
Peptide−Calcium Chelate from Antler (Cervus elaphus) Bone Enhances Calcium Absorption in Intestinal Caco-2 Cells and D-gal-Induced Aging Mouse Model


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Bone Calcium Solution and Total Ash Determination
2.3. Peptide Extraction
2.4. MS Identification and Sequence Analysis of ABPs
2.5. Calcium Binding Capacity of ABPs
2.6. FTIR Measurement
2.7. SEM Measurement
2.8. The Caco-2 Cells Monolayers Establishment
2.9. Caco-2 Cells Monolayer Calcium Transport Studies
2.10. Animals
2.11. Feeding Procedures and Daily Observation of Physical Condition
2.12. Serum Biochemistry
2.13. Estimation of Visceral Indices
2.14. Micro-CT Analysis
2.15. Determination of Total Ash
2.16. Statistical Analyses
3. Results and Discussion
3.1. Antler Bone Calcium (AB−Ca)
3.2. Calcium-Binding Capacity and Amino Sequences Analysis of ABPs
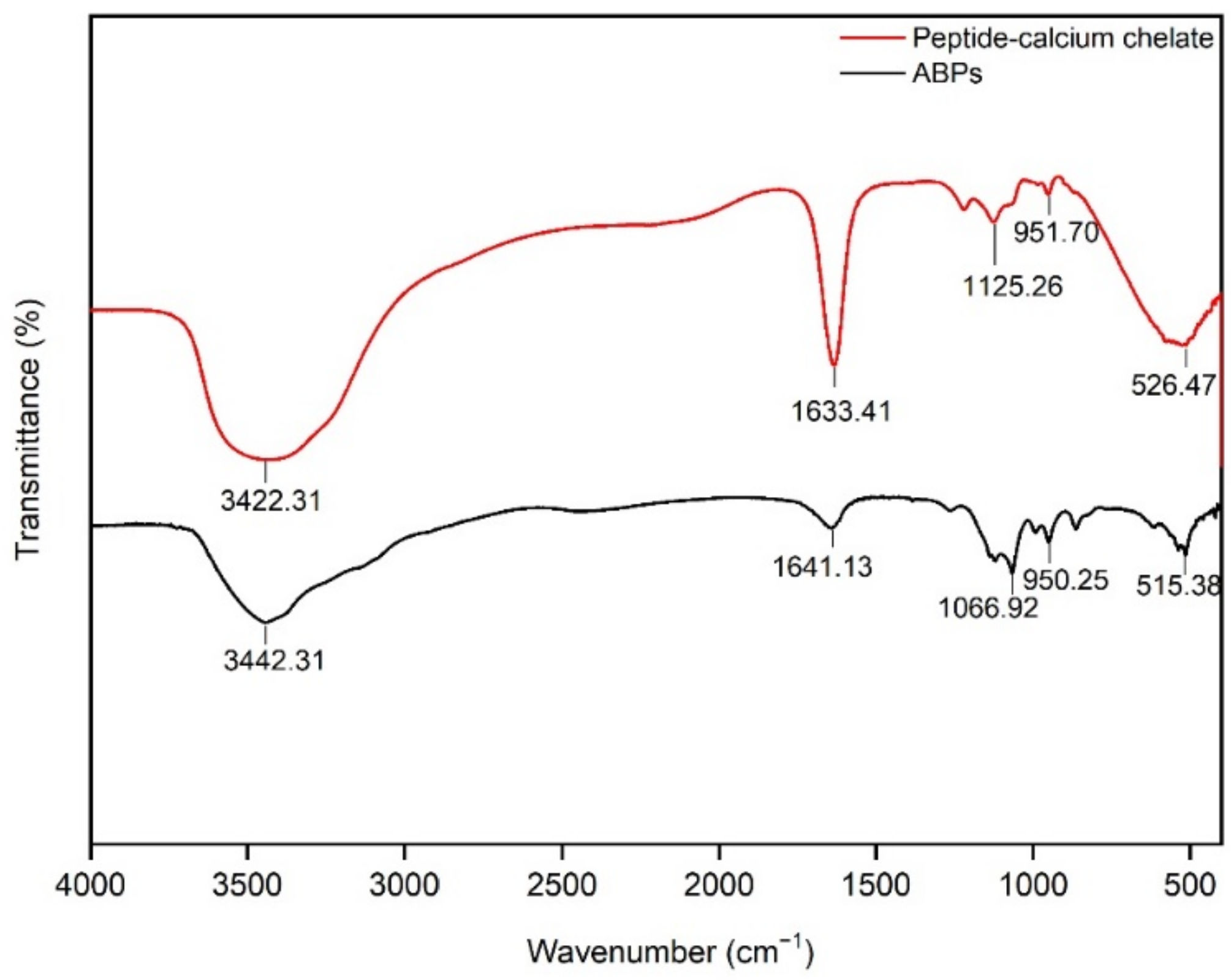
3.3. FTIR Measurement of Peptide–Calcium Chelate
3.4. SEM Measurement of Peptide–Calcium Chelate
3.5. ABPs Significantly Improved Calcium Transport in Intestinal Caco-2 Cells
3.6. No Substantial Variation in Weight Gain and Visceral Index Were Found among the Groups
3.7. ABPs + AB−Ca Group Had Significant Increase in Ca and PINP and Decrease in P, ALP, and CTX-1 Content in the Serum
3.8. Tibia Index and Ca Content Were Significantly Increased in ABPs + AB−Ca Group
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Laurent, M.R.; Dedeyne, L.; Dupont, J.; Mellaerts, B.; Dejaeger, M.; Gielen, E. Age-related bone loss and sarcopenia in men. Maturitas 2019, 122, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Cosman, F.; de Beur, S.J.; LeBoff, M.S.; Lewiecki, E.M.; Tanner, B.; Randall, S.; Lindsay, R. Clinician’s Guide to Prevention and Treatment of Osteoporosis. Osteoporos. Int. 2014, 25, 2359–2381. [Google Scholar] [CrossRef]
- Nuti, R.; Brandi, M.L.; Checchia, G.; Di Munno, O.; Dominguez, L.; Falaschi, P.; Fiore, C.E.; Iolascon, G.; Maggi, S.; Michieli, R.; et al. Guidelines for the management of osteoporosis and fragility fractures. Intern. Emerg. Med. 2019, 14, 85–102. [Google Scholar] [CrossRef]
- Coughlan, T.; Dockery, F. Osteoporosis and fracture risk in older people. Clin. Med. 2014, 14, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Clynes, M.A.; Harvey, N.C.; Curtis, E.M.; Fuggle, N.R.; Dennison, E.M.; Cooper, C. The epidemiology of osteoporosis. Br. Med. Bull. 2020, 133, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Rinonapoli, G.; Ruggiero, C.; Meccariello, L.; Bisaccia, M.; Ceccarini, P.; Caraffa, A. Osteoporosis in Men: A Review of an Underestimated Bone Condition. Int. J. Mol. Sci. 2021, 22, 2105. [Google Scholar] [CrossRef]
- Reid, I.R.; Bristow, S.M.; Bolland, M.J. Calcium supplements: Benefits and risks. J. Intern. Med. 2015, 278, 354–368. [Google Scholar] [CrossRef]
- Cristina, N.M.; Lucia, D. Nutrition and Healthy Aging: Prevention and Treatment of Gastrointestinal Diseases. Nutrients 2021, 13, 2337. [Google Scholar] [CrossRef]
- Maraka, S.; Kennel, K.A. Bisphosphonates for the prevention and treatment of osteoporosis. BMJ 2015, 351, h3783. [Google Scholar] [CrossRef]
- Barrett-Connor, E.; Stuenkel, C.A. Hormone replacement therapy (HRT)—Risks and benefits. Int. J. Epidemiol. 2001, 30, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Neer, R.M.; Arnaud, C.D.; Zanchetta, J.R.; Prince, R.; Gaich, G.A.; Reginster, J.Y.; Hodsman, A.B.; Eriksen, E.F.; Ish-Shalom, S.; Genant, H.K.; et al. Effect of parathyroid hormone (1-34) on fractures and bone mineral density in postmenopausal women with osteoporosis. N. Engl. J. Med. 2001, 344, 1434–1441. [Google Scholar] [CrossRef]
- Whitaker, M.; Guo, J.; Kehoe, T.; Benson, G. Bisphosphonates for Osteoporosis—Where Do We Go from Here? N. Engl. J. Med. 2012, 366, 2048–2051. [Google Scholar] [CrossRef]
- Hou, T.; Lu, Y.L.; Guo, D.J.; Liu, W.W.; Shi, W.; He, H. A pivotal peptide (Val-Ser-Glu-Glu) from duck egg white promotes calcium uptake and structure-activity relationship study. J. Funct. Foods 2018, 48, 448–456. [Google Scholar] [CrossRef]
- Jin, Y.G.; Fu, W.W.; Ma, M.H. Preparation and structure characterization of soluble bone collagen peptide chelating calcium. Afr. J. Biotechnol. 2011, 10, 10204–10211. [Google Scholar]
- Liu, G.; Sun, S.; Guo, B.; Miao, B.; Luo, Z.; Xia, Z.; Ying, D.; Liu, F.; Guo, B.; Tang, J.; et al. Bioactive peptide isolated from casein phosphopeptides promotes calcium uptake in vitro and in vivo. Food Funct 2018, 9, 2251–2260. [Google Scholar] [CrossRef]
- Liu, G.; Guo, B.Y.; Sun, S.W.; Luo, M.N.; Liu, F.; Miao, J.Y.; Tang, J.; Huang, Y.H.; Cao, Y.; Song, M.Y. Promoting the Calcium-Uptake Bioactivity of Casein Phosphopeptides in vitro and in vivo. Front. Nutr. 2021, 8, 743791. [Google Scholar] [CrossRef]
- Wu, W.M.; He, L.C.; Liang, Y.H.; Yue, L.L.; Peng, W.M.; Jin, G.F.; Ma, M.H. Preparation process optimization of pig bone collagen peptide-calcium chelate using response surface methodology and its structural characterization and stability analysis. Food Chem. 2019, 284, 80–89. [Google Scholar] [CrossRef]
- Malison, A.; Arpanutud, P.; Keeratipibul, S. Chicken foot broth byproduct: A new source for highly effective peptide-calcium chelate. Food Chem. 2021, 345, 128713. [Google Scholar] [CrossRef]
- Guo, L.D.; Harnedy, P.A.; O’Keeffe, M.B.; Zhang, L.; Li, B.F.; Hou, H.; FitzGerald, R.J. Fractionation and identification of Alaska pollock skin collagen-derived mineral chelating peptides. Food Chem. 2015, 173, 536–542. [Google Scholar] [CrossRef]
- Liu, B.T.; Zhuang, Y.L.; Sun, L.P. Identification and characterization of the peptides with calcium-binding capacity from tilapia (Oreochromis niloticus) skin gelatin enzymatic hydrolysates. J. Food Sci. 2020, 85, 114–122. [Google Scholar] [CrossRef]
- Wang, L.; Ding, Y.Y.; Zhang, X.X.; Li, Y.F.; Wang, R.; Luo, X.H.; Li, Y.N.; Li, J.; Chen, Z.X. Isolation of a novel calcium-binding peptide from wheat germ protein hydrolysates and the prediction for its mechanism of combination. Food Chem. 2018, 239, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, A.; Chen, Y.; Zhang, X.Y.; Li, S.H.; Chen, Y. Preparation of cucumber seed peptide-calcium chelate by liquid state fermentation and its characterization. Food Chem. 2017, 229, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Fordtran, J.S.; Locklear, T.W. Ionic constituents and osmolality of gastric and small-intestinal fluids after eating. Am. J. Dig. Dis. 1966, 11, 503–521. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.H.; Sung, C.H.; Chen, L.G.; Lai, Y.J.; Chang, W.S.; Sung, H.C.; Wang, C.C. Comparison of chemical compositions and osteoprotective effects of different sections of velvet antler. J. Ethnopharmacol. 2014, 151, 352–360. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Currey, J.D.; Ceacero, F.; Garcia, A.J.; Gallego, L.; Gomez, S. Does nutrition affect bone porosity and mineral tissue distribution in deer antlers? The relationship between histology, mechanical properties and mineral composition. Bone 2012, 50, 245–254. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Estevez, J.A.; Martinez, A.; Ceacero, F.; Garcia, A.; Gallego, L. Does chemical composition of antler bone reflect the physiological effort made to grow it? Bone 2007, 40, 1095–1102. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Kierdorf, H.; Gomez, S.; Luna, S.; Garcia, A.J.; Cappelli, J.; Perez-Serrano, M.; Perez-Barberia, J.; Gallego, L.; Kierdorf, U. Antlers—Evolution, development, structure, composition, and biomechanics of an outstanding type of bone. Bone 2019, 128, 115046. [Google Scholar] [CrossRef]
- Wu, F.; Li, H.; Jin, L.; Li, X.; Ma, Y.; You, J.; Li, S.; Xu, Y. Deer antler base as a traditional Chinese medicine: A review of its traditional uses, chemistry and pharmacology. J. Ethnopharmacol. 2013, 145, 403–415. [Google Scholar] [CrossRef]
- Kierdorf, U.; Stoffels, D.; Kierdorf, H. Element concentrations and element ratios in antler and pedicle bone of yearling red deer (Cervus elaphus) stags-a quantitative X-ray fluorescence study. Biol. Trace Element Res. 2014, 162, 124–133. [Google Scholar] [CrossRef]
- Gilbey, A.; Perezgonzalez, J.D. Health benefits of deer and elk velvet antler supplements: A systematic review of randomised controlled studies. N. Z. Med. J. 2012, 125, 80–86. [Google Scholar]
- Zhou, R.; Li, S.F. In vitro antioxidant analysis and characterisation of antler velvet extract. Food Chem. 2009, 114, 1321–1327. [Google Scholar] [CrossRef]
- Hung, Y.K.; Ho, S.T.; Kuo, C.Y.; Chen, M.J. In vitro effects of velvet antler water extracts from Formosan Sambar deer and red deer on barrier integrity in Caco-2 cell. Int. J. Med. Sci. 2021, 18, 1778–1785. [Google Scholar] [CrossRef]
- Zheng, K.X.; Li, Q.L.; Lin, D.O.; Zong, X.Y.; Luo, X.; Yang, M.; Yue, X.Q.; Ma, S.L. Peptidomic analysis of pilose antler and its inhibitory effect on triple-negative breast cancer at multiple sites. Food Funct. 2020, 11, 7481–7494. [Google Scholar] [CrossRef]
- Hou, F.J.; Chang, S.H.; Wen, Y.Y. Brief introduction to deer in China. Pratacultural Sci. 2003, 11, 47–50. [Google Scholar]
- Weaver, C.M.; Martin, B.R.; Costa, N.; Saleeb, F.Z.; Huth, P.J. Absorption of Calcium Fumarate Salts Is Equivalent to Other Calcium Salts When Measured in the Rat Model. J. Agr. Food Chem. 2002, 50, 4974–4975. [Google Scholar] [CrossRef]
- Sun, N.; Jin, Z.Q.; Li, D.M.; Yin, H.J.; Lin, S.Y. An Exploration of the Calcium-Binding Mode of Egg White Peptide, Asp-His-Thr-Lys-Glu, and In Vitro Calcium Absorption Studies of Peptide-Calcium Complex. J. Agr. Food Chem. 2017, 65, 9782–9789. [Google Scholar] [CrossRef]
- Wang, L.P.; Chen, Q.W.; Zhuang, S.Q.; Wen, Y.Y.; Cheng, W.Q.; Zeng, Z.J.; Jiang, T.; Tang, C.P. Effect of Anoectochilus roxburghii flavonoids extract on H2O2—Induced oxidative stress in LO2 cells and D-gal induced aging mice model. Ethnopharmacol. 2020, 254, 112670. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Huang, G.R.; Ren, Z.Y.; Jiang, J.X.; Chen, W.W. Purification of a Hepta-peptide with Iron Binding Activity from Shrimp Processing By-products Hydrolysates. Adv. J. Food Sci. Technol. 2012, 4, 207–212. [Google Scholar]
- Huang, G.; Ren, Z.; Jiang, J. Separation of Iron-Binding Peptides from Shrimp Processing By-products Hydrolysates. Food Bioprocess Technol. 2011, 4, 1527–1532. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, S.Q.; Song, H.D.; Li, B. Effect of Collagen Hydrolysates from Silver Carp Skin (Hypophthalmichthys molitrix) on Osteoporosis in Chronologically Aged Mice: Increasing Bone Remodeling. Nutrients 2018, 10, 1434. [Google Scholar] [CrossRef]
- Zhang, K.; Li, B.F.; Chen, Q.R.; Zhang, Z.H.; Zhao, X.; Hou, H. Functional Calcium Binding Peptides from Pacific Cod (Gadus macrocephalus) Bone: Calcium Bioavailability Enhancing Activity and Anti-Osteoporosis Effects in the Ovariectomy-Induced Osteoporosis Rat Model. Nutrients 2018, 10, 1325. [Google Scholar] [CrossRef]
- Jung, W.K.; Park, P.J.; Byun, H.G.; Moon, S.H.; Kim, S.K. Preparation of hoki (Johnius belengerii) bone oligophosphopeptide with a high affinity to calcium by carnivorous intestine crude proteinase. Food Chem. 2005, 91, 333–340. [Google Scholar] [CrossRef]
- Qian, J.Y.; Chen, W.; Zhang, W.M.; Zhang, H. Adulteration identification of some fungal polysaccharides with SEM, XRD, IR and optical rotation: A primary approach. Carbohydr. Polym. 2009, 78, 620–625. [Google Scholar] [CrossRef]
- Nara, M.; Morii, H.; Tanokura, M. Coordination to divalent cations by calcium-binding proteins studied by FTIR spectroscopy. Biochim. Biophys. Acta 2013, 1828, 2319–2327. [Google Scholar] [CrossRef]
- Doyle, B.B.; Bendit, E.G.; Blout, E.R. Infrared spectroscopy of collagen and collagen-like polypeptides. Biopolymers 1975, 14, 937–957. [Google Scholar] [CrossRef]
- Chen, D.; Liu, Z.Y.; Huang, W.Q.; Zhao, Y.H.; Dong, S.Y.; Zeng, M.Y. Purification and characterisation of a zinc-binding peptide from oyster protein hydrolysate. J. Funct. Foods 2013, 5, 689–697. [Google Scholar] [CrossRef]
- Zhang, Z.R.; Zhou, F.B.; Liu, X.L.; Zhao, M.M. Particulate nanocomposite from oyster (Crassostrea rivularis) hydrolysates via zinc chelation improves zinc solubility and peptide activity. Food Chem. 2018, 258, 269–277. [Google Scholar] [CrossRef]
- Sun, N.; Wang, Y.X.; Bao, Z.J.; Cui, P.B.; Wang, S.; Lin, S.Y. Calcium binding to herring egg phosphopeptides: Binding characteristics, conformational structure and intermolecular forces. Food Chem. 2020, 310, 125867. [Google Scholar] [CrossRef]
- Cao, Y.; Miao, J.Y.; Liu, G.; Luo, Z.; Xia, Z.M.; Liu, F.; Yao, M.F.; Cao, X.Q.; Sun, S.W.; Lin, Y.Y.; et al. Bioactive Peptides Isolated from Casein Phosphopeptides Enhance Calcium and Magnesium Uptake in Caco-2 Cell Monolayers. J. Agr. Food Chem. 2017, 65, 2307–2314. [Google Scholar] [CrossRef]
- Lin, Q.L.; Xu, Q.B.; Bai, J.; Wu, W.; Hong, H.; Wu, J.P. Transport of soybean protein-derived antihypertensive peptide LSW across Caco-2 monolayers. J. Funct. Foods 2017, 39, 96–102. [Google Scholar] [CrossRef]
- Xu, F.R.; Wang, L.F.; Ju, X.R.; Zhang, J.; Yin, S.; Shi, J.Y.; He, R.; Yuan, Q. Transepithelial Transport of YWDHNNPQIR and Its Metabolic Fate with Cytoprotection against Oxidative Stress in Human Intestinal Caco-2 Cells. J. Agr. Food Chem. 2017, 65, 2056–2065. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Wang, L.Y.; Jones, G.; Trang, H.; Yin, Y.G.; Liu, J.B. Optimized extraction of calcium malate from eggshell treated by PEF and an absorption assessment in vitro. Int. J. Biol. Macromol. 2012, 50, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Wang, C.; Ma, Z.; Shi, W.; Weiwei, L.; He, H. Desalted Duck Egg White Peptides: Promotion of Calcium Uptake and Structure Characterization. J. Agric. Food Chem. 2015, 63, 8170–8176. [Google Scholar] [CrossRef]
- Hou, T.; Liu, W.W.; Shi, W.; Ma, Z.L.; He, H. Desalted duck egg white peptides promote calcium uptake by counteracting the adverse effects of phytic acid. Food Chem. 2017, 219, 428–435. [Google Scholar] [CrossRef]
- Daniel, H.; Kottra, G. The proton oligopeptide cotransporter family SLC15 in physiology and pharmacology. Pflugers Arch. 2004, 447, 610–618. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, P.; Liu, Y.T.; Wu, Y.S.; Chen, Y.Q.; Guo, Y.C.; Zhang, S.W.; Zheng, X.F.; Zhou, L.Y.; Liu, W.Q.; et al. Alpha-ketoglutarate ameliorates age-related osteoporosis via regulating histone methylations. Nat. Commun. 2020, 11, 5596. [Google Scholar] [CrossRef]
- Wang, P.P.; Sun, H.X.; Liu, C.J.; Hu, M.H.; He, X.Q.; Yue, S.; Jiao, Z.Z.; Xiang, L. Racemic oleracein E increases the survival rate and attenuates memory impairment in D-galactose/NaNO(2)-induced senescent mice. Phytomedicine 2016, 23, 460–467. [Google Scholar] [CrossRef]
- Yang, J.; He, Y.; Zou, J.; Xu, L.; Fan, F.; Ge, Z. Effect of Polygonum Multiflorum Thunb on liver fatty acid content in aging mice induced by D-galactose. Lipids Health Dis. 2019, 18, 128. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, H.; Xu, M.; Wang, X.; Wang, C.; Lian, Y.; Mehmood, A.; Dai, H. Stevia residue extract ameliorates oxidative stress in d-galactose-induced aging mice via Akt/Nrf2/HO-1 pathway. J. Funct. Foods 2019, 52, 587–595. [Google Scholar] [CrossRef]
- Liu, H.Y.; Pei, X.L.; Wang, J.; Zhou, Y.; Wang, L.M.; Qi, B. Effect of loach paste on the liver and immune organs of D-galactose-induced ageing mice. Food Agr. Immunol. 2018, 29, 316–331. [Google Scholar] [CrossRef]
- Mo, Z.Z.; Liu, Y.H.; Li, C.L.; Xu, L.Q.; Wen, L.L.; Xian, Y.F.; Lin, Z.X.; Zhan, J.Y.; Chen, J.N.; Xu, F.F.; et al. Protective Effect of SFE-CO2 of Ligusticum chuanxiong Hort Against d-Galactose-Induced Injury in the Mouse Liver and Kidney. Rejuvenation Res. 2017, 20, 231–243. [Google Scholar] [CrossRef]
- Nam, S.M.; Seo, M.; Seo, J.S.; Rhim, H.; Nahm, S.S.; Cho, I.H.; Chang, B.J.; Kim, H.J.; Choi, S.H.; Nah, S.Y. Ascorbic Acid Mitigates D-galactose-Induced Brain Aging by Increasing Hippocampal Neurogenesis and Improving Memory Function. Nutrients 2019, 11, 176. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Guo, Y.; Xu, L.; Wang, H. Phlorizin exerts potent effects against aging induced by D-galactose in mice and PC12 cells. Food Funct. 2021, 12, 2148–2160. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Xin, J.L.; Zhang, X.P.; Fu, Q.; Zhang, Y.; Zhou, Q.L. The anti-osteoporotic effect of velvet antler polypeptides from Cervus elaphus Linnaeus in ovariectomized rats. J. Ethnopharmacol. 2013, 150, 181–186. [Google Scholar] [CrossRef]
- Zhang, H.; Wanwimolruk, S.; Coville, P.F.; Schofield, J.C.; Williams, G.; Haines, S.R.; Suttie, J.M. Toxicological evaluation of New Zealand deer velvet powder. Part I: Acute and subchronic oral toxicity studies in rats. Food Chem. Toxicol. 2000, 38, 985–990. [Google Scholar] [CrossRef]
- Peng, Z.; Hou, H.; Zhang, K.; Li, B.F. Effect of calcium-binding peptide from Pacific cod (Gadus macrocephalus) bone on calcium bioavailability in rats. Food Chem. 2017, 221, 373–378. [Google Scholar] [CrossRef]
- Wemeau, J.L.; Gautier, J.F. Management of type 2 diabetes: New drugs, new recommendations. Presse Med. 2013, 42, 828–829. [Google Scholar] [CrossRef]
- Goldsmith, R.S.; Jowsey, J.; Dube, W.J.; Riggs, B.L.; Arnaud, C.D.; Kelly, P.J. Effects of phosphorus supplementation on serum parathyroid hormone and bone morphology in osteoporosis. J. Clin. Endocrinol. Metab. 1976, 43, 523–532. [Google Scholar] [CrossRef]
- Morgan, E.N.; Alsharidah, A.S.; Mousa, A.M.; Edrees, H.M. Irisin Has a Protective Role against Osteoporosis in Ovariectomized Rats. Biomed Res. Int. 2021, 2021, 5570229. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, T.; Tang, H.; Ren, Z.H.; Li, Q.W.; Jia, J.; Chen, H.Y.; Fu, J.; Ding, S.C.; Hao, Q.; et al. Ganoderma lucidum Immune Modulator Protein rLZ-8 Could Prevent and Reverse Bone Loss in Glucocorticoids-Induced Osteoporosis Rat Model. Front. Pharmacol. 2020, 11, 731. [Google Scholar] [CrossRef]
- Gossiel, F.; Ugur, A.; Peel, N.F.A.; Walsh, J.S.; Eastell, R. The clinical utility of TRACP-5b to monitor anti-resorptive treatments of osteoporosis. Osteoporos. Int. 2022, 33, 1357–1363. [Google Scholar] [CrossRef]
- Wang, W.; Huang, C.Y.; Wang, Z.P.; Xu, S.S.; Qian, T.Y.; Chen, Y.D.; Wu, W.G. Serum C-C Motif Ligand 11/Eotaxin-1 May Serve as a Candidate Biomarker for Postmenopausal Osteoporosis. J. Med. Biochem. 2019, 38, 353–360. [Google Scholar] [CrossRef]
- Guo, D.; He, H.; Zhao, M.; Zhang, G.; Hou, T. Desalted duck egg white peptides promoted osteogenesis via wnt/beta-catenin signal pathway. J. Food Sci. 2020, 85, 834–842. [Google Scholar] [CrossRef]
- Yu, Y.J.; Wu, J.K.; Li, J.; Liu, Y.Z.; Zheng, X.Y.; Du, M.Z.; Zhou, L.M.; Yang, Y.J.; Luo, S.Y.; Hu, W.J.; et al. Cycloastragenol prevents age-related bone loss: Evidence in D-galactose-treated and aged rats. Biomed. Pharmacother. 2020, 128, 110304. [Google Scholar] [CrossRef]
- Sun, X.; Ruan, S.; Zhuang, Y.; Sun, L. Anti-osteoporosis effect and purification of peptides with high calcium-binding capacity from walnut protein hydrolysates. Food Funct. 2021, 12, 8454–8466. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Sequences | Molecular Weight (Da) |
|---|---|
| Gly-Pro-Gly-Ser-Pro-Gly-Pro | 567.27 |
| Gly-Asp-Gln-Gly-Val-Pro-Gly | 628.28 |
| Gly-Pro-Ala-Gly-Pro-Gly-Pro-Pro | 648.32 |
| Gly-Ala-Pro-Gly-Pro-Ser-Gly-Pro | 680.31 |
| Gly-Pro-Gly-Pro-Ile-Gly-Asn | 707.36 |
| Gly-Ser-Pro-Pro-Ala-Thr-Ser-Cys | 760.31 |
| Gly-Pro-Ala-Gly-Pro-Pro-Gly-Ala-Pro | 761.37 |
| Gly-Glu-Pro-Gly-Lys-Gln-Gly-Pro | 768.38 |
| Gly-Ala-Pro-Gly-Pro-Ser-Pro-Gly-Pro | 777.37 |
| Gly-Ser-Pro-Pro-Gly-Glu-Gly-Ala-Pro | 809.36 |
| Gly-Ala-Pro-Gly-Pro-Pro-Ser-Gly-Gly-Pro | 834.39 |
| Gly-Glu-Pro-Gly-Pro-Glu-Gly-Pro-Ala-Gly | 866.38 |
| Gly-Glu-Arg-Gly-Glu-Gln-Gly-Ala-Pro | 899.41 |
| Gly-Pro-Pro-Gly-Glu-Pro-Gly-Pro-Pro-Gly-Pro-Pro-Gly-Pro | 1208.58 |
| Gly-Pro-Glu-Gly-Pro-Pro-Gly-Glu-Pro-Gly-Pro-Pro-Gly-Pro | 1240.57 |
| Gly-Pro-Glu-Gly-Pro-Pro-Gly-Glu-Pro-Gly-Pro-Pro-Gly-Pro-Pro | 1337.63 |
| Gly-Asp-Ile-Gly-Pro-Pro-Gly-Pro-Gln-Gly-Pro-Pro-Gly-Pro | 1241.60 |
| Gly-Glu-Val-Gly-Gln-Ile-Gly-Pro-Arg-Gly-Glu-Asp-Gly-Pro-Glu-Gly-Pro | 1649.76 |
| Measurements | NC | CaCl2 | AB − Ca | ABPs + AB − Ca |
|---|---|---|---|---|
| Serum biochemistry | ||||
| ALP (U/L) | 12.95 ± 2.92 a | 11.79 ± 1.83 a | 6.56 ± 0.84 b | 5.53 ± 0.40 b |
| Ca (μmol/dL) | 38.09 ± 3.74 a | 39.68 ± 5.16 a | 40.07 ± 1.67 a | 63.59 ± 6.12 b |
| P (mmol/L) | 3.17 ± 0.13 a | 3.17 ± 0.09 a | 3.25 ± 0.52 a | 2.48 ± 0.13 b |
| PINP (ng/mL) | 3.30 ± 0.56 a | 3.23 ± 0.19 a | 4.13 ± 0.05 b | 4.65 ± 0.33 b |
| CTX-1 (pg/mL) | 218.24 ± 15.77 a | 205.60 ± 10.66 a | 172.70 ± 6.72 b | 160.99 ± 1.07 b |
| Tibia physicochemical indices | ||||
| Length (mm) | 19.65 ± 1.20 a | 19.85 ± 0.55 a | 20.10 ± 0.11 a | 19.68 ± 0.49 a |
| Dry bone weight (g) | 0.045 ± 0.008 a | 0.048 ± 0.007 a | 0.050 ± 0.004 a | 0.045 ± 0.002 a |
| Ca/Ash bone (μg/mg) | 387.18 ± 2.57 a | 386.14 ± 0.10 a | 387.64 ± 1.71 a | 398.92 ± 2.55 b |
| BMD (mg/cm2) | 531.38 ± 34.51 a | 570.37 ± 2.82 a | 599.92 ± 34.01 a | 643.51 ± 57.33 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhai, X.; Fang, J.; Wu, H.; Cheng, Y.; Gao, Y.; Chen, X.; Zheng, S.; Liu, S.; Hao, L. Peptide−Calcium Chelate from Antler (Cervus elaphus) Bone Enhances Calcium Absorption in Intestinal Caco-2 Cells and D-gal-Induced Aging Mouse Model. Nutrients 2022, 14, 3738. https://doi.org/10.3390/nu14183738
Wang Z, Zhai X, Fang J, Wu H, Cheng Y, Gao Y, Chen X, Zheng S, Liu S, Hao L. Peptide−Calcium Chelate from Antler (Cervus elaphus) Bone Enhances Calcium Absorption in Intestinal Caco-2 Cells and D-gal-Induced Aging Mouse Model. Nutrients. 2022; 14(18):3738. https://doi.org/10.3390/nu14183738
Chicago/Turabian StyleWang, Zhaoguo, Xiaorui Zhai, Jiayuan Fang, Hongyan Wu, Yunyun Cheng, Yuan Gao, Xi Chen, Shuo Zheng, Songcai Liu, and Linlin Hao. 2022. "Peptide−Calcium Chelate from Antler (Cervus elaphus) Bone Enhances Calcium Absorption in Intestinal Caco-2 Cells and D-gal-Induced Aging Mouse Model" Nutrients 14, no. 18: 3738. https://doi.org/10.3390/nu14183738
APA StyleWang, Z., Zhai, X., Fang, J., Wu, H., Cheng, Y., Gao, Y., Chen, X., Zheng, S., Liu, S., & Hao, L. (2022). Peptide−Calcium Chelate from Antler (Cervus elaphus) Bone Enhances Calcium Absorption in Intestinal Caco-2 Cells and D-gal-Induced Aging Mouse Model. Nutrients, 14(18), 3738. https://doi.org/10.3390/nu14183738