Abstract
Fatty acids play important roles in maintaining ovarian steroidogenesis and endometrial receptivity. Porcine primary ovarian granulosa cells (PGCs) and endometrial epithelial cells (PEECs) were treated with or without medium- and short-chain fatty acids (MSFAs) for 24 h. The mRNA abundance of genes was detected by fluorescence quantitative PCR. The hormone levels in the PGCs supernatant and the rate of adhesion of porcine trophoblast cells (pTrs) to PEECs were measured. Sows were fed diets with or without MSFAs supplementation during early gestation. The fecal and vaginal microbiomes were identified using 16S sequencing. Reproductive performance was recorded at parturition. MSFAs increased the mRNA abundance of genes involved in steroidogenesis, luteinization in PGCs and endometrial receptivity in PEECs (p < 0.05). The estrogen level in the PGC supernatant and the rate of adhesion increased (p < 0.05). Dietary supplementation with MSFAs increased serum estrogen levels and the total number of live piglets per litter (p < 0.01). Moreover, MSFAs reduced the fecal Trueperella abundance and vaginal Escherichia-Shigella and Clostridium_sensu_stricto_1 abundance. These data revealed that MSFAs improved pregnancy outcomes in sows by enhancing ovarian steroidogenesis and endometrial receptivity while limiting the abundance of several intestinal and vaginal pathogens at early stages of pregnancy.
1. Introduction
In mammals, 30–50% of pregnancy loss occurs during early pregnancy, profoundly influenced by embryo implantation [1]. Successful embryo implantation requires proper embryo development and a receptive endometrium. Maintaining ovarian function has great significance for both embryo development and endometrial receptivity. Ovarian granulosa cells synthesize various hormones and growth factors to regulate the growth, differentiation and maturation of follicular membrane cells and oocytes, as well as follicular development [2]. The reproductive hormones produced by ovarian granulosa cells also play important roles in ensuring the development of embryos and endometrial receptivity [3,4,5,6]. Meanwhile, the endometrium goes through a series of cyclic changes and remains receptive for a short time during early pregnancy [7].
Lipid metabolism plays significant roles in embryo development and implantation by converting lipids to lysophosphatidic acid, cannabinoids, prostaglandins, sphingosine-1-phosphate and steroid hormones [8,9,10,11,12]. As important metabolites of lipids, fatty acids play critical roles in embryonic development and implantation [13,14,15]. In vitro experiments showed that adding 1 μM docosahexaenoic acid (DHA) to the medium improved the quality of bovine oocytes, embryo development capacity and blastocyst cell number, while arachidonic acid treatment increased plasma phospholipase A-2α (CPA-2 α) phosphorylation in mouse endometrial stromal cells. Additionally, arachidonic acid could induce prostaglandin-endoperoxide synthase 2 (PTGS2) expression in endometrial interstitial cells and then promote embryo implantation and decidualization [16]. Similarly, in vivo studies found that conjugated linoleic acid rich diet reduced the expression of cannabinoid receptor 2 (CNR2) and increased the progesterone content in cattle [17]. However, long-chain saturated fatty acids often have a negative effect on reproductive performance. Nonesterified fatty acids reduce the bovine blastocyst development rate and total cell number in vitro [18]. In vitro cultures of mouse embryos in palmitic acid increase the mRNA abundance of insulin-like growth factor 1 (IGF-1), and glutamate pyruvate transaminase (GPT2) reduces the total cell number of blastocysts and increases the apoptotic rate of trophoblast cells [19]. In contrast, in addition to providing energy for cells, short- and medium-chain fatty acids exhibit various regulatory and signaling functions [20,21,22]. However, there are only a few reports on the effects of medium- and short-chain fatty acids on reproductive performance. In a study of rats, diets were supplemented with either high fiber or butyric acid (SB), and both increased fetal number and reduced miscarriage rates. Among them, a high-fiber diet enhanced the antioxidant capacity in maternal rats and fetuses, while sodium butyrate had no effect on antioxidant capacity, demonstrating that the mechanism by which sodium butyrate promotes reproductive performance may be different from that of a high-fiber diet, but the specific mechanism is still unknown [23]. By measuring the metabolites (mainly amino acids and fatty acids) and labeling caprylic acid in the culture medium of early embryos of mice at different stages, it was found that caprylic acid could provide energy for the early mouse embryos. In particular, in the absence of energy substances in the culture medium, the early mouse embryos consumed more caprylic acid [24]. As mentioned above, research has shown that various fatty acids have impacts on the development of early pregnancy, but studies on the reproductive performance of short- and medium-chain fatty acids are relatively limited, and the related mechanism is not clear. In addition, the effect of the combined use of different fatty acids on reproductive performance is lacked.
Our previous study demonstrated that the supplementation of short- and medium-chain fatty acids in rat diet could improve embryo implantation [25]. However, the effects of a combination of short- and medium-chain fatty acids on the function of granulosa cells and endometrial cells in early pregnancy are unknown, as are the effects on intestinal and vaginal microbial composition in early pregnancy. The aim of this study was to investigate whether a rational combination of functional short- and medium-chain fatty acids could provide a new nutritional strategy for ensuring successful pregnancy by improving the function of gestational ovarian granulosa cells and endometrial cells while regulating the microbiological environment of the intestine and vagina in early pregnancy, thus providing stable hormone levels and a favorable attachment environment in early pregnancy.
2. Materials and Methods
2.1. Porcine Primary Cultured Ovarian Granulosa Cells
Porcine ovaries were purchased from a Hebei slaughterhouse (China) and transported to the laboratory at 37 °C in NaCl (0.9%) within 30 min. Ovaries (n = 60) were rinsed at 37 °C in 0.9% NaCl with 2% penicillin–streptomycin solution (0.1 mg/mL streptomycin + 10 mg/mL penicillin) (Gibco) until no blood was visible. The follicles with an estimated diameter greater than 5 mm (visually measured) were opened with a 5 mm syringe and 20-G needle (sterile). The follicular fluid (FF) and cumulus-oocyte complexes (COCs) were transferred to several sterile centrifuge tubes (15 mL) and then centrifuged at 200× g for 5 min, followed by the removal of the remnant supernatant. After that, 5 mL of PBS containing 10% penicillin–streptomycin solution at 37 °C was added to the centrifuge tubes, and the pellet was carefully resuspended and cleaned. Granulosa cells (GCs) were extracted through centrifugation (200× g, 5 min), then seeded onto culture dishes. Dulbecco’s modified Eagle’s medium (DMEM) (high glucose) (Sigma–Aldrich, St. Louis, MO, USA), fetal bovine serum (FBS) (Gibco, Carlsbad, CA, USA), 10 mg/mL penicillin and 0.1 mg/mL streptomycin were the components in the culture medium used. Cell cultivation was conducted at 37 °C under 5% CO2 aerobic conditions for 3 h, and then the medium was replaced with fresh culture medium. Next, the cells were cultured for 2 days and treated with 1 mM SB (sodium butyrate, Sigma-Aldrich), 0.04 mM SC (sodium caprylate, Sigma-Aldrich), 0.01 mM SL (sodium laurate, Sigma-Aldrich) and a combination of all three (MSFAs,1 mM SB + 0.04 mM SC + 0.01 mM SL) for 24 h; the optimal concentrations of these three fatty acids were obtained with a CCK-8 kit. The experiment was repeated three times, each with three parallels.
2.2. Cell Viability Assay
With different concentrations of SB (0, 0.1, 0.4, 0.6, 1.0 and 1.4 mM) and SC (0, 0.2, 0.4, 0.6, 0.8 and 1.0 mM), and SL (0, 0.005, 0.008, 0.01, 0.05 and 0.1 mM) treated PGCs for 24 h. The cell activity of PGCs under different treatments was detected according to the instructions of CCK-8 kit (Abcam, Shanghai, China). After testing, we found that 1.0 mM SB, 0.4 mM SC, 0.01 mM SL was the optimum concentration (Figure S1) and used for subsequent cell assays.
2.3. Sex Hormone and Blood Biochemistry Detection
Commercially available radioimmunoassay kits (Cusabio) (Beijing North Institute of Biotechnology, Beijing, China) were used to detect the progesterone and estradiol concentrations in PCG culture supernatants. Total cholesterol, HDL cholesterol and LDL cholesterol were measured using kits from Huaying Institute of Biotechnology (Beijing, China).
2.4. Cell Culture
Porcine endometrial epithelial cells (PEECs) were used to study endometrial receptivity and cultured in medium [90% Dulbecco’s modified Eagle’s medium (DMEM) (high glucose) and 10% FBS (Gibco)] and conducted at 37 °C in a humidified 5% CO2 incubator. Cells serum was starved before treatment. The cells were also treated with 1 mM SB, 0.04 mM SC, 0.01 mM SL and MSFAs for 24 h, and the experiment was repeated three times, each with three parallels. PEECs is a cell line that was kindly provided by Dr. Guoyao Wu from China Agricultural University.
Dulbecco′s Modified Eagle′s Medium/Nutrient Mixture F-12 Ham (DMEM/f-12) supplemented with 10% FBS (Gibco) and 1% insulin, transferrin and selenium solution (ITS-G) (Gibco) was used to culture the porcine trophoblast cells (pTrs), and the cells were maintained at 37 °C in a humidified 5% CO2 incubator. The pTr cells were purchased from the European Collection of Cell Cultures.
2.5. qPCR Analysis
Quantitative PCR (qPCR) was performed according to previously described methods [25]. Primers were designed using Primer 3.0 plus input online (Table 1).

Table 1.
Primers (pig) used for qPCR.
2.6. Adhesion Assay
PEECs (1.5 × 106 cells) were cultured in 6-well plates for 24 h. The medium was replaced and incubated in DMEM (high glucose) with different fatty acids (1 mM SB, 0.4 mM SC, 0.01 mM SL, MSFAs) for 24 h. pTr cells were labeled with fluorescent dye 5-chloromethylfluoresceindiacetate (CMFDA) (CellTracker Green; Invitrogen, Carlsbad, CA, USA) for 10 min at 37 °C. Then, the labeled pTr cells were cleaned in 1 × phosphate-buffered saline and incubated with PEECs for 30 min at 37 °C. The cells were vigorously washed to remove nonattached pTr cells. The attached pTr cells were observed with a fluorescence microscope (Axioimager M1 microscope, Zeiss, Aalen, Germany), and the cell adhesion rates were calculated [26].
2.7. Animals and Experimental Design
The animal procedures described in this experiment were approved by the Institutional Animal Care and Use Committee of China Agricultural University (CAU 20161108-2), and animal experiments were finished in the FengNing Swine Research Unit of China Agricultural University (Academician Workstation in Chengdejiuyun) Agricultural and Livestock Co., Ltd., Hebei, China.
To confirm whether maternal fatty acid supplementation during early pregnancy improved reproduction in sows, among 60 Large White × Landrace crossbred sows with similar body weight (BW) and backfat thickness (BF) were used in this study. After weaning, sows were randomly allocated into two groups based on BW, BF and parity. Sows were feed singly in gestation stalls (2.2 m × 0.65 m); using direct boar exposure to check daily for estrus in the morning, artificial insemination was performed during estrus (12 h apart). Sows were not used in this experiment if they returned to estrus after insemination. In the control group, sows were fed basal diets from the time of weaning to the early pregnancy period (days 0 to 28 of gestation). In the treatment group, sows were fed the basal diet supplemented with 0.1% sodium butyrate, 0.04% sodium caprylate, and 0.1% sodium laurate (purity > 98%, Newgrun Biotechnology, Changsha, China). After day 28 of gestation, all groups were fed the same basal diet. All sows were fed 2.00 kg of the diet during weaning to the end of breeding, 1.25 kg of the diet during early pregnancy and 1.75 kg of the diet from day 29 to delivery and fed twice daily at 06:30 h and 16:30 h. Sows drank water freely throughout the experiment. Diets met the nutritional requirements of pregnant sows (GB/T 39235-2020 Nutritional Requirements for Swine) (Table 2), and the dietary information for diets with fat supplementation is showed in Table S1. Sows’ body weight and back fat were measured on days 0 and 28 of gestation. On days 14 and 28 of gestation, after cleaning, microbes from the vagina and rectum of sows were collected with sterile cotton swabs (Baiao Biological Technology, Shenzhen, China) and immediately frozen in liquid nitrogen. On day 28 of pregnancy, after overnight starvation, 10 sows were randomly selected from each group for blood sample collection via jugular venipuncture into 5 mL tubes, and the samples were centrifuged at 3000× g for 10 min at 4 °C to obtain serum samples. Shortly after birth, litter size, live litter size, litter weight, number of mummified and stillborn piglets and birth weight were recorded.
Table 2.
Composition and Nutrient Composition of the Basal Diet for Sows (As-Fed Basis).
2.8. Bacterial DNA Extraction and 16S rRNA Gene Sequencing
Microbial community genomic DNA was extracted from vaginal and rectal swabs using the QIAmp DNA stool mini kit (Qiagen, GmbH, Hilden, Germany). The DNA extract was checked by agarose gel electrophoresis, and DNA concentration and purity were detected by NanoDrop 2000 UV–vis spectrophotometer (Thermo Scientific, Wilmington, NC, USA). The PCR primers flanking the V3-V4 of the bacterial 16S rDNA gene were amplified with primer 338F (5′-ACTCCTACGGGAGGCAGCAG-3′), and the reverse primer was 806R (5′-GGACTACHVGGGTWTCTAAT-3′) using an ABI GeneAmp® 9700 PCR thermocycler (ABI, Vernon, CA, USA). The optimized conditions for PCR amplification of the 16S rRNA gene were follows: 1 pre-denaturation cycle at 95 °C for 3 min, then 27 cycles of denaturing at 95 °C for 30 s, annealing at 55 °C for 30 s, elongation at 72 °C for 45 s and a single extension at 72 °C for 10 min, ending at 4 °C. Following the standard protocols by Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China), the purified amplicons were pooled in equimolar amounts and paired-end sequenced on an Illumina MiSeq PE300 platform/NovaSeq PE250 platform (Illumina, San Diego, CA, USA); fastp version 0.20.0 [27] was used to demultiplexed raw 16S rRNA gene sequencing reads, quality filtered by and merged in FLASH version 1.2.7 [28] with the following criteria: (i) the 300 bp reads were truncated at any site receiving an average quality score of <20 over a 50 bp sliding window, and the truncated reads shorter than 50 bp were discarded.
Operational taxonomic units (OTUs) with a 97% similarity cutoff [29,30] were clustered using UPARSE version 7.1 [29], and chimeric sequences were identified and removed. The taxonomy of each OTU representative sequence was analyzed in RDP Classifier version 2.2 [31] against the 16S rRNA database (Silva v138) using a confidence threshold of 0.7. We performed α- and β-diversity calculations and taxonomic community assessments using Qiime2-2020.11 scripts. The alpha diversity of the samples was calculated with the observed species, Chao 1 and Shannon indexes. Principal coordinate analysis (PCoA) based on the weighted UniFrac distance was used to summarize the beta diversity. Linear discriminant analysis coupled with effect size (LEfSe) was used to identify the key bacterial taxa between different treatments [32].
2.9. Statistical Analysis
The results are expressed as the mean ± standard error of the mean (SEM). Statistical analysis was performed using a nonpaired Student’s t test or one- or two-way ANOVA in Statistical Product and Service Solutions (SPSS) software (version 26.0.0, IBM, Armonk, NY, USA). Tukey’s post hoc test was applied to determine the differences among treatment means, and a probability of p ≤ 0.05 was considered significant. Linear discriminant analysis (LDA) effect size (LEfSe) was calculated in R 4.0 to identify enriched OTUs, the threshold of the logarithmic score of LDA was set to 2.0, and p ≤ 0.05 and LDA > 2.0 were considered significant across different stages of gestation. The data were visualized using R 4.0 and GraphPad Prism 8.1 (GraphPad Software, San Diego, CA, USA).
3. Results
3.1. MSFAs Can Increase the Expression Levels of Steroid Steroidogenesis Genes and Luteinizing Genes and Promote Estrogen Synthesis
As shown in Figure 1A, the relative mRNA abundance of cytochrome P450 family 19 subfamily A member 1 (Cyp19a1), follicle-stimulating hormone receptor (Fshr), luteinizing hormone receptor (Lhr), peroxisome proliferator activated receptor gamma (Pparγ) and steroidogenic acute regulatory protein (Star) were higher in the MSFA, SB and SC groups than in the control and SL groups. In the SB group, the relative mRNA abundance of Cyp11a1 was higher than that in the control group and SC group, but there was no significant difference compared with the abundance in the MSFA and SL groups. Compared with the SC group, the relative mRNA abundance of Cyp19a1, Pparγ, and Fshr was higher in the MSFA and SB groups. However, in the MSFA group, the relative mRNA abundance of Lhr was higher than that in the other groups.
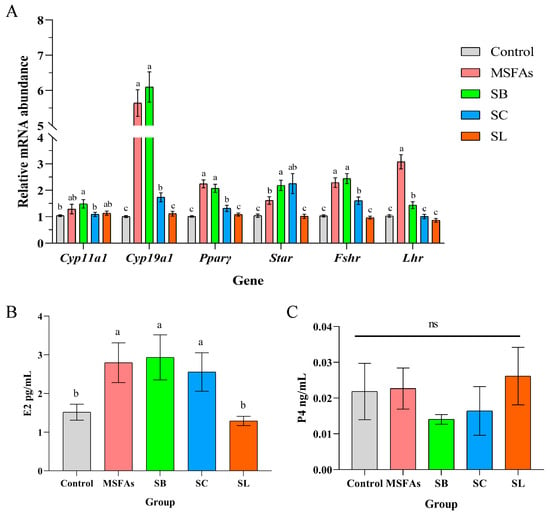
Figure 1.
MSFAs can increase the expression levels of steroid steroidogenesis genes and luteinizing genes and promote estrogen synthesis. (A) The relative mRNA abundance in porcine ovarian granulosa cells treated with 1 mM SB, 0.4 mM SC, 0.01 mM SL or their combination for 24 h. (B) Concentrations of progesterone and (C) estradiol in the supernatant of porcine ovarian granulosa cells treated with different short-chain fatty acids, medium-chain fatty acids or their combination for 24 h. 1 mM SB; 0.4 mM SC; 0.01 mM SL. The results are presented as the means ± SEM. n = 9. Different letters between bars indicate p ≤ 0.05 by one-way ANOVA followed by post hoc Tukey’s tests. ns, nonsignificant (p > 0.05). SB, sodium butyrate; SC, sodium caprylate; SL, sodium laurate; MSFAs, SB + SC + SL.
Based on the changes in the relative mRNA abundance of the genes related to hormone synthesis in cells, we examined the progesterone and estradiol concentrations in the supernatant and found that there was no significant influence on progesterone concentrations in any group but that the MSFA, SB and SC groups showed significant improvement in the cell supernatant concentrations of estradiol, which was consistent with the qPCR results (Figure 1B,C). Taken together, our results demonstrated that MSFAs could promote steroidogenesis and the expression of LH and FSH receptor-related genes and enhance the secretion of estrogen in PGCs.
3.2. MSFAs Can Improve the Receptivity of Pig Endometrial Cells
Improvement in the receptivity of pig endometrial cells is beneficial to early embryo implantation and can effectively improve the reproductive performance of sows. We measured the relative mRNA abundance of endometrial receptivity genes in porcine endometrial cells treated with different short-chain fatty acids and their combinations. As shown in Figure 2A, compared with the control group, MSFA supplementation significantly improved the relative mRNA expression levels of epidermal growth factor (Egf), homeobox A10 (Hoxa10), integrin subunit beta 3 (ανβ3) and leukemia inhibitory factor (Lif). SB supplementation improved the expression of ανβ3 and Lif, SC treatment improved the expression of Egf and Hoxa10, and SL treatment only improved the expression of Hoxa10. There were no differences in the relative mRNA abundance of Egf in the MSFAs, SB and SL groups, but the expression in the SC group was higher than that in the SB and SL groups. In both the MSFAs and SC groups, the expression of Hoxa10 was higher than that in the other groups. However, the expression of ανβ3 and Lif in the MSFA and SB groups was significantly increased than that in the other groups.
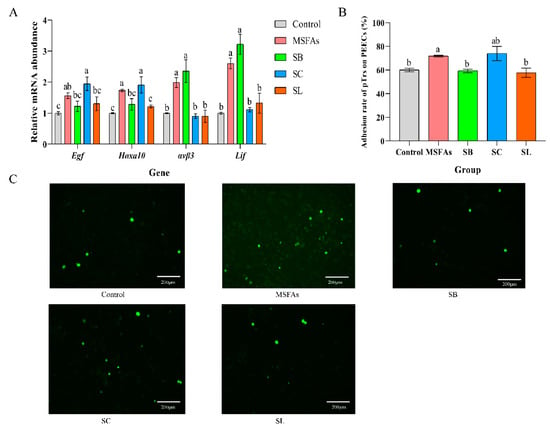
Figure 2.
The combination of MSFAs can improve the receptivity of pig endometrial cells. (A) The relative mRNA abundance in porcine endometrial epithelial cells treated with 1 mM SB, 0.4 mM SC, 0.01 mM SL or their combination for 24 h. (B) Adhesion rate of porcine trophoblast cells to porcine endometrial epithelial cells treated with 1 mM SB, 0.4 mM SC, 0.01 mM SL or their combination for 24 h. (C) Image of adhesion of porcine trophoblast cells to porcine endometrial epithelial cells treated with 1 mM SB, 0.4 mM SC, 0.01 mM SL or their combination for 24 h observed under fluorescence microscope. The results are presented as the means ± SEM. n = 9. Different letters between bars indicate p ≤ 0.05 by one-way ANOVA followed by post hoc Tukey’s tests. ns, nonsignificant (p > 0.05). SB, sodium butyrate; SC, sodium caprylate; SL, sodium laurate; MSFAs, SB + SC + SL.
As shown in Figure 2B, the combination of medium- and short-chain fatty acids improved the rate of adhesion of trophoblast cells to endometrial cells, which was consistent with the RT–PCR results. Collectively, these results indicated that MSFAs can improve the receptivity of endometrial cells in sows.
3.3. Effects of Dietary Supplementation with MSFAs on the Reproductive Performance of Sows
As shown in Table 3, four sows in both the control and treatment groups failed to fertilize during pregnancy; additionally, dietary supplementation of MSFAs in the estrus and early gestation had no effect on the body weight and backfat thickness of sows (p > 0.05). Dietary supplementation of short-chain fatty acids in the estrus and early gestation of sows tended to increase the total litter size (p = 0.078). Compared with that in the control group, the number of live litters born in the treatment group was significantly increased (p < 0.01), and there was also an increasing trend observed for the litter weights of live litters (p = 0.09). Treatment had no effect on average birth weight, sex, offspring strength, stillbirth occurrence, deformation or mummification (p > 0.05) (Table 4).
Table 3.
Effects of medium- and short-chain fatty acid combination on body condition of sows 1,2.
Table 4.
Effects of medium- and short-chain fatty acids on the reproductive performance of sows 1,2.
3.4. Effects of Dietary Supplementation of MSFAs on Serum Steroid Hormones and Serum Metabolites at Day 28 of Gestation
As shown in Table 5, dietary supplementation with the medium-/short-chain fatty acid combination in the treatment group significantly increased the estradiol content in the serum of sows on day 28 of gestation (p < 0.05) but had no effect on cholesterol synthesis related to serum lipid metabolism in the sows (p > 0.05).
Table 5.
Effects of medium- and short-chain fatty acids on reproductive performance of sows 1,2.
3.5. Effects of Dietary Supplementation with MSFAs on the Rectal Swab Microbial Diversity and Composition of Sows at 14 and 28 Days of Gestation
The microbiota diversity and compositions of rectal swab samples in sows at 14 and 28 days of gestation were determined by deep sequencing the 16S rRNA genes. To assess rectal swab microbial community structure, we calculated the alpha and beta diversity values (Figure 3). For richness indexes (observed species, Chao1), dietary supplementation with MSFAs had no effect (p > 0.05). On the other hand, beta diversity is shown in Figure 3. PCoA clustering revealed that the rectal swab microbiota of sows was not significantly dispersed in the control and treatment groups (p > 0.05). In conclusion, the addition of MSFAs to the diet did not effectively change the rectal swab microbiota diversity of sows at 14 or 28 days of gestation.
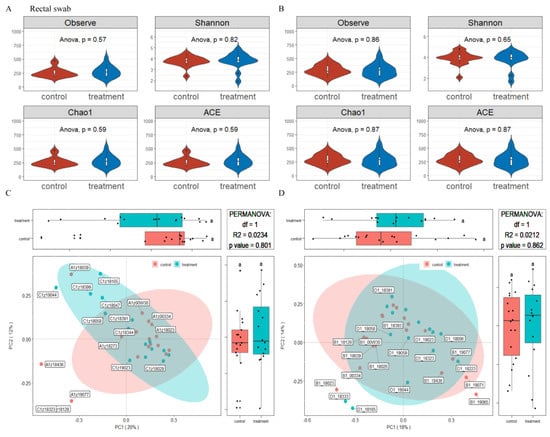
Figure 3.
Effects of MSFAs during gestation on the rectal swab microbiota diversity of sows. Comparison of the number of observed species and Shannon, Chao1 and ACE indexes between the control and treatment groups at day 14 (A) and day 28 (B) of gestation. Beta diversity of the fecal microbiota in sows between the control and treatment groups at day 14 (C) and day 28 (D) of gestation based on weighted UniFrac distance. Data are expressed as the mean ± SEM (n = 11).
The relative abundances at the phylum and genus levels in the rectal swab samples of all groups across different days are shown in Figure 4. The top six phyla were Firmicutes, Bacteroidota, Proteobacteria, Actinobacteriota, Fusobacteriota and Campylobacterota (Figure 4A). These six phyla accounted for more than 99% of the reads for all sows. In addition, the top six genera in the gut microbiota of sows were Escherichia-Shigella, Clostridium_sensu_stricto_1, Porphyromonas, Prevotellaceae, Anaerococcus and Romboutsia (Figure 4B). The microbial compositions of the rectal swab samples were further analyzed by LEfSe. The LefSe analysis showed that Burkholderia_Caballeronia_Paraburkholderia, Lachnospiraceae_NK4A136_group, Rhodococcus and Micrococcus were enriched in the control group at day 14 of gestation; only Collinsella was enriched in the MSFAs group at day 14 of gestation; Weissella, Trueperella, Desulfovibrio and Veillonella were enriched in the control group at day 28 of gestation; and Arcanobacterium and Candidatus_Saccharimonas were enriched in the MSFAs group at day 28 of gestation (Figure 5). Thus, dietary supplementation with medium- and short-chain fatty acids in sows during early gestation had no significant effect on the diversity of rectal swab microbes in the sows, and only some microbes at the genus level showed significant differences.
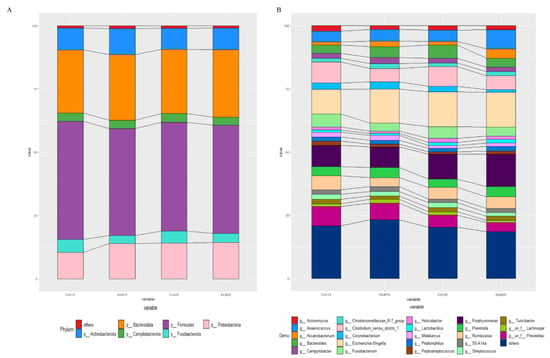
Figure 4.
Effects of MSFAs during gestation on the rectal swab microbiota compositions of sows. The relative abundances of different phyla (A) and genera (B) in the rectal swab microbiota of control and treatment group sows at day 14 and day 28 of gestation. N = 11.
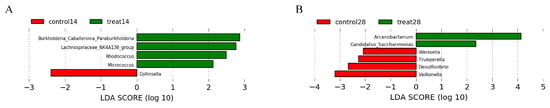
Figure 5.
Effects of dietary supplementation with MSFAs on the rectal swab microbial diversity and compositions of sows at 14 and 28 days of gestation. Linear discriminant analysis coupled with effect size (Lefse) of day 14 (A) and day 28 (B) rectal swab microbiota compositions in the control and treated sows. Control14 (28), day 14 (28) of gestation and fed the basal diet. Treat14 (28), day 14 (28) of gestation and fed a basal diet supplemented with 0.1 SB, 0.05 SC and 0.1% SL.
3.6. Effects of Dietary Supplementation with MSFAs on Vaginal Microbial Diversity and Composition of Sows at 14 and 28 Days of Gestation
The microbiota diversity and compositions of vaginal samples in sows at 14 and 28 days of gestation were determined in the same way as in rectal swab samples. As showed in Figure 6, the alpha diversity values were no effect in dietary supplementation with MSFAs (p > 0.05). PCoA clustering revealed that the vaginal microbiota of sows was not significantly dispersed in the control and treatment groups (p > 0.05). Similarly, the addition of MSFAs to the diet also did not effectively change the vaginal microbiota diversity of sows at 14 or 28 days of gestation.
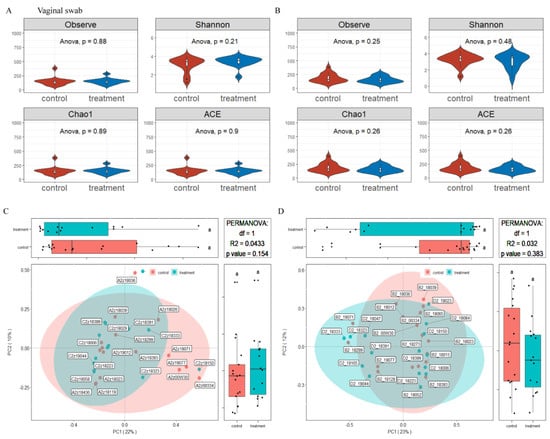
Figure 6.
Effects of MSFAs during gestation on the vagina microbiota diversity of sows. Comparison of the number of observed species and Shannon, Chao1 and ACE indexes between the control and treatment groups at day 14 (A) and day 28 (B) of gestation. Beta diversity of the fecal microbiota in sows between the control and treatment groups at day 14 (C) and day 28 (D) of gestation based on weighted UniFrac distance. Data are expressed as the mean ± SEM (n = 11). Day 14 of gestation n: control = 12, treatment = 13, Day 8 of gestation n: control = 18, treatment = 15.
The relative abundances at the phylum and genus levels in vaginal samples of all group across different days are shown in Figure 7. The top six phyla were Firmicutes, Proteobacteria, Bacteroidota, Actinobacteriota, Campylobacterota and Fusobacteriota (Figure 7A). These six phyla accounted for more than 99% of the reads for all sows. In addition, the top six genera in the gut microbiota of sows were Porphyromonas, Escherichia-Shigella, Acinetobacter, Enterococcus, Gallicola and Corynebacterium (Figure 7B). The microbial compositions of the rectal swab samples were further analyzed by LEfSe. The results showed that Pseudomonas, Delftia, Macrococcus, Rothia and Escherichia-Shigella were enriched in the control group at day 14 of gestation; Fusobacterium, Anaerococcus, TM7a, Helcococcus, Veillonella, DNF00809, Allofustis, Moryella, Sarcina and Christensenellaceae_R_7_group were enriched in the MSFA group at day 14 of gestation; Prevotellaceae_NK3B31_group, Iamia, Brevibacterium, Lachnospiraceae_XPB1014_group, Desulfovibrio, Dietzia, Truepera, Micrococcus, Clostridum_sensu_stricto_1, Rhodococcus and Burkholderia_Caballeronia_Paraburknolderia were enriched in the control group at day 28 of gestation; and Macrococcus, Ferruginibacter, Ruminococcus_gnavus_group, Negativicoccus, Alloprevotella, Muribaculaceae, Empedobacter and Lachnospiraceae_NK4A136_group were enriched in the MSFA group at day 28 of gestation (Figure 8). Similarly, dietary supplementation with medium- and short-chain fatty acids in sows during early gestation had no significant effect on the diversity of the vaginal microbes of sows, although there were more differences in genus-level microbes between the control and MSFA groups at day 14 or 28 of gestation.
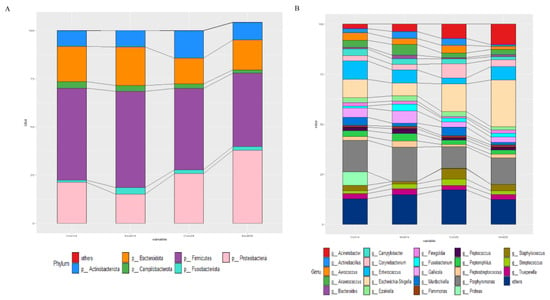
Figure 7.
Effects of MSFAs during gestation on vaginal microbiota composition of sows. The relative abundances of different phyla (A) and genera (B) in the vaginal microbiota of control and treatment group sows at day 14 and day 28 at gestation.
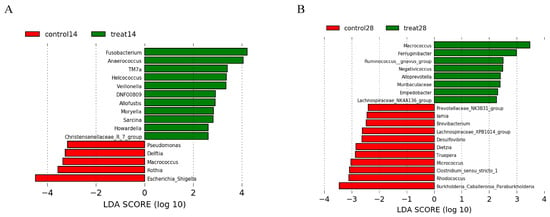
Figure 8.
Effects of dietary supplementation with MSFAs on vaginal microbial diversity and composition of sows at 14 and 28 days of gestation. Linear discriminant analysis coupled with effect size (Lefse) of day 14 (A) and day 28 (B) vaginal microbiota composition of control and treated sows. Control14 (28), day 14 (28) of gestation and fed the basal diet. Treat14 (28), day 14 (28) of gestation and fed a basal diet supplemented with 0.1 SB, 0.05 SC and 0.1% SL.
4. Discussion
Early embryo loss is one of the key factors limiting successful reproduction in mammals [33,34]. Optimal hormone levels and implantation conditions, strongly influenced by maternal nutritional status, can effectively reduce early embryo loss. Supplementation with individual short- or medium-chain fatty acids is reported to improve maternal hormone levels and energy status [23,24,25,35,36]. However, it is still unknown whether the proportional combined use of these short- and medium-chain fatty acids achieves better effects. In the present study, our results showed that short- and medium-chain fatty acid supply strengthened steroidogenesis and luteinization processes in porcine granulosa cells by enhancing the expression of critical genes involved in these processes, including Cyp19a1, Star, Pparγ, Fshr and Lhr. In addition, short- and medium-chain fatty acids enhanced the receptivity of porcine endometrial epithelial cells. Furthermore, dietary supplementation with short- and medium-chain fatty acids improved the estrogen level of sows in early pregnancy, improved the reproductive performance of sows and provided a good intestinal and vaginal microbial environment.
Maintaining ovarian steroidogenesis is an important event for ovarian function and successful pregnancy. Research has shown that estrogens are important to primordial follicle formation and maintaining follicular development, through stimulating the final stage of ovulation and granulosa cell proliferation, promoting follicle-stimulating hormone-stimulated gene expression and even affecting the function and structure of female reproductive tissue [37,38,39,40]. Our previous study showed that short-chain fatty acids such as sodium butyrate and sodium caprylate can affect ovarian steroidogenesis and granulosa cell luteinization in rats [25]. In this study, we detected the effects of different concentrations of SB, SC and SL on PGCs cell activity by CCK-8 kit and selected the most appropriate concentration for subsequent tests (Figure S1). Although the actual physiological concentration of these fatty acids in follicles is much lower than our optimal concentration, the optimal concentration of these fatty acids did not negatively affect cell activity in vitro. Our data indicated that treatment with sodium butyrate or sodium caprylate could elevate the expression of Cyp19a1, which is a critical gene for estrogen synthesis and the prevention of cell apoptosis, in porcine granulosa cells [41]. In addition, it has been shown that both FSH and LH strongly stimulate Cyp19a1 mRNA expression and estradiol and progesterone production in most mammalian mature granulosa cells by binding to the specific receptors Fshr and Lhr, respectively [42,43,44,45]. However, the impact of the combined use of short- and medium-chain fatty acids on the expression levels of Lhr and Fshr genes in granulosa cells has not been reported. Herein, our data showed that the combined use of short- and medium-chain fatty acids increased the mRNA level of the Lhr gene more effectively than sodium butyrate or sodium caprylate treatment, while sodium laurate had no effect on the expression of these genes. In addition, the combined use of short- and medium-chain fatty acids significantly increased the supernatant estrogen levels in porcine granulosa cells, while sodium butyrate or sodium caprylate treatment did not have such an effect, suggesting that the joint use of these fatty acids can more effectively promote estrogen secretion in porcine granulosa cells. Furthermore, our in vivo data indicated that dietary supplementation with MSFAs could significantly increase the serum estrogen level of sows during early pregnancy, indicating that the combined use of short- and medium-chain fatty acids could indeed strengthen ovarian estrogen secretion in sows during early pregnancy.
Embryo implantation is another decisive event for early embryo survival. During embryo implantation, approximately two thirds of implantation failures are related to poor uterine receptivity, while only one third of these failures are related to the embryo itself [46,47]. The biomarkers of uterine receptivity include Hoxa10, Lif, αvβ3 integrin and Egf. Hoxa10 is a homeobox gene required for endometrial receptivity to blastocyst implantation. Hoxa10 has been reported as important to the endometrium by regulating endometrial stromal cell proliferation and epithelial cell morphogenesis [48]. In addition, it has been demonstrated that Hoxa10 is related to pinopod development and is a morphological marker of high endometrial receptivity [49,50,51]. Blocking Hoxa10 expression significantly reduces the number of endometrial pinopods. It has also been reported that the development of endometrial pinopods is associated with the increased expression of Lif and its receptors [52] and of ανβ3 integrin [53]. In fact, the expression of αvβ3 integrin in the human endometrium can be regulated by Egf [54]. Moreover, Egf is a potent inducer of Lif expression [55]. Our data demonstrated that sodium butyrate treatment could significantly increase the mRNA abundance of the ανβ3 and Lif genes, while sodium caprylate treatment could increase the expression levels of Egf and Hoxa10. Sodium laurate only promotes the expression of Hoxa10. In contrast, the combined use of short- and medium-chain fatty acids significantly enhanced the endometrial expression levels of Hoxa10, ανβ3, Lif, and Egf. These results indicated that each fatty acid had different impacts on the biomarkers of endometrial receptivity. More importantly, the combined use of short- and medium-chain fatty acids was much more efficient in improving endometrial receptivity. These beneficial effects of the combined use of short- and medium-chain fatty acids were confirmed to effectively improve the embryo adhesion efficiency in vitro, and these beneficial effects were not due to differences in fat content but due to the specific types of fatty acid. Our previous data demonstrated that butyrate supplementation in the pregnant diet could not enhance the citrate cycle, and maintained the ATP levels and AMPK signaling, indicating that this level of butyrate supplementation could not influence the energy metabolism [56]. Our previous data also indicated that dietary butyrate supplementation during pregnancy primarily acted to promote the ovary function [25]. Therefore, we speculated that these functional fatty acids promoted the reproductive performance through enhancing the ovary function and uterine receptivity by not directly acting as the energy enhancing.
Notably, in addition to the improvement in ovarian steroidogenesis and endometrial receptivity, the combined use of short- and medium-chain fatty acids markedly inhibited specific pathogenic bacteria in the feces and vagina of sows during early pregnancy. It is well known that the gut and vaginal microbiomes during pregnancy have an impact on reproductive performance [57,58]. Gut microbiota imbalance can lead to pregnancy complications and adverse pregnancy outcomes, as well as polycystic ovary syndrome and endometriosis [58]. The vaginal microbiome is a key regulator of local inflammatory and immune pathways throughout pregnancy [59,60,61]. Studies have shown that dietary supplementation with sodium butyrate can promote intestinal health by modulating the microbial community in mice, piglets and poultry [62,63,64] and can also help to inhibit colitis and endometritis in mice [62,65,66]. Similarly, dietary supplementation with caprylic acid can significantly inhibit colonization by pathogenic bacteria (e.g., Salmonella enteritidis [67] and Campylobacter [68]) in the intestine of poultry. Moreover, caprylic acid can be used to treat monilial vaginitis [69]. In particular, lauric acid has been reported to have strong antibacterial activity, such as Staphylococcus aureus, Streptococcus mutans, S. pyogenes, Escherichia coli, and H. pylori [70,71]. In this study, the combined use of short- and medium-chain fatty acids in diets did not affect the diversity of intestinal and vaginal microorganisms in sows during early gestation; however, it significantly decreased, at the genus level, the rectal swab abundance of Trueperella and vaginal abundance of Escherichia-Shigella and Clostridium_sensu_stricto_1 in sows. Numerous studies have reported that two species of Trueperella, Trueperella pyogenes and Trueperella abortisuis, are closely associated with vaginitis and abortion. Trueperella pyogenes is a well-known pathogen in domestic ruminants and pigs that causes mastitis and a variety of pyogenic infections [72]. Recently, Trueperella pyogenes was reported to be significantly associated with metritis in postpartum cows [73]. The high abundance of the Trueperella abortisuis strain seems to be responsible for abortion in farms [74]. In addition, the abundances of Escherichia-Shigella and Clostridium_sensu_stricto_1 are relatively high in endometriotic sows [75]. Therefore, it is speculated that the combined use of short- and medium-chain fatty acids in diets protects against gut and vaginal pathogenic bacteria in sows during early pregnancy.
5. Conclusions
In conclusion, we found that the combined use of short- and medium-chain fatty acids improved the reproductive performance of sows associated with strengthening steroidogenesis and luteinization in ovarian granulosa cells and improving endometrial cell receptivity, while the treatment significantly decreased several specific pathogenic bacteria in the feces and vagina of sows during early pregnancy. These findings imply that the combined use of short- and medium-chain fatty acids has great potential to improve pregnancy outcomes in humans and other mammals.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/nu14204405/s1, Figure S1: CCK-8 assay of PGCs treated with different concentration of SB (A), SC (B), SL (C) for 24 h; Figure S2: Fatty acid composition in serum of sows at 28 days of gestation under different treatments.; Table S1: Dietary information for diets with fat supplementation.
Author Contributions
The authors’ responsibilities were as follows—X.Z. (Xiangfang Zeng), S.Q. (Shiyan Qiao), F.W., C.C., S.Z. and F.C.: conceived and designed the whole trial; X.Z. (Xiangzhou Zeng), S.L., Q.Y. and L.L.: conducted laboratory analyses; X.Z. (Xiangzhou Zeng) and S.Q. (Shuang Quan): conducted the Sows trial; X.Z. (Xiangzhou Zeng) and S.C.: wrote the manuscript; and all authors: read and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Natural Science Foundation of China, Grant/Award Number: 32022080 and S&T Program of Hebei (199A7310H).
Institutional Review Board Statement
The present study involved an in vitro digestion experiment and an animal experiment. Animal experiments were approved by the China Agricultural University Animal Care and Use Committee (Beijing, AW11102202-1-1).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the reported results and conclusions can be found in the submitted figure and tables. Additional research materials and protocols that are relevant to the study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank the participants and medical staff for their participation. This manuscript was polished by AJE website organizations.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results. The opinions expressed here belong to the authors and do not necessarily reflect those of any of the funding agencies or contributing institutions.
References
- Goff, A.K. Embryonic Signals and Survival. Reprod. Domest. Anim. 2002, 37, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J. Oocyte control of ovarian follicular development and function in mammals. Reproduction 2001, 122, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Boland, M.; Lonergan, P.; O’Callaghan, D. Effect of nutrition on endocrine parameters, ovarian physiology, and oocyte and embryo development. Theriogenology 2001, 55, 1323–1340. [Google Scholar] [CrossRef]
- Fleming, R.; Lloyd, F.; Herbert, M.; Fenwick, J.; Griffiths, T.; Murdoch, A. Effects of profound suppression of luteinizing hormone during ovarian stimulation on follicular activity, oocyte and embryo function in cycles stimulated with purified follicle stimulating hormone. Hum. Reprod. 1998, 13, 1788–1792. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Conejero, J.A.; Simón, C.; Pellicer, A.; Horcajadas, J.A. Is ovarian stimulation detrimental to the endo-metrium? Reprod. BioMedicine Online 2007, 15, 45–50. [Google Scholar] [CrossRef]
- Young, S.L. Oestrogen and progesterone action on endometrium: A translational approach to understanding endometrial receptivity. Reprod. Biomed. Online 2013, 27, 497–505. [Google Scholar] [CrossRef]
- Governini, L.; Luongo, F.P.; Haxhiu, A.; Piomboni, P.; Luddi, A. Main actors behind the endometrial receptivity and successful implantation. Tissue Cell 2021, 73, 101656. [Google Scholar] [CrossRef]
- Lu, H.-C.; Mackie, K. An Introduction to the Endogenous Cannabinoid System. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef]
- Weems, C.; Weems, Y.; Randel, R. Prostaglandins and reproduction in female farm animals. Vet. J. 2006, 171, 206–228. [Google Scholar] [CrossRef]
- Aoki, J. Mechanisms of lysophosphatidic acid production. Semin. Cell Dev. Biol. 2004, 15, 477–489. [Google Scholar] [CrossRef]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Sumida, C. Fatty acids: Ancestral ligands and modern co-regulators of the steroid hormone receptor cell signalling pathway. Prostaglandins Leukot. Essent. Fat. Acids 1995, 52, 137–144. [Google Scholar] [CrossRef]
- Herrera, E.; Ortega-Senovilla, H. Maternal lipid metabolism during normal pregnancy and its implications to fetal development. Clin. Lipidol. 2010, 5, 899–911. [Google Scholar] [CrossRef]
- Sturmey, R.G.; Reis, A.; Leese, H.J.; McEvoy, T.G. Role of Fatty Acids in Energy Provision During Oocyte Maturation and Early Embryo Development. Reprod. Domest. Anim. 2009, 44 (Suppl. S3), 50–58. [Google Scholar] [CrossRef]
- Mennitti, L.V.; Oliveira, J.L.; Morais, C.A.; Estadella, D.; Oyama, L.M.; Nascimento, C.M.O.D.; Pisani, L.P. Type of fatty acids in maternal diets during pregnancy and/or lactation and metabolic consequences of the offspring. J. Nutr. Biochem. 2015, 26, 99–111. [Google Scholar] [CrossRef]
- Zhao, Z.-A.; Zhang, Z.-R.; Xu, X.; Deng, W.-B.; Li, M.; Leng, J.-Y.; Liang, X.-H.; Yang, Z.-M. Arachidonic acid regulation of the cytosolic phospholipase A2α/cyclooxygenase-2 pathway in mouse endometrial stromal cells. Fertil. Steril. 2012, 97, 1199–1205.e9. [Google Scholar] [CrossRef]
- Abolghasemi, A.; Dirandeh, E.; Pirsaraei, Z.A.; Shohreh, B. Dietary conjugated linoleic acid supplementation alters the expression of genes involved in the endocannabinoid system in the bovine endometrium and increases plasma progesterone concentrations. Theriogenology 2016, 86, 1453–1459. [Google Scholar] [CrossRef]
- Van Hoeck, V.; Sturmey, R.G.; Bermejo-Alvarez, P.; Rizos, D.; Gutierrez-Adan, A.; Leese, H.J.; Bols, P.E.J.; Leroy, J.L.M.R. Elevated Non-Esterified Fatty Acid Concentrations during Bovine Oocyte Maturation Compromise Early Embryo Physiology. PLoS ONE 2011, 6, e23183. [Google Scholar] [CrossRef]
- Jungheim, E.S.; Louden, E.D.; Chi, M.M.-Y.; Frolova, A.I.; Riley, J.K.; Moley, K.H. Preimplantation Exposure of Mouse Embryos to Palmitic Acid Results in Fetal Growth Restriction Followed by Catch-Up Growth in the Offspring. Biol. Reprod. 2011, 85, 678–683. [Google Scholar] [CrossRef]
- Hague, A.; Elder, D.J.E.; Hicks, D.J.; Paraskeva, C. Apoptosis in colorectal tumour cells: Induction by the short chain fatty acids butyrate, propionate and acetate and by the bile salt deoxycholate. Int. J. Cancer 1995, 60, 400–406. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef] [PubMed]
- Liberato, M.V.; Nascimento, A.S.; Ayers, S.D.; Lin, J.Z.; Cvoro, A.; Silveira, R.L.; Martínez, L.; Souza, P.C.T.; Saidemberg, D.; Deng, T.; et al. Medium Chain Fatty Acids Are Selective Peroxisome Proliferator Activated Receptor (PPAR) γ Activators and Pan-PPAR Partial Agonists. PLoS ONE 2012, 7, e36297. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Fang, Z.-F.; Che, L.-Q.; Xu, S.-Y.; Wu, D.; Wu, C.-M.; Wu, X.-Q. Use of Sodium Butyrate as an Alternative to Dietary Fiber: Effects on the Embryonic Development and Anti-Oxidative Capacity of Rats. PLoS ONE 2014, 9, e97838. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Takanashi, K.; Hamatani, T.; Hirayama, A.; Akutsu, H.; Fukunaga, T.; Ogawa, S.; Sugawara, K.; Shinoda, K.; Soga, T.; et al. A medium-chain fatty acid as an alternative energy source in mouse preimplantation development. Sci. Rep. 2012, 2, 930. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Cai, S.; Wang, S.; Zeng, X.; Ye, C.; Chen, M.; Zeng, X.; Qiao, S. Maternal short and medium chain fatty acids supply during early pregnancy improves embryo survival through enhancing progesterone synthesis in rats. J. Nutr. Biochem. 2019, 69, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-Y.; Choi, H.-J.; Chung, T.-W.; Choi, J.-Y.; Kim, H.S.; Jung, Y.-S.; Lee, S.-O.; Ha, K.-T. Water-extracted Perilla frutescens increases endometrial receptivity though leukemia inhibitory factor-dependent expression of integrins. J. Pharmacol. Sci. 2016, 131, 259–266. [Google Scholar] [CrossRef]
- Ollion, J.; Cochennec, J.; Loll, F.; Escudé, C.; Boudier, T. TANGO: A generic tool for high-throughput 3D image analysis for studying nuclear organization. Bioinformatics 2013, 29, 1840–1841. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Norwitz, E.R.; Schust, D.J.; Fisher, S.J. Implantation and the Survival of Early Pregnancy. N. Engl. J. Med. 2001, 345, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Spencer, T.E.; Johnson, G.A.; Burghardt, R.C.; Wu, G. Comparative aspects of implantation. Reproduction 2009, 138, 195–209. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Wang, M.; Guo, H.; Jia, Y.; Yang, X.; Zhao, R. Effects of sodium butyrate supplementation on reproductive performance and colostrum composition in gilts. Animal 2016, 10, 1722–1727. [Google Scholar] [CrossRef]
- Vieira, E.; Watanabe, P.; Andrade, T.; Araújo, G.; Silva, B.; Pinheiro, R.; Mendonça, I. Dietary supplementation of sodium butyrate for mixed-parity sows during lactation. Livest. Sci. 2020, 232, 103915. [Google Scholar] [CrossRef]
- Zhai, J.; Liu, J.; Cheng, X.; Li, S.; Hong, Y.; Sun, K.; Chen, Z.-J.; Du, Y.; Li, W. Zinc finger gene 217 (ZNF217) Promoted Ovarian Hyperstimulation Syndrome (OHSS) through Regulating E2 Synthesis and Inhibiting Thrombospondin-1 (TSP-1). Sci. Rep. 2017, 7, 3245. [Google Scholar] [CrossRef]
- Kezele, P.; Skinner, M.K. Regulation of Ovarian Primordial Follicle Assembly and Development by Estrogen and Progesterone: Endocrine Model of Follicle Assembly. Endocrinology 2003, 144, 3329–3337. [Google Scholar] [CrossRef]
- Richards, J.S.; Pangas, S.A. The ovary: Basic biology and clinical implications. J. Clin. Investig. 2010, 120, 963–972. [Google Scholar] [CrossRef]
- Greco, T.L.; Payne, A.H. Ontogeny of expression of the genes for steroidogenic enzymes P450 side-chain cleavage, 3 beta-hydroxysteroid dehydrogenase, P450 17 alpha-hydroxylase/C17-20 lyase, and P450 aromatase in fetal mouse gonads. Endocrinology 1994, 135, 262–268. [Google Scholar] [CrossRef]
- Li, Q.; Du, X.; Pan, Z.; Zhang, L.; Li, Q. The transcription factor SMAD4 and miR-10b contribute to E2 release and cell apoptosis in ovarian granulosa cells by targeting CYP19A1. Mol. Cell Endocrinol. 2018, 476, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Simoni, M.; Gromoll, J.; Nieschlag, E. The Follicle-Stimulating Hormone Receptor: Biochemistry, Molecular Biology, Physiology, and Pathophysiology. Endocr. Rev. 1997, 18, 739–773. [Google Scholar] [CrossRef]
- Richards, J.S. Maturation of ovarian follicles: Actions and interactions of pituitary and ovarian hormones on follicular cell differentiation. Physiol. Rev. 1980, 60, 51–89. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, A.J.W.; Adashi, E.Y.; Jones, P.B.C.; Welsh, J.T.H. Hormonal Regulation of the Differentiation of Cultured Ovarian Granulosa Cells. Endocr. Rev. 1984, 5, 76–127. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Yu, Y.; Li, Y.; Wang, S.; Zhang, R.; Guo, Y.; Li, Y.; Yan, Y.; Sun, Y.P. Human chorionic gonadotropin-induced amphiregulin stimulates aromatase expression in human granulosa-lutein cells: A mechanism for estradiol production in the luteal phase. Hum. Reprod. 2019, 34, 2018–2026. [Google Scholar] [CrossRef] [PubMed]
- Simón, C.; Moreno, C.; Remohí, J.; Pellicer, A. Cytokines and embryo implantation. J. Reprod. Immunol. 1998, 39, 117–131. [Google Scholar] [CrossRef]
- Ledee-Bataille, N.; Laprée-Delage, G.; Taupin, J.-L.; Dubanchet, S.; Frydman, R.; Chaouat, G. Concentration of leukaemia inhibitory factor (LIF) in uterine flushing fluid is highly predictive of embryo implantation. Hum. Reprod. 2002, 17, 213–218. [Google Scholar] [CrossRef]
- Bagot, C.N.; Kliman, H.J.; Taylor, H.S. Maternal Hoxa10 is required for pinopod formation in the development of mouse uterine receptivity to embryo implantation. Dev. Dyn. 2001, 222, 538–544. [Google Scholar] [CrossRef]
- Nikas, G.; Develioglu, O.H.; Toner, J.P.; Jones, H.W. Endometrial pinopodes indicate a shift in the window of receptivity in IVF cycles. Hum. Reprod. 1999, 14, 787–792. [Google Scholar] [CrossRef]
- Nardo, L.; Sabatini, L.; Rai, R.; Nardo, F. Pinopode expression during human implantation. Eur. J. Obstet. Gynecol. Reprod. Biol. 2002, 101, 104–108. [Google Scholar] [CrossRef]
- Cavagna, M.; Mantese, J. Biomarkers of Endometrial Receptivity—A Review. Placenta 2003, 24, S39–S47. [Google Scholar] [CrossRef]
- Aghajanova, L.; Stavreus-Evers, A.; Nikas, Y.; Hovatta, O.; Landgren, B.M. Coexpression of pinopodes and leukemia inhibitory factor, as well as its receptor, in human endometrium. Fertil. Steril. 2003, 79 (Suppl. S1), 808–814. [Google Scholar] [CrossRef]
- Lessey, B.A.; Damjanovich, L.; Coutifaris, C.; Castelbaum, A.; Albelda, S.M.; Buck, C.A. Integrin adhesion molecules in the human endometrium. Correlation with the normal and abnormal menstrual cycle. J. Clin. Investig. 1992, 90, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Somkuti, S.G.; Yuan, L.; Fritz, M.A.; Lessey, B.A. Epidermal Growth Factor and Sex Steroids Dynamically Regulate a Marker of Endometrial Receptivity in Ishikawa Cells. J. Clin. Endocrinol. Metab. 1997, 82, 2192–2197. [Google Scholar] [CrossRef] [PubMed]
- Arici, A.; Engin, O.; Attar, E.; Olive, D.L. Modulation of leukemia inhibitory factor gene expression and protein biosynthesis in human endometrium. J. Clin. Endocrinol. Metab. 1995, 80, 1908–1915. [Google Scholar] [CrossRef]
- Ye, Q.; Zeng, X.; Wang, S.; Zeng, X.; Yang, G.; Ye, C.; Cai, S.; Chen, M.; Li, S.; Qiao, S. Butyrate drives the acetylation of histone H3K9 to activate steroidogenesis through PPARγ and PGC1α pathways in ovarian granulosa cells. FASEB J. 2021, 35, e21316. [Google Scholar] [CrossRef]
- Heil, B.A.; Paccamonti, D.L.; Sones, J.L. Role for the mammalian female reproductive tract microbiome in pregnancy out-comes. Physiol. Genom. 2019, 51, 390–399. [Google Scholar] [CrossRef]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1894070. [Google Scholar] [CrossRef]
- Brown, R.G.; Al-Memar, M.; Marchesi, J.R.; Lee, Y.S.; Smith, A.; Chan, D.; Lewis, H.; Kindinger, L.; Terzidou, V.; Bourne, T.; et al. Establishment of vaginal microbiota composition in early pregnancy and its association with subsequent preterm prelabor rupture of the fetal membranes. Transl. Res. 2019, 207, 30–43. [Google Scholar] [CrossRef]
- Al-Memar, M.; Bobdiwala, S.; Fourie, H.; Mannino, R.; Lee, Y.; Smith, A.; Marchesi, J.; Timmerman, D.; Bourne, T.; Bennett, P.; et al. The association between vaginal bacterial composition and miscarriage: A nested case–control study. BJOG Int. J. Obstet. Gynaecol. 2020, 127, 264–274. [Google Scholar] [CrossRef]
- Kindinger, L.M.; MacIntyre, D.A.; Lee, Y.S.; Marchesi, J.R.; Smith, A.; McDonald, J.A.K.; Terzidou, V.; Cook, J.R.; Lees, C.; Israfil-Bayli, F.; et al. Relationship between vaginal microbial dysbiosis, inflammation, and pregnancy outcomes in cervical cerclage. Sci. Transl. Med. 2016, 8, 350ra102. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Gao, N.; Yan, D.; Shan, A. Sodium Butyrate Alleviates Mouse Colitis by Regulating Gut Microbiota Dysbiosis. Animals 2020, 10, 1154. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Xiao, Z.; An, W.; Dong, Y.; Zhang, B. Dietary sodium butyrate improves intestinal development and function by modulating the microbial community in broilers. PLoS ONE 2018, 13, e0197762. [Google Scholar] [CrossRef]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary Sodium Butyrate Decreases Postweaning Diarrhea by Mod-ulating Intestinal Permeability and Changing the Bacterial Communities in Weaned Piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef]
- Zhang, T.; Ding, C.; Zhao, M.; Dai, X.; Yang, J.; Li, Y.; Gu, L.; Wei, Y.; Gong, J.; Zhu, W.; et al. Sodium Butyrate Reduces Colitogenic Immunoglobulin A-Coated Bacteria and Modifies the Composition of Microbiota in IL-10 Deficient Mice. Nutrients 2016, 8, 728. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, Y.; Jiang, P.; Yao, H.; Zhao, C.; Hu, X.; Cao, Y.; Zhang, N.; Fu, Y.; Shen, H. Sodium butyrate alleviates lipopolysaccharide-induced endometritis in mice through inhibiting inflammatory response. Microb. Pathog. 2019, 137, 103792. [Google Scholar] [CrossRef]
- Upadhyaya, I.; Upadhyay, A.; Yin, H.-B.; Nair, M.S.; Bhattaram, V.K.; Karumathil, D.; Kollanoor-Johny, A.; Khan, M.I.; Darre, M.J.; Curtis, P.A.; et al. Reducing Colonization and Eggborne Transmission of Salmonella Enteritidis in Layer Chickens by In-Feed Supplementation of Caprylic Acid. Foodborne Pathog. Dis. 2015, 12, 591–597. [Google Scholar] [CrossRef]
- Santos, F.S.D.L.; Hume, M.; Venkitanarayanan, K.; Donoghue, A.M.; Hanning, I.; Slavik, M.F.; Aguiar, V.F.; Metcalf, J.H.; Reyes-Herrera, I.; Blore, P.J.; et al. Caprylic Acid Reduces Enteric Campylobacter Colonization in Market-Aged Broiler Chickens but Does Not Appear To Alter Cecal Microbial Populations. J. Food Prot. 2010, 73, 251–257. [Google Scholar] [CrossRef]
- Reich, W.J.; Nechtow, M.J.; Kurzon, A.M.; Subotnik, N.; Reich, J.B. The treatment of monilial vaginitis with caprylic acid. Am. J. Obstet. Gynecol. 1953, 65, 180–185. [Google Scholar] [CrossRef]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty Acids and Derivatives as Antimicrobial Agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef]
- Rouse, M.S.; Rotger, M.; Piper, K.E.; Steckelberg, J.M.; Scholz, M.; Andrews, J.; Patel, R. In Vitro and In Vivo Evaluations of the Activities of Lauric Acid Monoester Formulations against Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 3187–3191. [Google Scholar] [CrossRef] [PubMed]
- Lämmler, C.; Hartwigk, H. Actinomyces pyogenes und Arcanobacterium haemolyticum. Handb. Der Bakteriellen Infekt. Bei Tieren 1995, 2, 3. [Google Scholar]
- Ashrafi Tamai, I.; Mohammadzadeh, A.; Zahraei Salehi, T.; Mahmoodi, P. Genomic characterisation, detection of genes encoding virulence factors and evaluation of antibiotic resistance of Trueperella pyogenes isolated from cattle with clinical metritis. Antonie Van Leeuwenhoek 2018, 111, 2441–2453. [Google Scholar] [CrossRef]
- Alssahen, M.; Hassan, A.A.; Wickhorst, J.-P.; Sammra, O.; Lämmler, C.; Glaeser, S.P.; Kämpfer, P.; Timke, M.; Prenger-Berninghoff, E.; Abdulmawjood, A. Epidemiological analysis of Trueperella abortisuis isolated from cases of pig abortion of a single farm. Folia Microbiol. 2020, 65, 491–496. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Nesengani, L.; Gong, Y.; Zhang, S.; Lu, W. Characterization of vaginal microbiota of endometritis and healthy sows using high-throughput pyrosequencing of 16S rRNA gene. Microb. Pathog. 2017, 111, 325–330. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).