
Recovery of ΔF508-CFTR Function by Citrate

1
Institute of Physiological Chemistry and Pathobiochemistry, Muenster University Hospital, Waldeyerstr. 15, 48129 Muenster, Germany
2
Institute for Anatomy and Experimental Morphology, University Hospital Hamburg-Eppendorf, Martinistrasse 52, 20246 Hamburg, Germany
*
Author to whom correspondence should be addressed.
Nutrients 2022, 14(20), 4283; https://doi.org/10.3390/nu14204283
Submission received: 26 September 2022
/
Revised: 9 October 2022
/
Accepted: 10 October 2022
/
Published: 14 October 2022
(This article belongs to the Special Issue Nutraceuticals and Human Health and Disease)
Abstract
:Treatment of cystic fibrosis relies so far on expensive and sophisticated drugs. A logical approach to rescuing the defective ΔF508-CFTR protein has not yet been published. Therefore, virtual docking of ATP and CFTR activators to the open conformation of the CFTR protein was performed. A new ATP binding site outside of the two known locations was identified. It was located in the cleft between the nucleotide binding domains NBD1 and NBD2 and comprised six basic amino acids in close proximity. Citrate and isocitrate were also bound to this site. Citrate was evaluated for its action on epithelial cells with intact CFTR and defective ΔF508-CFTR. It activated hyaluronan export from human breast carcinoma cells and iodide efflux, and recovered ΔF508-CFTR from premature intracellular degradation. In conclusion, citrate is an activator for ΔF508-CFTR and increases export by defective ΔF508-CFTR into the extracellular matrix of epithelial cells.
1. Introduction
Cystic fibrosis (CF) is one of the most common inherited diseases, afflicting 1 in approximately 2500 white individuals [1]. The primary cause of morbidity and mortality in CF is chronic lung infection and deterioration of lung function. CF is caused by mutations in the CF transmembrane conductance regulator (CFTR) gene, which encodes a chloride channel expressed at the apical membrane of epithelial cells in the airways, pancreas, testis, and other tissues [2,3]. The most common CFTR mutation producing CF is deletion of phenylalanine at residue 508 (ΔF508) in its amino acid sequence. The ΔF508-CFTR protein is misfolded and retained at the endoplasmic reticulum, where it is degraded rapidly. Small-molecule activators of defective ΔF508-CFTR folding/cellular processing (“correctors”) and channel gating (“potentiators”) may provide a strategy for therapy of CF that corrects the underlying defect. A number of small-molecule ΔF508-CFTR potentiators and correctors have been identified [4]. They were found by high-throughput screening for activation of the chloride channel.
We discovered that hyaluronan is exported from mesenchymal fibroblasts by MRP5 (multidrug resistance associated protein 5) [5] and from epithelial cells by CFTR [6]. Hyaluronan has an important role in epithelial clearing of the bronchial surface by facilitating cilial beating [7,8]. In an attempt to evaluate interaction between CFTR and hyaluronan transmembrane transport, we synthesized a new class of drug-like compounds (Hylout4) that mimic the non-reducing end of hyaluronan and discovered that they activated chloride export from bronchial epithelial cells and hyaluronan export from breast cancer cells [9]. The compounds together with other known modulators were docked to the open and closed conformation of ΔF508-CFTR [10] to identify possible optimized activators for ΔF508-CFTR. Molecular modelling and physiological experiments showed that citrate serves as an activator of hyaluronan transport in ΔF508-CFTR.
2. Materials and Methods
2.1. Materials
Mouse-anti-CFTR-IgM was from Acris Antibodies, Hiddenhausen, Germany. Other chemicals were from Sigma-Aldrich Chemical Corporation (St. Louis, MO, USA).
2.2. Computational Studies
In silico docking studies were carried out to evaluate the affinity and binding interactions by molecular docking simulation using the AutoDock 4.2 and PyRx software—Python Prescription 0.8 to coordinates of the open and closed CFTR conformation [10,11,12]. ATP, citrate and docosahexaenoic acid (DHA) were docked as fully ionized molecules to enable ionic interaction with basic amino acids. The theoretical KI-values of binding are listed in Table 1.
2.3. Cell Lines
2.4. Western Blotting of CFTR
Western blotting was performed as described [9].
2.5. Determination of Hyaluronan Export
Hyaluronan export of HMT3552 cells was performed as described [20].
2.6. Iodide Efflux
Iodide efflux experiments were performed as described [21].
3. Results
3.1. Virtual Docking
In an attempt to identify optimized CFTR activators, we docked Hylout4 [9] and several other known activators and inhibitors to the open and closed conformation of CFTR [10,12]. In preliminary docking calculations, we found that the outward-facing CFTR configuration [10,11] bound Hylout4 with higher affinities than the inward-facing ATP free configurations. The predicted Hylout4 binding site was a cleft between the nucleotide binding domains NBD1 (433 to 634) and NBD2 (1225 to 1415). The CFTR activators corr-4a, CFTRact-06, VX809, the inhibitor CFTRinh-172 and dietary supplement docosahexaenoic acid (DHA) also bound to this cavity with the highest affinity (Table 1). It comprised the amino acids R170, A171, E267, W465, F494, R553, K968, K1060, W1063, K1292, G1342, and K1351. The most striking feature of this site was the close neighbourhood of six basic amino acids (underlined) and the preference of lysines that have long flexible arms for ligand binding by ionic interactions and hydrogen bonds. The distances of positive charges in the open conformation ranged from 5 Å to 16 Å and in the closed conformation from 12 Å to 32 Å.
3.2. Activation of Hyaluronan Export
Since hyaluronan is exported from human breast carcinoma cells by CFTR [5], citrate and isocitrate were tested for their influence on hyaluronan export in cell culture. Figure 2 shows that both compounds activated hyaluronan export in a concentration-dependent manner. Citrate showed a higher increase of hyaluronan export than isocitrate. Therefore, citrate was chosen for further evaluation.
3.3. Iodide Efflux
In addition to chloride and hyaluronan, CFTR also exports iodide that can be measured by an iodide selective electrode. Iodide efflux from 16HBE14o- and CFBE41o- cells was assessed with increasing citrate concentrations. Citrate activated immediately iodide efflux from 16HBE14o- as well as from CFBE41o- cells (Figure 3), indicating that citrate recovered the activity of ΔF508-CFTR.
3.4. Recovery of ΔF508-CFTR Cell Surface Expression
The ΔF508-CFTR mutation impairs maturation and destabilizes the protein in post-Golgi compartments. Figure 4 shows that citrate increased ΔF508-CFTR cell surface expression, indicating that citrate recovered cellular processing of ΔF508-CFTR.
4. Discussion
It is surprising that virtual docking of ATP has so far been conducted on computed CFTR fragments [22], but not the whole structural model, in spite of the long history of intensive pharmaceutical research. It is even more surprising that the highest docking affinity for ATP and other activators of CFTR function did not reside in the two known ATP binding sites, but in a cleft between the nucleotide binding sites NBD1 and NBD2. This site has been claimed to open the CFTR channel due to salt bridges between opposing basic and acidic residues [23]. Remarkably, the external phosphate of ATP overlaps with the positions of citrate and the carboxyl group of DHA. Thus, our docking results confirm the notion of salt bridges between the NBD1 and NBD2 domains; however, we propose that the salt bridges are mediated by anionic activators.
These basic amino acids are probably responsible for ionic nucleotide binding and stabilization of the open CFTR conformation. The ATP binding affinity to this binding site is very high with KD = 22 nM in comparison to normal intracellular nucleotide triphosphate concentrations in the mM range, but such high ATP concentrations are required for reliable gating [24,25]. The ATP binding sites had previously been located by site-directed CFTR mutations to W401 in NBD1 and Y1219 in NBD2 [26] which are only 9.8 Å and 11.4 Å apart from our predicted amino acids R170 and K968, respectively. Virtual docking also revealed that the most known CFTR activators bound to this cavity by bridging both nucleotide binding domains and thus appear to function as ATP agonists stabilizing the open conformation. This mechanism of action has been shown for the CFTR activator VX-770, which opens the ΔF508-CFTR in an ATP-independent manner [27] and decouples gating from ATP hydrolysis [28]. It is interesting to note that also the inhibitor CFTRinh-172 had high affinity to the ATP binding site. It could be an ATP antagonist which displaces ATP, destabilizing the open conformation.
The conspicuous proximity of six basic amino acids led us to investigate other possible ligands. We tested citrate and isocitrate and found considerable affinity of KD = 2.7 µM and KD = 1.1 µM, respectively. These theoretical affinities were again much higher than the normal cytosolic concentrations of 80 µM to 220 µM for citrate and about 10 µM for isocitrate [29,30], indicating that high saturation density is again necessary for gating. The levels of both ATP and citrate are indicators of the cellular energy level [31].
Accordingly, our cell culture experiments showed that citrate activated hyaluronan export from breast carcinoma cells, stimulated iodide efflux from epithelial cells carrying normal CFTR as well as ΔF508-CFTR and recovered it from premature intracellular degradation. It can thus be regarded as a potentiator as well as a corrector for the defective ΔF508-CFTR protein.
Part of the cytosolic citrate is exported into the circulation, giving rise to citrate in plasma and urine. It is known that urinary citrate secretion from cystic fibrosis patients was significantly lower [32,33]. A striking similarity of citrate and hyaluronan is their high Ca2+-chelating property with KD = 171 µM [34] and KD = 1.4 mM [35], respectively, as compared to the normal plasma concentration of about 2 mM. Cystic fibrosis patients have abnormal Ca2+ homeostasis [36], and chelate formation of cytosolic citrate and Ca2+ could reduce the effective Ca2+ concentration which is required for normal ΔF508-CFTR trafficking [37].
The function of citrate as Ca2+ chelator and as hyaluronan export activator may explain some of the observed metabolic disturbances of cystic fibrosis patients such as the associated diabetes [38] and lithiasis [33]. Diabetes favours a shift from the aerobic energy production by the citric acid cycle and respiration towards gluconeogenesis, causing withdrawal of metabolites from the citric acid cycle. Low citrate in other tissues such as kidney again decreases extracellular hyaluronan required for Ca2+ complexation and solubilization, causing lithiasis in cystic fibrosis patients [39,40,41,42]. Patients thus remain in a vicious cycle, unless they are treated by citrate as a food additive for stone solubilization [43,44,45,46,47,48,49,50]. Citrate is freely available in the supermarket and also a nutritional additive for treatment of lithiasis.
Citrate has been patented as an ingredient of inhalation solutions to reduce the viscosity of the sticky mucous in cystic fibrosis patients [51]. It was speculated that citrate might correct all other symptoms of cystic fibrosis by systemically recovering the function of ΔF508-CFTR.
Citrate and isocitrate are intermediates of the citric acid cycle within mitochondria. They are nutrients taken up by mammalian cells and form two separated citrate pools: mitochondrial and cytosolic. The intracellular citrate level depends on nutrition: A carbohydrate-rich nutrition induces the citrate lyase to reduce the citrate concentration by transformation into fatty acids. A protein-rich nutrition induces anaplerotic reactions to increase intracellular citrate concentrations. A protein-rich diet has frequently been evaluated over the past decades with no final conclusion, but it seemed to be underestimated [52] probably because activation of hyaluronan export by citrate by the regulatory balance of carbohydrate-rich and protein-rich diets was not taken into account. Indeed, a low glycemic index diet has already been shown to be beneficial for cystic fibrosis patients [53,54].
Frequent and severe infections of cystic fibrosis patients by Pseudomonas aeruginosa are often responsible for morbidity in CF patients. These bacteria can upregulate an isocitrate lyase as a virulence factor for persistent infection [55], which further depletes citrate levels and thus inactivates hyaluronan export. Sufficient hyaluronan shedding is required for efficient ciliary beating and debris removal from airways [8]. Thus, a protein-rich and low carbohydrate diet supplemented with DHA and citrate could have benefits similar to expensive drugs and lucrative treatments.
Author Contributions
B.B. performed experiments. P.P. conceived the project and performed experiments. All authors have read and agreed to the published version of the manuscript.
Funding
Deutsche Forschungsgemeinschaft (SFB 492).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
We have no conflict of interests.
References
- Bobadilla, J.L.; Macek, M., Jr.; Fine, J.P.; Farrell, P.M. Cystic fibrosis: A worldwide analysis of CFTR mutations--correlation with incidence data and application to screening. Hum. Mutat. 2002, 19, 575–606. [Google Scholar] [CrossRef] [PubMed]
- Pilewski, J.M.; Frizzell, R.A. Role of CFTR in airway disease. Physiol. Rev. 1999, 79, S215–S255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratt, S.; Shepard, R.L.; Kandasamy, R.A.; Johnston, P.A.; Perry, W., III; Dantzig, A.H. The multidrug resistance protein 5 (ABCC5) confers resistance to 5-fluorouracil and transports its monophosphorylated metabolites. Mol. Cancer Ther. 2005, 4, 855–863. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Kota, P.; Aleksandrov, A.A.; Cui, L.; Jensen, T.; Dokholyan, N.V.; Riordan, J.R. Correctors of {Delta}F508 CFTR restore global conformational maturation without thermally stabilizing the mutant protein. FASEB J. 2012, 27, 536–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, T.; Schumacher, U.; Prehm, P. Hyaluronan export by the ABC-transporter MRP5 and its modulation by intracellular cGMP. J. Biol. Chem. 2007, 282, 20999–21004. [Google Scholar] [CrossRef] [Green Version]
- Schulz, T.; Schumacher, U.; Prante, C.; Sextro, W.; Prehm, P. Cystic Fibrosis Transmembrane Conductance Regulator Can Export Hyaluronan. Pathobiology 2010, 77, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Lieb, T.; Forteza, R.; Salathe, M. Hyaluronic acid in cultured ovine tracheal cells and its effect on ciliary beat frequency in vitro. J. Aerosol. Med. 2000, 13, 231–237. [Google Scholar] [CrossRef]
- Forteza, R.; Lieb, T.; Aoki, T.; Savani, R.C.; Conner, G.E.; Salathe, M. Hyaluronan serves a novel role in airway mucosal host defense. FASEB J. 2001, 15, 2179–2186. [Google Scholar] [CrossRef] [PubMed]
- Nowakowska, E.; Schulz, T.; Molenda, N.; Schillers, H.; Prehm, P. Recovery of ΔF508-CFTR function by analogs of hyaluronan disaccharide. J. Cell Biochem. 2012, 113, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Mornon, J.P.; Lehn, P.; Callebaut, I. Atomic model of human cystic fibrosis transmembrane conductance regulator: Membrane-spanning domains and coupling interfaces. Cell Mol. Life Sci. 2008, 65, 2594–2612. [Google Scholar] [CrossRef] [PubMed]
- Serohijos, A.W.; Hegedus, T.; Aleksandrov, A.A.; He, L.; Cui, L.; Dokholyan, N.V.; Riordan, J.R. Phenylalanine-508 mediates a cytoplasmic-membrane domain contact in the CFTR 3D structure crucial to assembly and channel function. Proc. Natl. Acad. Sci. USA 2008, 105, 3256–3261. [Google Scholar] [CrossRef] [Green Version]
- Mornon, J.P.; Lehn, P.; Callebaut, I. Molecular models of the open and closed states of the whole human CFTR protein. Cell Mol. Life Sci. 2009, 66, 3469–3486. [Google Scholar] [CrossRef]
- Szellas, T.; Nagel, G. Apparent affinity of CFTR for ATP is increased by continuous kinase activity. FEBS Lett. 2003, 535, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Mills, A.D.; Yoo, C.; Butler, J.D.; Yang, B.; Verkman, A.S.; Kurth, M.J. Design and synthesis of a hybrid potentiator-corrector agonist of the cystic fibrosis mutant protein DeltaF508-CFTR. Bioorg. Med. Chem. Lett. 2010, 20, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Van, G.F.; Hadida, S.; Grootenhuis, P.D.; Burton, B.; Stack, J.H.; Straley, K.S.; Decker, C.J.; Miller, M.; McCartney, J.; Olson, E.R.; et al. Correction of the F508del-CFTR protein processing defect in vitro by the investigational drug VX-809. Proc. Natl. Acad. Sci. USA 2011, 108, 18843–18848. [Google Scholar]
- Ma, T.; Thiagarajah, J.R.; Yang, H.; Sonawane, N.D.; Folli, C.; Galietta, L.J.; Verkman, A.S. Thiazolidinone CFTR inhibitor identified by high-throughput screening blocks cholera toxin-induced intestinal fluid secretion. J. Clin. Investig. 2002, 110, 1651–1658. [Google Scholar] [CrossRef]
- López-Neyra, A.; Suárez, L.; Munoz, M.; de Blas, A.; Ruiz de Valbuena, M.; Garriga, M.; Calvo, J.; Ribes, C.; Giron, M.R.; Máiz, L.; et al. Long-term docosahexaenoic acid (DHA) supplementation in cystic fibrosis patients: A randomized, multi-center, double-blind, placebo-controlled trial. Prostaglandins Leukot. Essent. Fatty Acids 2020, 162, 102186. [Google Scholar] [CrossRef]
- Jojovic, M.; Delpech, B.; Prehm, P.; Schumacher, U. Expression of hyaluronate and hyaluronate synthase in human primary tumours and their metastases in scid mice. Cancer Lett. 2002, 188, 181–189. [Google Scholar] [CrossRef]
- Grskovic, B.; Pollaschek, C.; Mueller, M.M.; Stuhlmeier, K.M. Expression of hyaluronan synthase genes in umbilical cord blood stem/progenitor cells. Biochim. Biophys. Acta 2006, 1760, 890–895. [Google Scholar] [CrossRef]
- Monz, K.; Maas-Kuck, K.; Schumacher, U.; Schulz, T.; Hallmann, R.; Schnaker, E.M.; Schneider, S.W.; Prehm, P. Inhibition of hyaluronan export attenuates cell migration and metastasis of human melanoma. J. Cell Biochem. 2008, 105, 1260–1266. [Google Scholar] [CrossRef]
- Lansdell, K.A.; Kidd, J.F.; Delaney, S.J.; Wainwright, B.J.; Sheppard, D.N. Regulation of murine cystic fibrosis transmembrane conductance regulator Cl- channels expressed in Chinese hamster ovary cells. J. Physiol. 1998, 512, 751–764. [Google Scholar] [CrossRef]
- Huang, S.Y.; Bolser, D.; Liu, H.Y.; Hwang, T.C.; Zou, X. Molecular modeling of the heterodimer of human CFTR’s nucleotide-binding domains using a protein-protein docking approach. J. Mol. Graph. Model. 2009, 27, 822–828. [Google Scholar] [CrossRef] [Green Version]
- Strickland, K.M.; Stock, G.; Cui, G.; Hwang, H.; Infield, D.T.; Schmidt-Krey, I.; McCarty, N.A.; Gumbart, J.C. ATP-Dependent Signaling in Simulations of a Revised Model of Cystic Fibrosis Transmembrane Conductance Regulator (CFTR). J. Phys. Chem. B 2019, 123, 3177–3188. [Google Scholar] [CrossRef]
- Shimizu, H.; Yu, Y.C.; Kono, K.; Kubota, T.; Yasui, M.; Li, M.; Hwang, T.C.; Sohma, Y. A stable ATP binding to the nucleotide binding domain is important for reliable gating cycle in an ABC transporter CFTR. J. Physiol. Sci. 2010, 60, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.F.; Li, M.; Hwang, T.C. Stable ATP binding mediated by a partial NBD dimer of the CFTR chloride channel. J. Gen. Physiol. 2010, 135, 399–414. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Wang, X.; Liu, H.Y.; Zou, X.; Li, M.; Hwang, T.C. The two ATP binding sites of cystic fibrosis transmembrane conductance regulator (CFTR) play distinct roles in gating kinetics and energetics. J. Gen. Physiol. 2006, 128, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Eckford, P.D.; Li, C.; Ramjeesingh, M.; Bear, C.E. Cystic fibrosis transmembrane conductance regulator (CFTR) potentiator VX-770 (ivacaftor) opens the defective channel gate of mutant CFTR in a phosphorylation-dependent but ATP-independent manner. J. Biol. Chem. 2012, 287, 36639–36649. [Google Scholar] [CrossRef] [Green Version]
- Jih, K.Y.; Hwang, T.C. Vx-770 potentiates CFTR function by promoting decoupling between the gating cycle and ATP hydrolysis cycle. Proc. Natl. Acad. Sci. USA 2013, 110, 4404–4409. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Akerboom, T.P.; Tager, J.M. Mitochondrial and cytosolic NADPH systems and isocitrate dehydrogenase indicator metabolites during ureogensis from ammonia in isolated rat hepatocytes. Eur. J. Biochem. 1977, 72, 301–307. [Google Scholar] [CrossRef]
- Kauppinen, R.A.; Hiltunen, J.K.; Hassinen, I.E. Compartmentation of citrate in relation to the regulation of glycolysis and the mitochondrial transmembrane proton electrochemical potential gradient in isolated perfused rat heart. Biochim. Biophys. Acta 1982, 681, 286–291. [Google Scholar] [CrossRef]
- Lin, C.C.; Cheng, T.L.; Tsai, W.H.; Tsai, H.J.; Hu, K.H.; Chang, H.C.; Yeh, C.W.; Chen, Y.C.; Liao, C.C.; Chang, W.T. Loss of the respiratory enzyme citrate synthase directly links the Warburg effect to tumor malignancy. Sci. Rep. 2012, 2, 785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campuzano, S.; Diaz, J.J.; Bousono, C.; Rodriguez, M.; Campos, C.; Malaga, S. Risk of urolithiasis in patients with cystic fibrosis. Nefrologia 2009, 29, 61–66. [Google Scholar]
- Gibney, E.M.; Goldfarb, D.S. The association of nephrolithiasis with cystic fibrosis. Am. J. Kidney Dis. 2003, 42, 1–11. [Google Scholar] [CrossRef]
- Wiley, W.J. The dissociation of calcium citrate. Biochem. J. 1930, 24, 856–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, G.K.; Wong, K.S.; Kim, J.J. Binding of calcium to glycosaminoglycans: An equilibrium dialysis study. Arch. Biochem. Biophys. 1988, 260, 161–167. [Google Scholar] [CrossRef]
- Antigny, F.; Norez, C.; Cantereau, A.; Becq, F.; Vandebrouck, C. Abnormal spatial diffusion of Ca2+ in F508del-CFTR airway epithelial cells. Respir. Res. 2008, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Norez, C.; Antigny, F.; Becq, F.; Vandebrouck, C. Maintaining low Ca2+ level in the endoplasmic reticulum restores abnormal endogenous F508del-CFTR trafficking in airway epithelial cells. Traffic 2006, 7, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C. Cystic fibrosis-related diabetes. Nat. Rev. Endocrinol. 2011, 7, 375. [Google Scholar] [CrossRef]
- Terribile, M.; Capuano, M.; Cangiano, G.; Carnovale, V.; Ferrara, P.; Petrarulo, M.; Marangella, M. Factors increasing the risk for stone formation in adult patients with cystic fibrosis. Nephrol. Dial. Transpl. 2006, 21, 1870–1875. [Google Scholar] [CrossRef]
- von der Heiden, R.; Balestra, A.P.; Bianchetti, M.G.; Casaulta, A.C.; Mullis, P.E.; Lippuner, K.; Jaeger, P. Which factors account for renal stone formation in cystic fibrosis? Clin. Nephrol. 2003, 59, 160–163. [Google Scholar] [CrossRef]
- Perez-Brayfield, M.R.; Caplan, D.; Gatti, J.M.; Smith, E.A.; Kirsch, A.J. Metabolic risk factors for stone formation in patients with cystic fibrosis. J. Urol. 2002, 167, 480–484. [Google Scholar] [CrossRef]
- Scott, W.J.; Block, G.E. Biliary stone disease in adults with cystic fibrosis. Surgery 1989, 105, 671–673. [Google Scholar]
- Spivacow, F.R.; Negri, A.L.; Polonsky, A.; Del Valle, E.E. Long-term treatment of renal lithiasis with potassium citrate. Urology 2010, 76, 1346–1349. [Google Scholar] [CrossRef] [PubMed]
- Conte, A.; Piza, P.; Garcia-Raja, A.; Grases, F.; Costa-Bauza, A.; Prieto, R.M. Urinary lithogen risk test: Usefulness in the evaluation of renal lithiasis treatment using crystallization inhibitors (citrate and phytate). Arch. Esp. Urol. 1999, 52, 305–310. [Google Scholar]
- Miyake, O.; Yoshimura, K.; Yoshioka, T.; Koide, T.; Okuyama, A. High urinary excretion level of citrate and magnesium in children: Potential etiology for the reduced incidence of pediatric urolithiasis. Urol. Res. 1998, 26, 209–213. [Google Scholar] [CrossRef]
- Guitron, A.; Gonzalez-Loya, H.; Barinagarrementeria, R.; Sarol, J.C.; Adalid, R.; Rodriguez-Delgado, J. Dissolution of pancreatic lithiasis by direct citrate application into the pancreatic duct in two patients with chronic idiopathic pancreatitis. Dig. Dis. 1997, 15, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Barcelo, P.; Wuhl, O.; Servitge, E.; Rousaud, A.; Pak, C.Y. Randomized double-blind study of potassium citrate in idiopathic hypocitraturic calcium nephrolithiasis. J. Urol. 1993, 150, 1761–1764. [Google Scholar] [CrossRef]
- Pak, C.Y.; Fuller, C.; Sakhaee, K.; Preminger, G.M.; Britton, F. Long-term treatment of calcium nephrolithiasis with potassium citrate. J. Urol. 1985, 134, 11–19. [Google Scholar] [CrossRef]
- Pak, C.Y.; Sakhaee, K.; Fuller, C.J. Physiological and physiochemical correction and prevention of calcium stone formation by potassium citrate therapy. Trans. Assoc. Am. Physicians 1983, 96, 294–305. [Google Scholar] [PubMed]
- Schwille, P.O.; Scholz, D.; Paulus, M.; Engelhardt, W.; Sigel, A. Citrate in daily and fasting urine: Results of controls, patients with recurrent idiopathic calcium urolithiasis, and primary hyperparathyroidism. Investig. Urol. 1979, 16, 457–462. [Google Scholar]
- Elliott, R.B. Methods and Compositions for the Treatment of Lung Disorders. WO2011142677. 2011. Available online: https://patentscope2.wipo.int/search/en/detail.jsf?docId=WO2011142677 (accessed on 9 October 2022).
- Engelen, M.P.; Com, G.; Deutz, N.E. Protein is an important but undervalued macronutrient in the nutritional care of patients with cystic fibrosis. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 515–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorji, Z.; Modaresi, M.; Yekanni-Nejad, S.; Mahmoudi, M. Effects of low glycemic index/high-fat, high-calorie diet on glycemic control and lipid profiles of children and adolescence with cystic fibrosis: A randomized double-blind controlled clinical trial. Diabetes Metab. Syndr. 2020, 14, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Gorji, Z.; Modaresi, M.; Yekanni-Nejad, S.; Rezaei, N.; Mahmoudi, M. Comparing effects of low glycemic index/high-fat, high-calorie diet and high-fat, high-calorie diet on cytokine levels of patients with cystic fibrosis: A randomized controlled clinical trial. Eur. Cytokine Netw. 2020, 31, 32–38. [Google Scholar] [PubMed]
- Hagins, J.M.; Locy, R.; Silo-Suh, L. Isocitrate lyase supplies precursors for hydrogen cyanide production in a cystic fibrosis isolate of Pseudomonas aeruginosa. J. Bacteriol. 2009, 191, 6335–6339. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Binding site of citrate and ATP to the open conformation of CFTR. (a) Citrate (green) binds to a cleft between the nucleotide binding domains NBD1 (red) and NBD2 (blue). (b) Citrate (green) and ATP (yellow) bind to basic amino acid residues in the vicinity of F508.
Figure 1.
Binding site of citrate and ATP to the open conformation of CFTR. (a) Citrate (green) binds to a cleft between the nucleotide binding domains NBD1 (red) and NBD2 (blue). (b) Citrate (green) and ATP (yellow) bind to basic amino acid residues in the vicinity of F508.

Figure 2.
Activation of hyaluronan export by citrate (■) and isocitrate (▲) from HMT3552 cells. The error bars indicate the means of two determinations.
Figure 2.
Activation of hyaluronan export by citrate (■) and isocitrate (▲) from HMT3552 cells. The error bars indicate the means of two determinations.
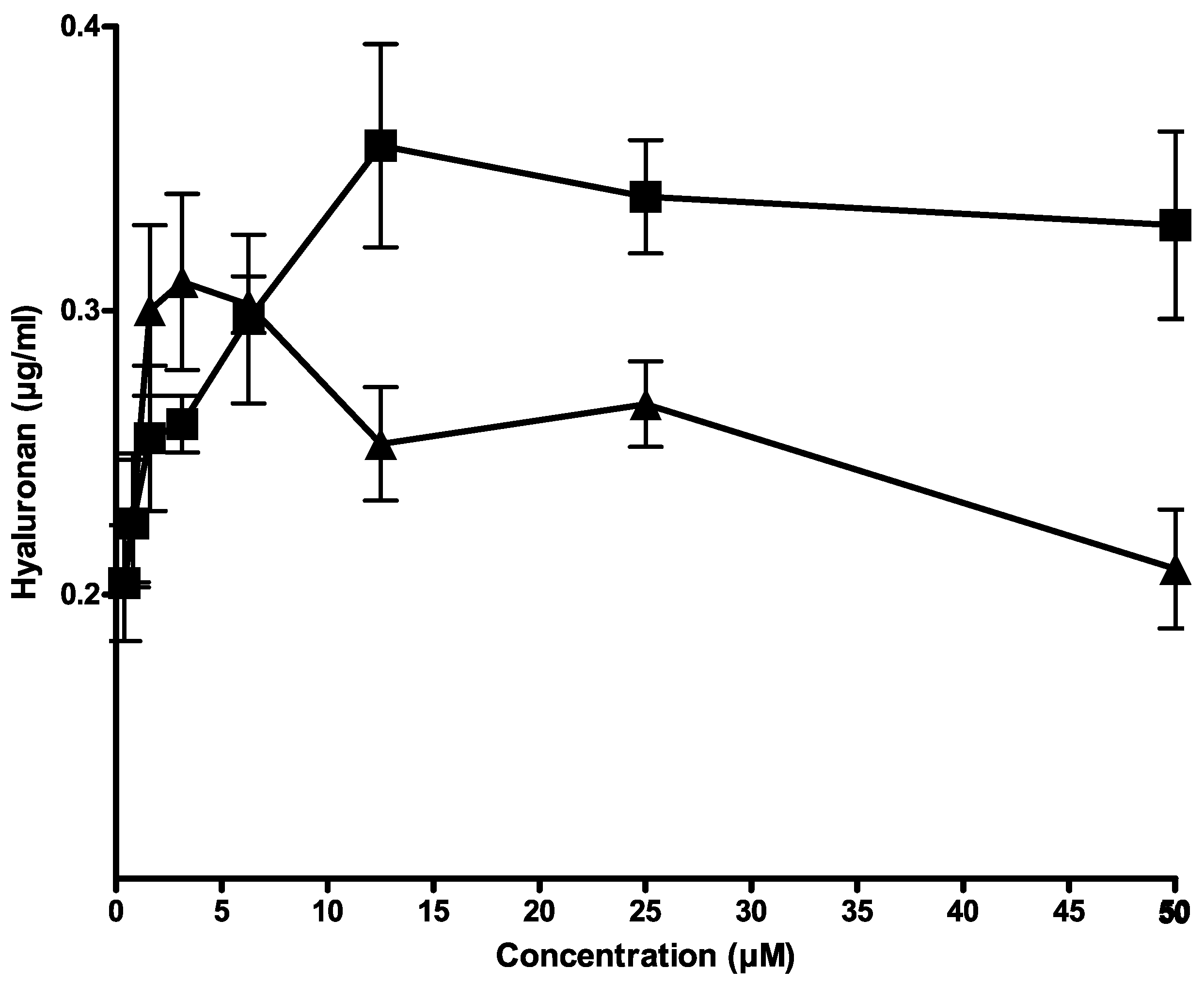
Figure 3.
Citrate stimulated iodide efflux. CFTR can also export iodide instead of chloride. 16HBE14o- or CFBE41o- were loaded with iodide and the iodide concentration was measured by an iodide-sensitive electrode in 1 min intervals. The bar indicates the time period of exposure to 100 µM of citrate.
Figure 3.
Citrate stimulated iodide efflux. CFTR can also export iodide instead of chloride. 16HBE14o- or CFBE41o- were loaded with iodide and the iodide concentration was measured by an iodide-sensitive electrode in 1 min intervals. The bar indicates the time period of exposure to 100 µM of citrate.

Figure 4.
(a) Citrate recovered from ΔF508-CFTR cell surface expression as analysed by Western blotting. (b) relative abundance of cell surface-expressed ΔF508-CFTR.
Figure 4.
(a) Citrate recovered from ΔF508-CFTR cell surface expression as analysed by Western blotting. (b) relative abundance of cell surface-expressed ΔF508-CFTR.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Virtual docking of activators and inhibitors to the open conformation of CFTR.
| Name | Structure | Calculated Affinity Km (µM) | Experimental Activation/ Inhibition EC50 (µM) | Ref. |
|---|---|---|---|---|
| Hylout4 |  | 7 | 100 for HA | [9] |
| ATP |  | 0.022 | 41 for Cl− | [13] |
| CFTRact-06 |  | 1.4 | 1 for HA | [6] |
| Corr-4a |  | 0.01 | 1 for Cl− | [14] |
| VX809 |  | 0.01 | 0.081 for Cl− | [15] |
| CFTRinh127 |  | 0.22 | 0.3 for Cl− | [16] |
| DHA |  | 17 | [17] | |
| Citrate |  | 2.7 | 6 for HA | |
| Isocitrate |  | 1.1 |
Docking was performed with the AutoDock software 1.5.6.rc3. The affinities of the best fits are listed.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Borkenhagen, B.; Prehm, P. Recovery of ΔF508-CFTR Function by Citrate. Nutrients 2022, 14, 4283. https://doi.org/10.3390/nu14204283
AMA Style
Borkenhagen B, Prehm P. Recovery of ΔF508-CFTR Function by Citrate. Nutrients. 2022; 14(20):4283. https://doi.org/10.3390/nu14204283
Chicago/Turabian StyleBorkenhagen, Beatrice, and Peter Prehm. 2022. "Recovery of ΔF508-CFTR Function by Citrate" Nutrients 14, no. 20: 4283. https://doi.org/10.3390/nu14204283
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.