | 0.00002 | 1.2 ± 0.77 * | 0.000001 |
| +PLs (1 g/L) | 91.3 ± 15.1 | 0.10 | 6.1 ± 3.02 * | 0.000001 |
| Condition | Phenylephrine (10 µM) | |
|---|---|---|
| % Contraction | p Value | |
| + XeC (0.5 µM) | 130.8 ± 2.32 | |
| + XeC (0.5 µM) + PJ (1 g/L) | 1.2 ± 1.17 * | 0.0001 |
| + XeC (0.5 µM) + PLs (1 g/L) | 13.5 ± 2.91 * | 0.0002 |
| Ca2+ (-) | 48.2 ± 18.9 | |
| Ca2+ (-) + PJ (1 g/L) | 2.1 ± 1.10 * | 0.04 |
| Ca2+ (-) + PLs (1 g/L) | 16.4 ± 7.48 | 0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudo, R.; Yuui, K.; Kasuda, S. Endothelium-Independent Relaxation of Vascular Smooth Muscle Induced by Persimmon-Derived Polyphenol Phytocomplex in Rats. Nutrients 2022, 14, 89. https://doi.org/10.3390/nu14010089
Kudo R, Yuui K, Kasuda S. Endothelium-Independent Relaxation of Vascular Smooth Muscle Induced by Persimmon-Derived Polyphenol Phytocomplex in Rats. Nutrients. 2022; 14(1):89. https://doi.org/10.3390/nu14010089
Chicago/Turabian StyleKudo, Risa, Katsuya Yuui, and Shogo Kasuda. 2022. "Endothelium-Independent Relaxation of Vascular Smooth Muscle Induced by Persimmon-Derived Polyphenol Phytocomplex in Rats" Nutrients 14, no. 1: 89. https://doi.org/10.3390/nu14010089
APA StyleKudo, R., Yuui, K., & Kasuda, S. (2022). Endothelium-Independent Relaxation of Vascular Smooth Muscle Induced by Persimmon-Derived Polyphenol Phytocomplex in Rats. Nutrients, 14(1), 89. https://doi.org/10.3390/nu14010089