
Myo-Inositol Moderates Glucose-Induced Effects on Human Placental 13C-Arachidonic Acid Metabolism

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
1.1. Arachidonic Acid in Pregnancy
1.2. Myo-Inositol as a Potential Treatment for Disordered Placental Lipid Metabolism
1.3. Placental AA Metabolism
2. Methods and Materials
2.1. Placenta Collection and Placental Explant Culture
2.2. LC-MS/MS Methodology
2.3. Data and Statistical Analysis
2.4. Analysis of Data from Snap Frozen Placenta
3. Results
3.1. Placental Incorporation of Exogenous 13C-AA into 13C-AA Labeled Lipids
3.2. Association of Maternal Glycemia and BMI with Placental 13C-AA Incorporation under Culture with Physiological Glucose (5 mM)
3.3. Effect of In Vitro Glucose Treatment on Placental 13C-AA Lipid Metabolism
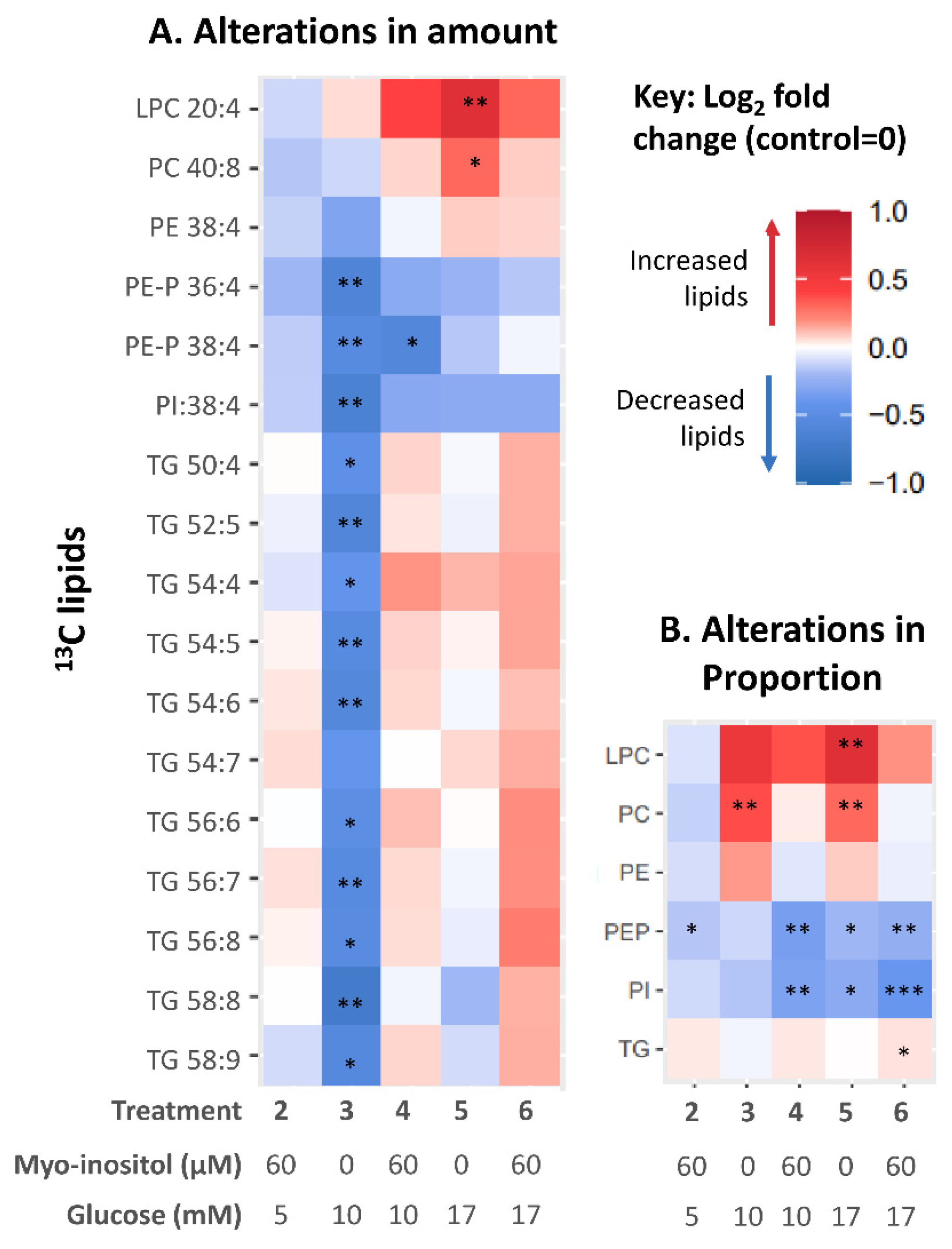
3.4. Effect of In Vitro Myo-Inositol or Combined Glucose and Myo-Inositol Treatment on Placental 13C-AA Lipids
3.5. Association of Prior Maternal Fasting Glycemia Exposure In Vivo with Alterations in Placental AA Lipids Induced by Glucose and Myoinositol Treatment In Vitro
3.6. Supporting Evidence from a Separate Cohort: Endogenous 12C-AA Lipids in Snap-Frozen Placental Biopsies
4. Discussion
4.1. Maternal Glycemia and Placental AA-Lipid Compartmentalization and Bioavailability
4.2. Glucose-Induced Changes in Placental AA-Lipids In Vitro
4.3. Placental Programming by Prior Maternal Glycemia
4.4. Myo-Inositol as a Moderator of Placental AA Metabolism
4.5. Clinical Implications
5. Strengths and Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bitsanis, D.; Crawford, M.A.; Moodley, T.; Holmsen, H.; Ghebremeskel, K.; Djahanbakhch, O. Arachidonic Acid Predominates in the Membrane Phosphoglycerides of the Early and Term Human Placenta. J. Nutr. 2005, 135, 2566–2571. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, T.; Winkler, L.; Möller, U.; Schubert, H.; Goetze, E. Synthesis of arachidonic acid in the human placenta in vitro. Neonatology 1979, 35, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Chambaz, J.; Ravel, D.; Manier, M.-C.; Pepin, D.; Mulliez, N.; Bereziat, G. Essential fatty acids interconversion in the human fetal liver. Neonatology 1985, 47, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M. Placental delivery of arachidonic and docosahexaenoic acids: Implications for the lipid nutrition of preterm infants. Am. J. Clin. Nutr. 2000, 71, 275S–284S. [Google Scholar] [CrossRef] [PubMed]
- Ghebremeskel, K.; Thomas, B.; Lowy, C.; Min, Y.; Crawford, M.A. Type 1 diabetes compromises plasma arachidonic and docosahexaenoic acids in newborn babies. Lipids 2004, 39, 335–342. [Google Scholar] [CrossRef]
- Min, Y.; Lowy, C.; Ghebremeskel, K.; Thomas, B.; Offley-Shore, B.; Crawford, M. Unfavorable effect of type 1 and type 2 diabetes on maternal and fetal essential fatty acid status: A potential marker of fetal insulin resistance. Am. J. Clin. Nutr. 2005, 82, 1162–1168. [Google Scholar] [CrossRef]
- Thomas, B.; Ghebremeskel, K.; Lowy, C.; Offley-Shore, B.; Crawford, M.A. Plasma fatty acids of neonates born to mothers with and without gestational diabetes. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 335–341. [Google Scholar] [CrossRef]
- Kuhn, D.C.; Botti, J.J.; Cherouny, P.H.; Demers, L.M. Eicosanoid production and transfer in the placenta of the diabetic pregnancy. Prostaglandins 1990, 40, 205–215. [Google Scholar] [CrossRef]
- Jawerbaum, A.; Catafau, J.R.; González, E.T.; Novaro, V.; Gómez, G.; Gelpí, E.; Gimeno, M.A.F. Eicosanoid production by placental and amnion tissues from control and non-insulin-dependent diabetic rats. Influence of oxytocin in the incubating medium. Prostaglandins Leukot. Essent. Fat. Acids 1997, 56, 425–429. [Google Scholar] [CrossRef]
- White, V.; Jawerbaum, A.; Sinner, D.; Pustovrh, C.; Capobianco, E.; González, E. Oxidative stress and altered prostanoid production in the placenta of streptozotocin-induced diabetic rats. Reprod. Fertil. Dev. 2002, 14, 117–123. [Google Scholar] [CrossRef]
- Pearson, T.; Warren, A.Y.; Barrett, D.A.; Khan, R.N. Detection of EETs and HETE-generating cytochrome P-450 enzymes and the effects of their metabolites on myometrial and vascular function. Am. J. Physiol.-Endocrinol. Metab. 2009, 297, E647–E656. [Google Scholar] [CrossRef] [PubMed]
- Uhl, O.; Demmelmair, H.; Segura, M.T.; Florido, J.; Rueda, R.; Campoy, C.; Koletzko, B. Effects of obesity and gestational diabetes mellitus on placental phospholipids. Diabetes Res. Clin. Pract. 2015, 109, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Bitsanis, D.; Ghebremeskel, K.; Moodley, T.; Crawford, M.A.; Djahanbakhch, O. Gestational diabetes mellitus enhances arachidonic and docosahexaenoic acids in placental phospholipids. Lipids 2006, 41, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Godhamgaonkar, A.A.; Wadhwani, N.S.; Joshi, S.R. Exploring the role of LC-PUFA metabolism in pregnancy complications. Prostaglandins Leukot. Essent. Fat. Acids 2020, 163, 102203. [Google Scholar] [CrossRef]
- Clements, R., Jr. The metabolism of myo-inositol by the human kidney. J. Lab. Clin. Med. 1979, 93, 210–219. [Google Scholar]
- Holub, B.J. Metabolism and function of myo-inositol and inositol phospholipids. Annu. Rev. Nutr. 1986, 6, 563–597. [Google Scholar] [CrossRef]
- Watkins, O.C.; Yong, H.E.J.; Sharma, N.; Chan, S.Y. A review of the role of inositols in conditions of insulin dysregulation and in uncomplicated and pathological pregnancy. Crit. Rev. Food Sci. Nutr. 2022, 62, 1626–1673. [Google Scholar] [CrossRef]
- Chan, K.Y.; Wong, M.M.H.; Pang, S.S.H.; Lo, K.K.H. Dietary supplementation for gestational diabetes prevention and management: A meta-analysis of randomized controlled trials. Arch. Gynecol. Obstet. 2021, 303, 1381–1391. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Barton, S.J.; El-Heis, S.; Kenealy, T.; Nield, H.; Baker, P.N.; Chong, Y.S.; Cutfield, W.; Chan, S.-Y. Myo-Inositol, Probiotics, and Micronutrient Supplementation From Preconception for Glycemia in Pregnancy: NiPPeR International Multicenter Double-Blind Randomized Controlled Trial. Diabetes Care 2021, 44, 1091–1099. [Google Scholar] [CrossRef]
- Zhang, H.; Lv, Y.; Li, Z.; Sun, L.; Guo, W. The efficacy of myo-inositol supplementation to prevent gestational diabetes onset: A meta-analysis of randomized controlled trials. J. Matern-Fetal Neonatal Med. 2019, 32, 2249–2255. [Google Scholar] [CrossRef]
- Chu, A.H.Y.; Tint, M.T.; Chang, H.F.; Wong, G.; Yuan, W.L.; Tull, D.; Nijagal, B.; Narayana, V.K.; Meikle, P.J.; Chang, K.T.E.; et al. High placental inositol content associated with suppressed pro-adipogenic effects of maternal glycaemia in offspring: The GUSTO cohort. Int. J. Obes. 2021, 45, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Groenen, P.M.; Merkus, H.M.; Sweep, F.C.; Wevers, R.A.; Janssen, F.S.; Steegers-Theunissen, R.P. Kinetics of myo-inositol loading in women of reproductive age. Ann. Clin. Biochem. 2003, 40, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Cogram, P.; Tesh, S.; Tesh, J.; Wade, A.; Allan, G.; Greene, N.D.; Copp, A.J. D-chiro-inositol is more effective than myo-inositol in preventing folate-resistant mouse neural tube defects. Hum. Reprod. 2002, 17, 2451–2458. [Google Scholar] [CrossRef]
- Baker, L.; Piddington, R.; Goldman, A.; Egler, J.; Moehring, J. Myo-inositol and prostaglandins reverse the glucose inhibition of neural tube fusion in cultured mouse embryos. Diabetologia 1990, 33, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Zeitler, P.; Wu, Y.Q.; Handwerger, S. Melittin stimulates phosphoinositide hydrolysis and placental lactogen release: Arachidonic acid as a link between phospholipase A2 and phospholipase C signal-transduction pathways. Life Sci. 1991, 48, 2089–2095. [Google Scholar] [CrossRef]
- Group, H.S.C.R. Hyperglycaemia and Adverse Pregnancy Outcome (HAPO) Study: Associations with maternal body mass index. BJOG Int. J. Obstet. Gynaecol. 2010, 117, 575–584. [Google Scholar]
- Association, A.D. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36, S67. [Google Scholar] [CrossRef]
- Chen, L.-W.; Soh, S.E.; Tint, M.-T.; Loy, S.L.; Yap, F.; Tan, K.H.; Lee, Y.S.; Shek, L.P.-C.; Godfrey, K.M.; Gluckman, P.D. Combined analysis of gestational diabetes and maternal weight status from pre-pregnancy through post-delivery in future development of type 2 diabetes. Sci. Rep. 2021, 11, 5021. [Google Scholar] [CrossRef]
- WHO Expert Consultation. Appropriate body-mass index for Asian populations and its implications for policy and intervention strategies. Lancet 2004, 363, 157. [Google Scholar] [CrossRef]
- Watkins, O.C.; Selvam, P.; Pillai, R.A.; Cracknell-Hazra, V.; Yong, H.E.J.; Sharma, N.; Cazenave-Gassiot, A.; Bendt, A.K.; Godfrey, K.M.; Lewis, R.M.; et al. Placental 13C-DHA metabolism and relationship with maternal BMI, glycemia and birthweight. Mol. Med. 2021, 27, 84. [Google Scholar] [CrossRef]
- Benassayag, C.; Mignot, T.; Haourigui, M.; Civel, C.; Hassid, J.; Carbonne, B.; Nunez, E.; Ferre, F. High polyunsaturated fatty acid, thromboxane A2, and alpha-fetoprotein concentrations at the human feto-maternal interface. J. Lipid Res. 1997, 38, 276–286. [Google Scholar] [CrossRef]
- Weir, J.M.; Wong, G.; Barlow, C.K.; Greeve, M.A.; Kowalczyk, A.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Jowett, J.B.; Shaw, J. Plasma lipid profiling in a large population-based cohort. J. Lipid Res. 2013, 54, 2898–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, R.A.; Islam, M.O.; Selvam, P.; Sharma, N.; Chu, A.H.; Watkins, O.C.; Godfrey, K.M.; Lewis, R.M.; Chan, S.Y. Placental Inositol Reduced in Gestational Diabetes as Glucose alters Inositol Transporters and IMPA1 enzyme expression. J. Clin. Endocrinol. Metab. 2021, 106, e875–e890. [Google Scholar] [CrossRef]
- Islam, M.O.; Selvam, P.; Pillai, R.A.; Watkins, O.C.; Chan, S.Y. An enzymatic assay for quantification of inositol in human term placental tissue. Anal. Biochem. 2019, 586, 113409. [Google Scholar] [CrossRef]
- Watkins, O.C.; Islam, M.O.; Selvam, P.; Pillai, R.A.; Cazenave-Gassiot, A.; Bendt, A.K.; Karnani, N.; Godfrey, K.M.; Lewis, R.M.; Wenk, M.R.; et al. Metabolism of 13C-Labeled Fatty Acids in Term Human Placental Explants by Liquid Chromatography–Mass Spectrometry. Endocrinology 2019, 160, 1394–1408. [Google Scholar] [CrossRef] [PubMed]
- Watkins, O.C.; Yong, H.E.J.; Mah, T.K.; Cracknell-Hazra, V.K.B.; Pillai, R.A.; Selvam, P.; Sharma, N.; Cazenave-Gassiot, A.; Bendt, A.K.; Godfrey, K.M.; et al. Sex-Dependent Regulation of Placental Oleic Acid and Palmitic Acid Metabolism by Maternal Glycemia and Associations with Birthweight. Int. J. Mol. Sci. 2022, 23, 8685. [Google Scholar] [CrossRef]
- Russo, M.; Forte, G.; Montanino Oliva, M.; Laganà, A.S.; Unfer, V. Melatonin and Myo-Inositol: Supporting Reproduction from the Oocyte to Birth. Int. J. Mol. Sci. 2021, 22, 8433. [Google Scholar] [CrossRef] [PubMed]
- Watkins, O.C.; Islam, M.O.; Selvam, P.; Pillai, R.A.; Cazenave-Gassiot, A.; Bendt, A.K.; Karnani, N.; Godfrey, K.M.; Lewis, R.M.; Wenk, M.R.; et al. Myo-inositol alters 13C-labeled fatty acid metabolism in human placental explants. J. Endocrinol. 2019, 243, 73–84. [Google Scholar] [CrossRef]
- Tint, M.-T.; Sadananthan, S.A.; Soh, S.-E.; Aris, I.M.; Michael, N.; Tan, K.H.; Shek, L.P.; Yap, F.; Gluckman, P.D.; Chong, Y.-S. Maternal glycemia during pregnancy and offspring abdominal adiposity measured by MRI in the neonatal period and preschool years: The Growing Up in Singapore Towards healthy Outcomes (GUSTO) prospective mother–offspring birth cohort study. Am. J. Clin. Nutr. 2020, 112, 39–47. [Google Scholar] [CrossRef]
- Fonteh, A.N.; Chilton, F.H. Mobilization of different arachidonate pools and their roles in the generation of leukotrienes and free arachidonic acid during immunologic activation of mast cells. J. Immunol. 1993, 150, 563–570. [Google Scholar]
- Tessner, T.; Greene, D.G.; Wykle, R. Selective deacylation of arachidonate-containing ethanolamine-linked phosphoglycerides in stimulated human neutrophils. J. Biol. Chem. 1990, 265, 21032–21038. [Google Scholar] [CrossRef]
- Waku, K. Origins and fates of fatty acyl-CoA esters. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1992, 1124, 101–111. [Google Scholar] [CrossRef]
- Johnson, M.M.; Vaughn, B.; Triggiani, M.; Swan, D.D.; Fonteh, A.N.; Chilton, F.H. Role of arachidonyl triglycerides within lipid bodies in eicosanoid formation by human polymorphonuclear cells. Am. J. Respir. Cell Mol. Biol. 1999, 21, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Triggiani, M.; Marone, G. Differential roles for triglyceride and phospholipid pools of arachidonic acid in human lung macrophages. J. Immunol. 1994, 152, 1394–1403. [Google Scholar] [PubMed]
- Balsinde, J.; Winstead, M.V.; Dennis, E.A. Phospholipase A2 regulation of arachidonic acid mobilization. FEBS Lett. 2002, 531, 2–6. [Google Scholar] [CrossRef]
- Triggiani, M.; Giannattasio, G.; Granata, F.; Loffredo, S.; Rossi, F.W.; Salzano, S.; Marone, G. Remodeling of arachidonic acid in inflammatory cells of the human lung. In Arachidonate Remodeling and Inflammation; Springer: Berlin/Heidelberg, Germany, 2004; pp. 115–130. [Google Scholar]
- Kuhn, D.C.; Crawford, M.A.; Stuart, M.J.; Botti, J.J.; Demers, L.M. Alterations in transfer and lipid distribution of arachidonic acid in placentas of diabetic pregnancies. Diabetes 1990, 39, 914–918. [Google Scholar] [CrossRef]
- Araújo, J.R.; Correia-Branco, A.; Ramalho, C.; Keating, E.; Martel, F. Gestational diabetes mellitus decreases placental uptake of long-chain polyunsaturated fatty acids: Involvement of long-chain acyl-CoA synthetase. J. Nutr. Biochem. 2013, 24, 1741–1750. [Google Scholar] [CrossRef]
- Setton-Avruj, C.P.; Speziale, E.H.S.; Sterin-Speziale, N.B. High Glucose Concentrations Stimulate Renal Papillary Phosphatidylcholine Biosynthesis. Nephron Exp. Nephrol. 2001, 9, 301–308. [Google Scholar] [CrossRef]
- Tashiro, S.-I.; Sudou, K.; Imoh, A.Y.A.; Koide, M.; Akazawa, Y. Phosphatidylethanolamine Methyltransferase Activity in Developing, Demyelinating, and Diabetic Mouse Brain. Tohoku J. Exp. Med. 1983, 141, 485–490. [Google Scholar] [CrossRef]
- Hartz, C.S.; Nieman, K.M.; Jacobs, R.L.; Vance, D.E.; Schalinske, K.L. Hepatic phosphatidylethanolamine N-methyltransferase expression is increased in diabetic rats. J. Nutr. 2006, 136, 3005–3009. [Google Scholar] [CrossRef]
- Freed, K.; Moses, E.; Brennecke, S.; Rice, G. Differential expression of type II, IV and cytosolic PLA2 messenger RNA in human intrauterine tissues at term. Mol. Hum. Reprod. 1997, 3, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, W.; Rice, G.; Wong, M.; Scott, K.; Brennecke, S. Release of Type II phospholipase A2 immunoreactivity and phospholipase A2 enzymatic activity from human placenta. J. Endocrinol. 1997, 153, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Rice, G.; Wong, M.; Farrugia, W.; Scott, K. Contribution of Type II phospholipase A2 to in vitro phospholipase A2 enzymatic activity in human term placenta. J. Endocrinol. 1998, 157, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schliefsteiner, C.; Hirschmugl, B.; Kopp, S.; Curcic, S.; Bernhart, E.M.; Marsche, G.; Lang, U.; Desoye, G.; Wadsack, C. Maternal Gestational Diabetes Mellitus increases placental and foetal lipoprotein-associated Phospholipase A2 which might exert protective functions against oxidative stress. Sci. Rep. 2017, 7, 12628. [Google Scholar] [CrossRef]
- Diez, E.; Chilton, F.; Stroup, G.; Mayer, R.; Winkler, J.; Fonteh, A. Fatty acid and phospholipid selectivity of different phospholipase A2 enzymes studied by using a mammalian membrane as substrate. Biochem. J. 1994, 301, 721–726. [Google Scholar] [CrossRef]
- Ishizaki, J.; Suzuki, N.; Higashino, K.; Yokota, Y.; Ono, T.; Kawamoto, K.; Fujii, N.; Arita, H.; Hanasaki, K. Cloning and characterization of novel mouse and human secretory phospholipase A(2)s. J. Biol. Chem. 1999, 274, 24973–24979. [Google Scholar] [CrossRef]
- Suzuki, N.; Ishizaki, J.; Yokota, Y.; Higashino, K.; Ono, T.; Ikeda, M.; Fujii, N.; Kawamoto, K.; Hanasaki, K. Structures, enzymatic properties, and expression of novel human and mouse secretory phospholipase A(2)s. J. Biol. Chem. 2000, 275, 5785–5793. [Google Scholar] [CrossRef]
- Ferchaud-Roucher, V.; Kramer, A.; Silva, E.; Pantham, P.; Weintraub, S.T.; Jansson, T.; Powell, T.L. A potential role for lysophosphatidylcholine in the delivery of long chain polyunsaturated fatty acids to the fetal circulation. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 394–402. [Google Scholar] [CrossRef]
- Hellmuth, C.; Uhl, O.; Standl, M.; Demmelmair, H.; Heinrich, J.; Koletzko, B.; Thiering, E. Cord blood metabolome is highly associated with birth weight, but less predictive for later weight development. Obes. Facts 2017, 10, 85–100. [Google Scholar] [CrossRef]
- Foreman-van Drongelen, M.M.; van Houwelingen, A.C.; Kester, A.D.; Hasaart, T.H.; Blanco, C.E.; Hornstra, G. Long-chain polyunsaturated fatty acids in preterm infants: Status at birth and its influence on postnatal levels. J. Pediatrics 1995, 126, 611–618. [Google Scholar] [CrossRef]
- Ruiz-Palacios, M.; Prieto-Sánchez, M.T.; Ruiz-Alcaraz, A.J.; Blanco-Carnero, J.E.; Sanchez-Campillo, M.; Parrilla, J.J.; Larqué, E. Insulin Treatment May Alter Fatty Acid Carriers in Placentas from Gestational Diabetes Subjects. Int. J. Mol. Sci. 2017, 18, 1203. [Google Scholar] [CrossRef] [PubMed]
- Van Rossum, D.B.; Patterson, R.L. PKC and PLA2: Probing the complexities of the calcium network. Cell Calcium 2009, 45, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Beeson, M.; Sajan, M.P.; Dizon, M.; Grebenev, D.; Gomez-Daspet, J.; Miura, A.; Kanoh, Y.; Powe, J.; Bandyopadhyay, G.; Standaert, M.L. Activation of protein kinase C-ζ by insulin and phosphatidylinositol-3, 4, 5-(PO4) 3 is defective in muscle in type 2 diabetes and impaired glucose tolerance: Amelioration by rosiglitazone and exercise. Diabetes 2003, 52, 1926–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.E.; Saraceno, G.E.; Capani, F.; Castilla, R. Long-chain acyl-CoA synthetase 4 is regulated by phosphorylation. Biochem. Biophys. Res. Commun. 2013, 430, 272–277. [Google Scholar] [CrossRef]
- Radaelli, T.; Lepercq, J.; Varastehpour, A.; Basu, S.; Catalano, P.M.; Hauguel-De Mouzon, S. Differential regulation of genes for fetoplacental lipid pathways in pregnancy with gestational and type 1 diabetes mellitus. Am. J. Obstet. Gynecol. 2009, 201, 209.e1–209.e10. [Google Scholar] [CrossRef]
- Wentzel, P.; Welsh, N.; Eriksson, U.J. Developmental damage, increased lipid peroxidation, diminished cyclooxygenase-2 gene expression, and lowered prostaglandin E2 levels in rat embryos exposed to a diabetic environment. Diabetes 1999, 48, 813–820. [Google Scholar] [CrossRef]
- Hayashi, D.; Mouchlis, V.D.; Dennis, E.A. Unique enzyme specificity of three human phospholipases A2 toward phospholipids containing sn-2 omega-3 and omega-6 fatty acids. FASEB J. 2019, 33, 489–493. [Google Scholar] [CrossRef]
- Colomiere, M.; Permezel, M.; Riley, C.; Desoye, G.; Lappas, M. Defective insulin signaling in placenta from pregnancies complicated by gestational diabetes mellitus. Eur. J. Endocrinol. 2009, 160, 567–578. [Google Scholar] [CrossRef]
- Varma, S.; Lal, B.K.; Zheng, R.; Breslin, J.W.; Saito, S.; Pappas, P.J.; Hobson, R.W.; Durán, W.N. Hyperglycemia alters PI3k and Akt signaling and leads to endothelial cell proliferative dysfunction. Am. J. Physiol.-Heart Circ. Physiol. 2005, 289, H1744–H1751. [Google Scholar] [CrossRef]
- Xu, K.-P.; Li, Y.; Ljubimov, A.V.; Fu-Shin, X.Y. High glucose suppresses epidermal growth factor receptor/phosphatidylinositol 3-kinase/Akt signaling pathway and attenuates corneal epithelial wound healing. Diabetes 2009, 58, 1077–1085. [Google Scholar] [CrossRef]
- Hayashi, E.; Maeda, T.; Tomita, T. The effect of myo-inositol deficiency on lipid metabolism in rats: I. The alteration of lipid metabolism in myo-inositol deficient rats. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1974, 360, 134–145. [Google Scholar] [CrossRef]
- Adam, O. Immediate and long range effects of the uptake of increased amounts of arachidonic acid. Clin. Investig. 1992, 70, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Subbaiah, P.V.; Horvath, P.; Achar, S.B. Regulation of the activity and fatty acid specificity of lecithin-cholesterol acyltransferase by sphingomyelin and its metabolites, ceramide and ceramide phosphate. Biochemistry 2006, 45, 5029–5038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, L.E.; Wells, W.W. Characterization of the lactation-dependent fatty liver in myo-inositol deficient rats. J. Nutr. 1977, 107, 1871–1883. [Google Scholar] [CrossRef]
- Wells, I.C.; Hogan, J. Effects of dietary deficiencies of lipotropic factors on plasma cholesterol esterification and tissue cholesterol in rats. J. Nutr. 1968, 95, 55–62. [Google Scholar] [CrossRef]
- Petit, A.; Guillon, G.; Tence, M.; Jard, S.; Gallo-Payet, N.; Bellabarba, D.; Lehoux, J.G.; Belisle, S. Angiotensin II stimulates both inositol phosphate production and human placental lactogen release from human trophoblastic cells. J. Clin. Endocrinol. Metab. 1989, 69, 280–286. [Google Scholar] [CrossRef]
- Deykin, D.; Jakubowski, J.; Brown, M. Ionophore-induced metabolism of phospholipids and arachidonic acid (AA) in porcine aortic endothelial cells (PAEC): Release of AA from alkenyl-linked phosphatidylethanolamine (PE). Thromb. Haemost. 1987, 58, 1826. [Google Scholar]
- Chilton, F.H. Potential phospholipid source (s) of arachidonate used for the synthesis of leukotrienes by the human neutrophil. Biochem. J. 1989, 258, 327–333. [Google Scholar] [CrossRef]
- Astudillo, A.M.; Pérez-Chacón, G.; Meana, C.; Balgoma, D.; Pol, A.; del Pozo, M.A.; Balboa, M.A.; Balsinde, J. Altered arachidonate distribution in macrophages from caveolin-1 null mice leading to reduced eicosanoid synthesis. J. Biol. Chem. 2011, 286, 35299–35307. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | Total (n = 19) | Non-GDM (n = 10) | GDM (n = 9) |
|---|---|---|---|
| Maternal Age (years) | 32.7 (2.4) | 32.4 (2.3) | 33 (2.6) |
| Chinese:Indian ethnicity | 12 (63%), 7 (37%) | 6 (60%), 4 (40%) | 6 (67%), 3 (33%) |
| Maternal BMI in first trimester (kg/m2) | 25.2 (5.0) | 25.4 (5.5) | 25 (4.8) |
| Normal weight: Overweight: Obese † | 6 (32%), 8 (42%), 5 (26%) | 4 (40%), 3 (30%), 3 (30%) | 4 (44%), 3 (33%), 2 (22%) |
| Fasting glycemia (mmol/L) # | 4.4 (0.2) | 4.4 (0.2) | 4.3 (0.2) |
| 1-h glycemia (mmol/L) # | 9.0 (1.6) | 7.9 (1.3) | 10.3 (0.6) |
| 2-h glycemia (mmol/L) # | 7.3 (1.6) | 6.3 (0.9) | 8.4 (1.6) |
| Gestational age at delivery (days) | 271.0 (5.6) | 272.8 (6.6) | 268.7 (3.4) |
| Female neonates | 9 (47%) | 5 (50%) | 4 (44%) |
| Birthweight (g) | 3150 (356) | 3258 (331) | 3032 (365) |
| Birthweight Centile (%) | 52.4 (33) | 57.3 (33) | 47 (34.8) |
| Treatment | 1 (Control) | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Glucose (mM) | 5 | 5 | 10 | 10 | 17 | 17 |
| Myo-inositol (µM) | 0.3 | 60 | 0.3 | 60 | 0.3 | 60 |
| Number of samples | 19 | 17 ^ | 17 ^ | 18 ^ | 18 ^ | 18 ^ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watkins, O.C.; Cracknell-Hazra, V.K.B.; Pillai, R.A.; Selvam, P.; Yong, H.E.J.; Sharma, N.; Patmanathan, S.N.; Cazenave-Gassiot, A.; Bendt, A.K.; Godfrey, K.M.; et al. Myo-Inositol Moderates Glucose-Induced Effects on Human Placental 13C-Arachidonic Acid Metabolism. Nutrients 2022, 14, 3988. https://doi.org/10.3390/nu14193988
Watkins OC, Cracknell-Hazra VKB, Pillai RA, Selvam P, Yong HEJ, Sharma N, Patmanathan SN, Cazenave-Gassiot A, Bendt AK, Godfrey KM, et al. Myo-Inositol Moderates Glucose-Induced Effects on Human Placental 13C-Arachidonic Acid Metabolism. Nutrients. 2022; 14(19):3988. https://doi.org/10.3390/nu14193988
Chicago/Turabian StyleWatkins, Oliver C., Victoria K. B. Cracknell-Hazra, Reshma Appukuttan Pillai, Preben Selvam, Hannah E. J. Yong, Neha Sharma, Sathya Narayanan Patmanathan, Amaury Cazenave-Gassiot, Anne K. Bendt, Keith M. Godfrey, and et al. 2022. "Myo-Inositol Moderates Glucose-Induced Effects on Human Placental 13C-Arachidonic Acid Metabolism" Nutrients 14, no. 19: 3988. https://doi.org/10.3390/nu14193988