
Anti-Inflammatory Effect of Korean Propolis on Helicobacter pylori-Infected Gastric Mucosal Injury Mice Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Propolis Solution
2.2. Bacterial Culture
2.3. Ethics
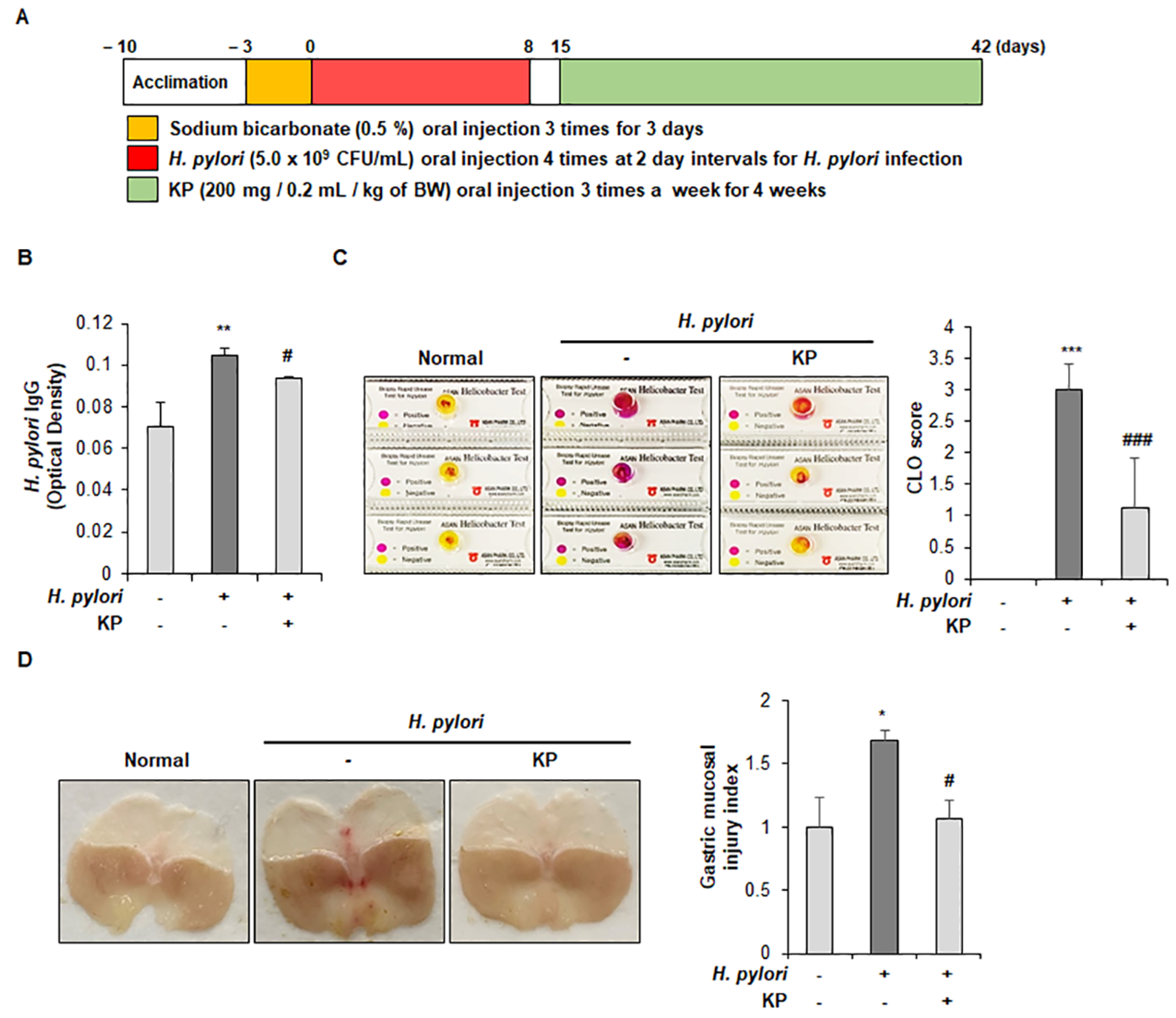
2.4. Animal Experiment
2.5. H. pylori Antigen Test in Mouse Serum
2.6. Campylobacter-Like Organism (CLO) Test
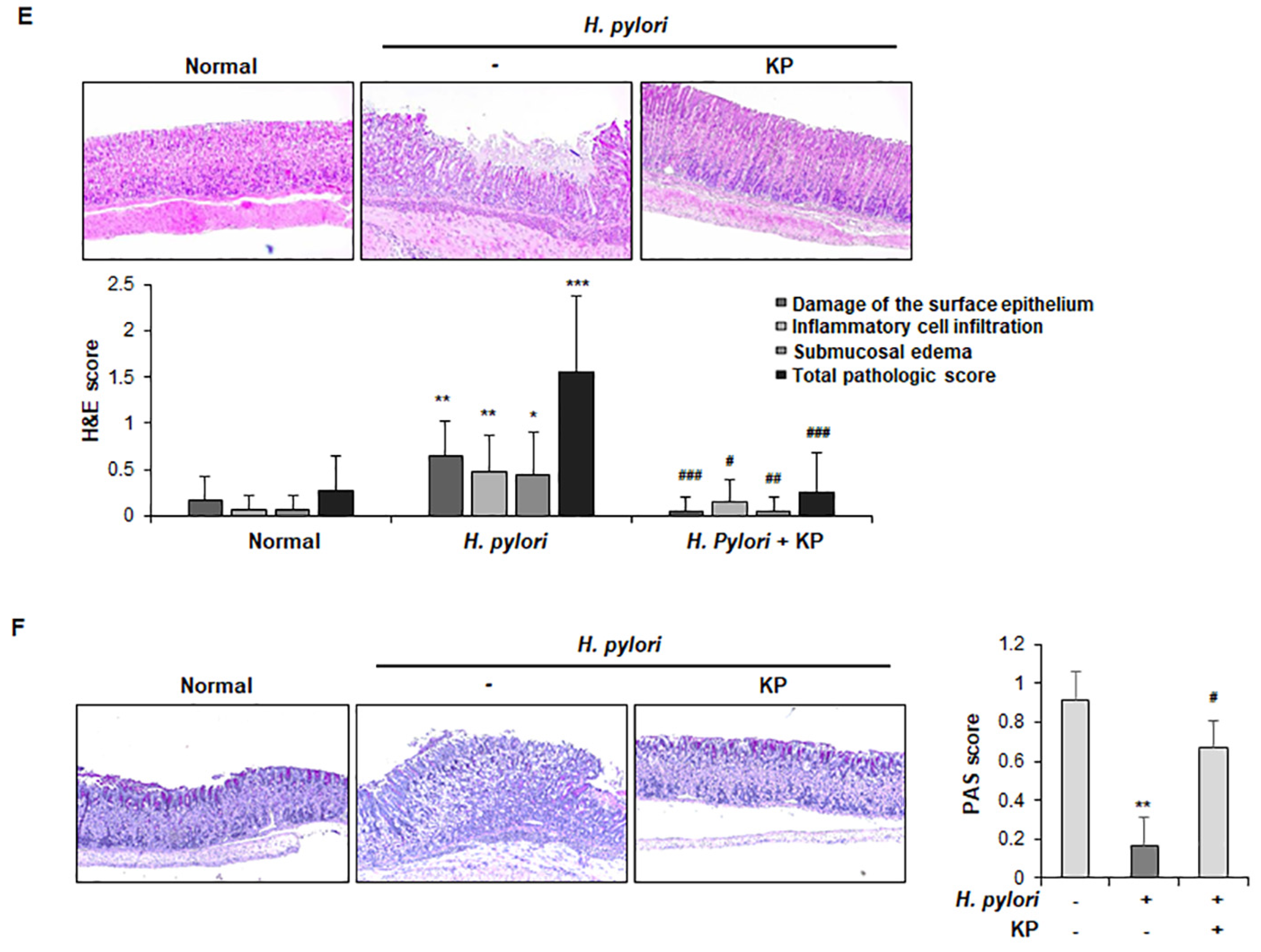
2.7. Histological Analysis
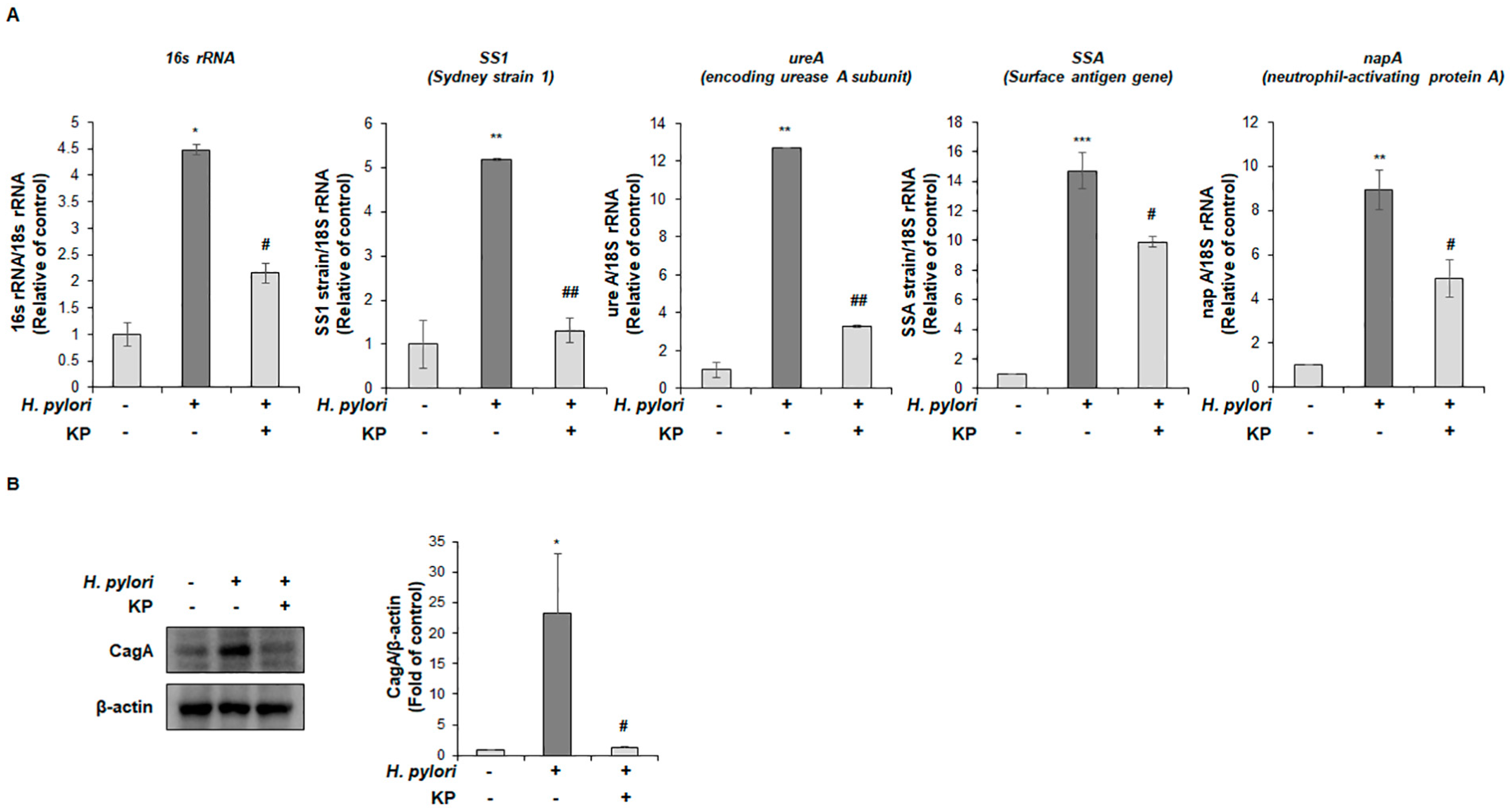
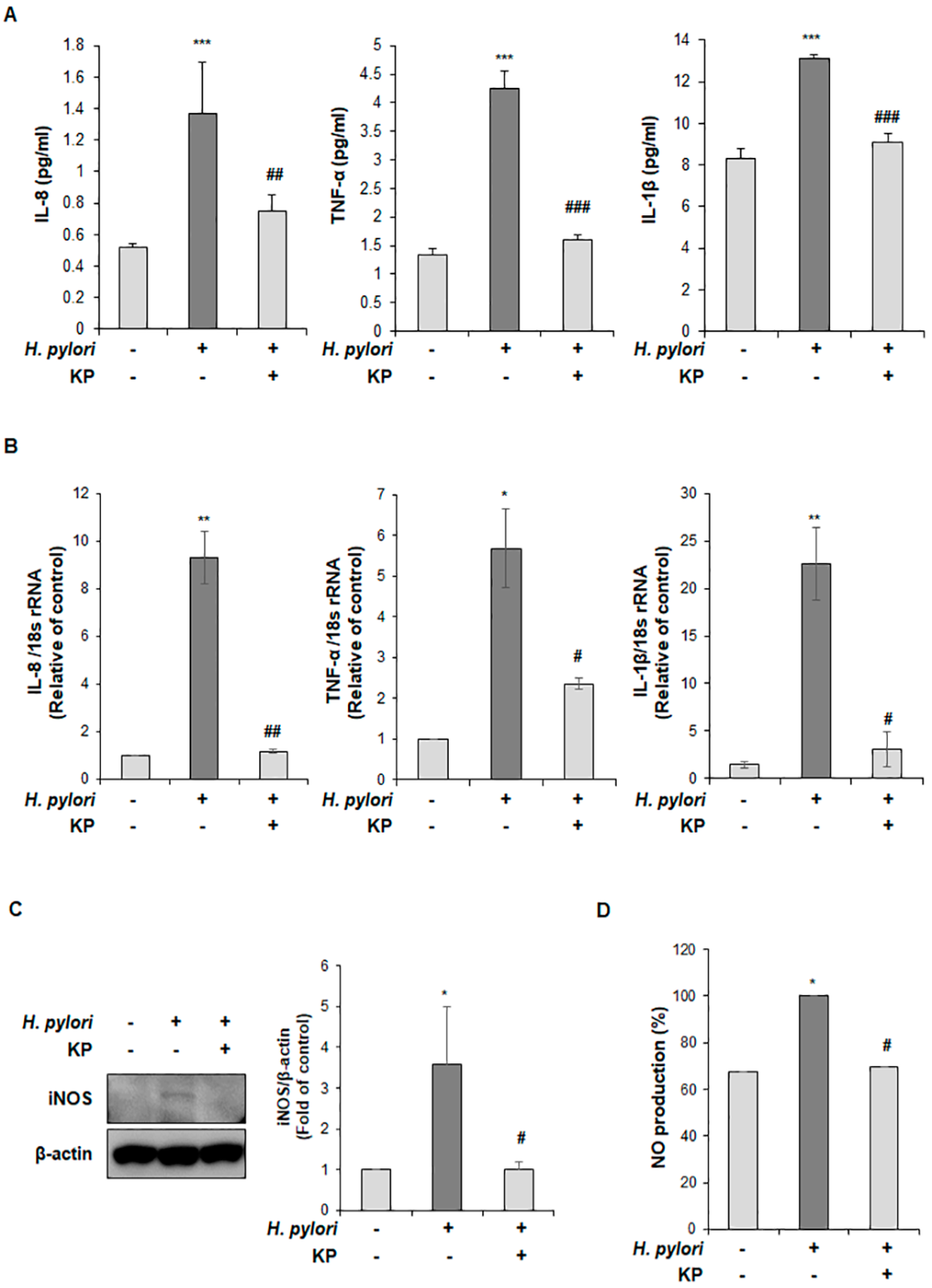
2.8. Quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR) Analysis
2.9. Western Blot Analysis
2.10. Cytokine Measurement via Enzyme-Linked Iimmunoassay (ELISA)
2.11. Measurement of Nitric Oxide (NO) Production
2.12. Statistical Analysis
3. Results
3.1. Gastric Mucosal Therapeutic Effect of KP in H. pylori-Infected Mice
3.2. KP Attenuates H. pylori-Related Virulence Factors in H. pylori-Infected Mice
3.3. KP Restrains the Pro-Inflammatory Response and NO Production in H. pylori-Infected Mice
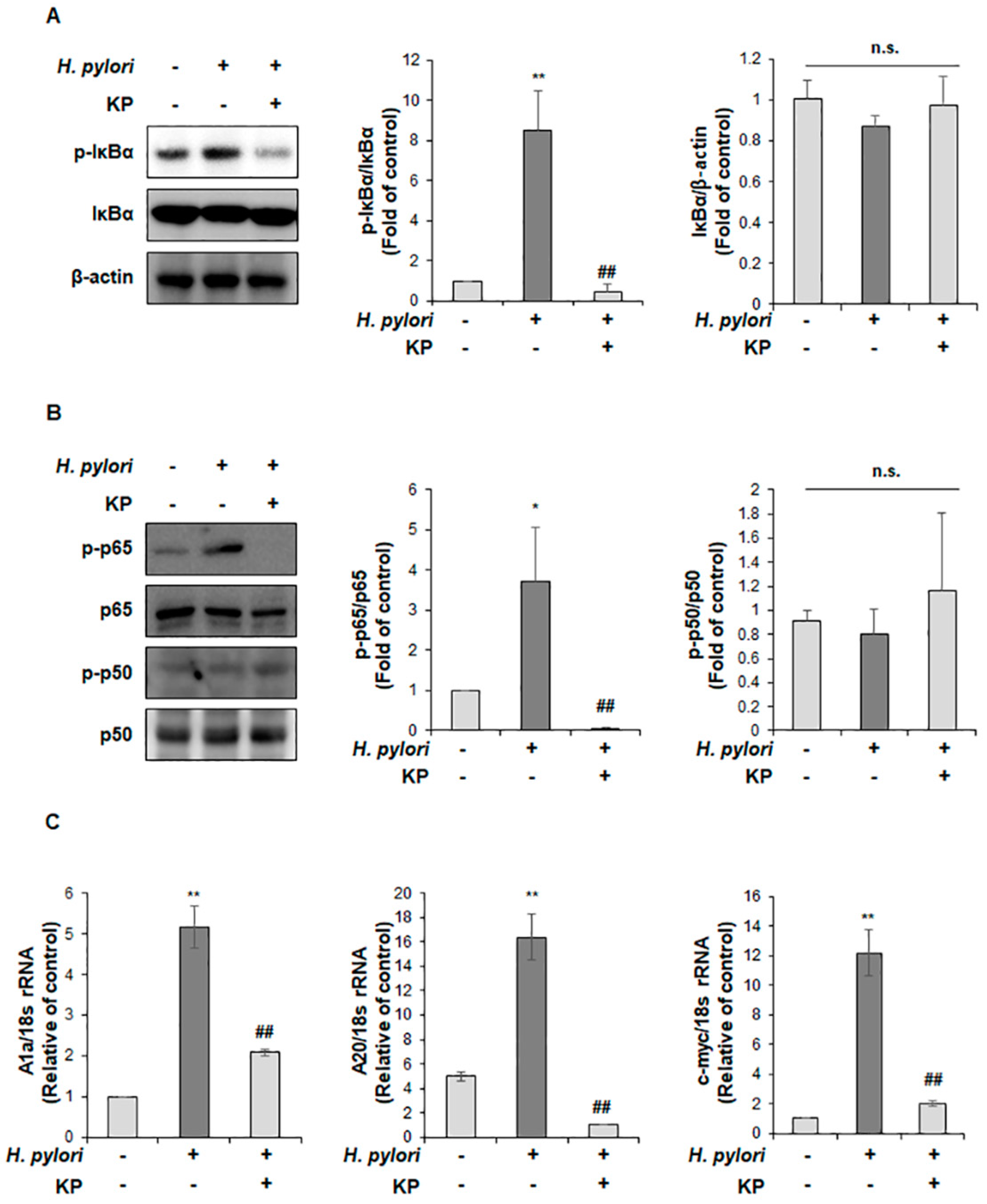
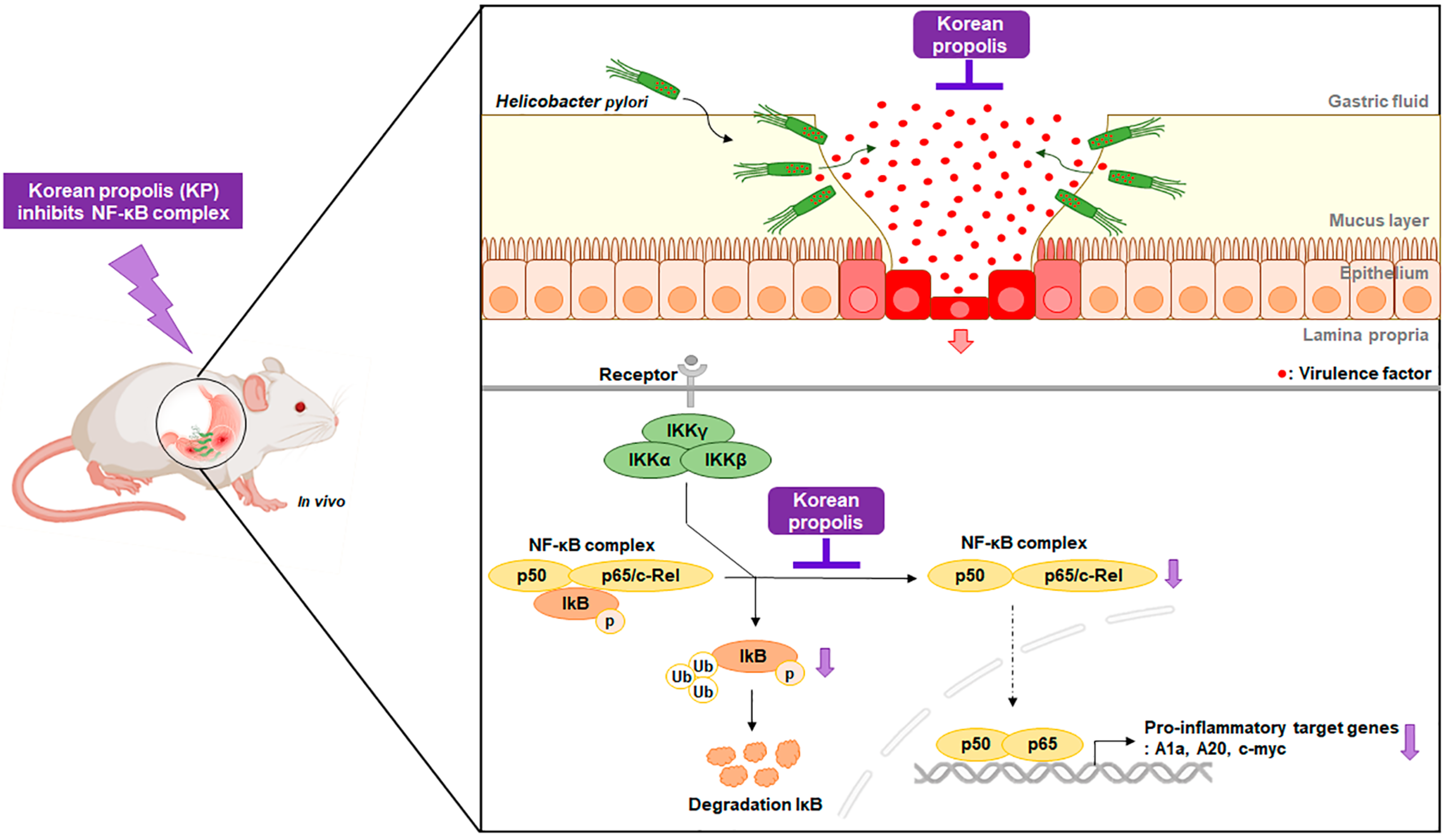
3.4. KP Regulates the NF-κB Signaling Pathway in H. pylori-Infected Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hooi, J.K.; Lai, W.Y.; Ng, W.K.; Suen, M.M.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.; Wu, J.C. Global prevalence of Helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjbar, R.; Behzadi, P.; Farshad, S. Advances in diagnosis and treatment of Helicobacter pylori infection. Acta Microbiol. Immunol. Hung. 2017, 64, 273–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamani, M.; Ebrahimtabar, F.; Zamani, V.; Miller, W.; Alizadeh-Navaei, R.; Shokri-Shirvani, J.; Derakhshan, M. Systematic review with meta-analysis: The worldwide prevalence of Helicobacter pylori infection. Aliment. Pharmacol. Ther. 2018, 47, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cover, T.L.; Blaser, M.J. Helicobacter pylori in health and disease. Gastroenterology 2009, 136, 1863–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testerman, T.L.; Morris, J. Beyond the stomach: An updated view of Helicobacter pylori pathogenesis, diagnosis, and treatment. World J. Gastroenterol. 2014, 20, 12781. [Google Scholar] [CrossRef] [PubMed]
- Marcus, E.A.; Sachs, G.; Scott, D.R. Eradication of Helicobacter pylori infection. Curr. Gastroenterol. Rep. 2016, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Suerbaum, S.; Michetti, P. Helicobacter pylori infection. N. Engl. J. Med. 2002, 347, 1175–1186. [Google Scholar] [CrossRef] [Green Version]
- Roesler, B.M.; Costa, S.C.; Zeitune, J.M. Eradication treatment of Helicobacter pylori infection: Its importance and possible relationship in preventing the development of gastric cancer. Int. Sch. Res. Notices 2012, 2012, 935410. [Google Scholar] [CrossRef] [Green Version]
- Sepulveda, A.R. Helicobacter, inflammation, and gastric cancer. Curr. Pathobiol. Rep. 2013, 1, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.A.; Tummuru, M.K.; Blaser, M.J.; Kerr, L.D. Activation of IL-8 gene expression by Helicobacter pylori is regulated by transcription factor nuclear factor-κB in gastric epithelial cells. J. Immunol. Res. 1998, 160, 2401–2407. [Google Scholar]
- Lamb, A.; Yang, X.D.; Tsang, Y.H.N.; Li, J.D.; Higashi, H.; Hatakeyama, M.; Peek, R.M.; Blanke, S.R.; Chen, L.F. Helicobacter pylori cagA activates NF-κB by targeting TAK1 for TRAF6-mediated Lys 63 ubiquitination. EMBO Rep. 2009, 10, 1242–1249. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Signaling to NF-κB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, B.B. Nuclear factor-κB: The enemy within. Cancer Cell 2004, 6, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Crabtree, J.E.; Lindley, I. Mucosal interleukin-8 and Helicobacter pylori-associated gastroduodenal disease. Eur. J. Gastroenterol. Hepatol. 1994, 6, S33–S38. [Google Scholar] [PubMed]
- Brandt, S.; Kwok, T.; Hartig, R.; König, W.; Backert, S. NF-κB activation and potentiation of proinflammatory responses by the Helicobacter pylori CagA protein. Proc. Natl. Acad. Sci. USA 2005, 102, 9300–9305. [Google Scholar] [CrossRef] [Green Version]
- Veenendaal, R.A.; Götz, J.M.; Lamers, C.B. Mucosal inflammation and disease in Helicobacter pylori infection. Scand. J. Gastroenterol. 1996, 218, 86–91. [Google Scholar] [CrossRef]
- Wang, Y.-C. Medicinal plant activity on Helicobacter pylori related diseases. World J. Gastroenterol. 2014, 20, 10368. [Google Scholar] [CrossRef]
- Islam, M.; Kusumoto, Y.; Al-Mamun, M.A. Cytotoxicity and cancer (HeLa) cell killing efficacy of aqueous garlic (Allium sativum) extract. J. Sci. Res. 2011, 3, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Kumar, M.; Saravanan, C.; Singh, S.K. Curcumin: A potential candidate for matrix metalloproteinase inhibitors. Expert Opin. Ther. Targets 2012, 16, 959–972. [Google Scholar] [CrossRef]
- Shapla, U.M.; Raihan, J.; Islam, A.; Alam, F.; Solayman, N.; Gan, S.H.; Hossen, S.; Khalil, I. Propolis: The future therapy against Helicobacter pylori-mediated gastrointestinal diseases. J. Appl. Biomed. 2018, 16, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Cardos, I.A.; Zaha, D.C.; Sindhu, R.K.; Cavalu, S. Revisiting therapeutic strategies for H. pylori treatment in the context of antibiotic resistance: Focus on alternative and complementary therapies. Molecules 2021, 26, 6078. [Google Scholar] [CrossRef] [PubMed]
- Salatino, A.; Teixeira, É.W.; Negri, G. Origin and chemical variation of Brazilian propolis. Evid.-Based Complement. Altern. Med. 2005, 2, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, A.; De, R.; Mukhopadhyay, A.K. Curcumin as a potential therapeutic candidate for Helicobacter pylori associated diseases. World J. Gastroenterol. 2016, 22, 2736. [Google Scholar] [CrossRef] [PubMed]
- Osés, S.M.; Pascual-Maté, A.; Fernández-Muiño, M.A.; López-Díaz, T.M.; Sancho, M.T. Bioactive properties of honey with propolis. Food Chem. 2016, 196, 1215–1223. [Google Scholar] [CrossRef]
- Pascual, C.; Gonzalez, R.; Torricella, R. Scavenging action of propolis extract against oxygen radicals. J. Ethnopharmacol. 1994, 41, 9–13. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, C.-P.; Wang, K.; Li, G.Q.; Hu, F.-L. Recent advances in the chemical composition of propolis. Molecules 2014, 19, 19610–19632. [Google Scholar] [CrossRef] [Green Version]
- Ahn, M.R.; Kumazawa, S.; Hamasaka, T.; Bang, K.S.; Nakayama, T. Antioxidant activity and constituents of propolis collected in various areas of Korea. J. Agric. Food Chem. 2004, 52, 7286–7292. [Google Scholar] [CrossRef]
- Kujumgiev, A.; Tsvetkova, I.; Serkedjieva, Y.; Bankova, V.; Christov, R.; Popov, S. Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. J. Ethnopharmacol. 1999, 64, 235–240. [Google Scholar] [CrossRef]
- Lotfy, M. Biological activity of bee propolis in health and disease. Asian Pac. J. Cancer Prev. 2006, 7, 22–31. [Google Scholar]
- Song, M.Y.; Lee, D.Y.; Kim, E.H. Anti-inflammatory and anti-oxidative effect of Korean propolis on Helicobacter pylori-induced gastric damage in vitro. J. Microbiol. 2020, 58, 878–885. [Google Scholar] [CrossRef]
- Han, S.M.; Hong, I.P.; Woo, S.O.; Kim, S.G.; Jang, H.R.; Jang, J.S. Anti-Helicobacter pylori activity of Korean propolis. Korean J. Food Nutr. 2016, 29, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Sipponen, P.; Price, A.B. The Sydney system for classification of gastritis 20 years ago. J. Gastroenterol. Hepatol. 2011, 26, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-Y.; Lee, D.-Y.; Yun, S.-M.; Kim, E.-H. GLUT3 promotes epithelial–mesenchymal transition via TGF-β/JNK/ATF2 signaling pathway in colorectal cancer cells. Biomedicines 2022, 10, 1837. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lu, A.P.; Zhang, L.; Li, Y.D. Anti-Helicobacter pylori immunoglobulin G (IgG) and IgA antibody responses and the value of clinical presentations in diagnosis of H. pylori infection in patients with precancerous lesions. World J. Gastroenterol. 2003, 9, 755–758. [Google Scholar] [CrossRef]
- Unver, S.; Kubilay, U.; Sezen, O.S.; Coskuner, T. Investigation of Helicobacter pylori colonization in adenotonsillectomy specimens by means of the CLO Test. Laryngoscope 2001, 111, 2183–2186. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.; Newton, J.; Oliver, L.; Jordan, N.; Strugala, V.; Pearson, J.P.; Dettmar, P.W. Mucus and H. pylori. J. Physiol. Pharmacol. 1997, 48, 297–305. [Google Scholar]
- Hidaka, E.; Ota, H.; Hidaka, H.; Hayama, M.; Matsuzawa, K.; Akamatsu, T.; Nakayama, J.; Katsuyama, T. Helicobacter pylori and two ultrastructurally distinct layers of gastric mucous cell mucins in the surface mucous gel layer. Gut 2001, 49, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.L.; Yeh, Y.C.; Sheu, B.S. The impacts of H. pylori virulence factors on the development of gastroduodenal diseases. J. Biomed. Sci. 2018, 25, 68. [Google Scholar] [CrossRef] [Green Version]
- Dincă, A.L.; Meliț, L.E.; Mărginean, C.O. Old and new aspects of H. pylori-associated Inflammation and gastric cancer. Children 2022, 9, 1083. [Google Scholar] [CrossRef]
- Sharma, J.; Al-Omran, A.; Parvathy, S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Cho, K.; Lee, H.G.; Piao, J.Y.; Kim, S.J.; Na, H.K.; Surh, Y.J. Protective effects of Silibinin on Helicobacter pylori-induced gastritis: NF-κB and STAT3 as potential targets. J. Cancer Prev. 2021, 26, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.G.; Rossi, A.; Amici, C. NF-κB and virus infection: Who controls whom. EMBO Rep. 2003, 22, 2552–2560. [Google Scholar] [CrossRef] [PubMed]
- Catrysse, L.; Vereecke, L.; Beyaert, R.; van Loo, G. A20 in inflammation and autoimmunity. Trends Immunol. 2014, 35, 22–31. [Google Scholar] [CrossRef]
- Moser, B.; Hochreiter, B.; Basílio, J.; Gleitsmann, V.; Panhuber, A.; Pardo-Garcia, A.; Hoesel, B.; Salzmann, M.; Resch, U.; Noreen, M.; et al. The inflammatory kinase IKKα phosphorylates and stabilizes c-Myc and enhances its activity. Mol. Cancer 2021, 20, 16. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.M.; Yun, S.M.; Choi, Y.H.; Heo, J.; Kim, N.J.; Kim, S.H.; Kim, E.H. Xanthohumol prevents dextran sulfate sodium-induced colitis via inhibition of IKKβ/NF-κB signaling in mice. Oncotarget 2018, 9, 866–880. [Google Scholar] [CrossRef] [Green Version]
- Beales, I.L. Efficacy of Helicobacter pylori eradication therapies: A single centre observational study. BMC Gastroenterol. 2001, 1, 7. [Google Scholar] [CrossRef] [Green Version]
- Ford, A.C.; Yuan, Y.; Moayyedi, P. Helicobacter pylori eradication therapy to prevent gastric cancer: Systematic review and meta-analysis. Gut 2020, 69, 2113–2121. [Google Scholar] [CrossRef]
- Ghotaslou, R.; Leylabadlo, H.E.; Asl, Y.M. Prevalence of antibiotic resistance in Helicobacter pylori: A recent literature review. World J. Methodol. 2015, 5, 164. [Google Scholar] [CrossRef]
- Thung, I.; Aramin, H.; Vavinskaya, V.; Gupta, S.; Park, J.; Crowe, S.; Valasek, M. The global emergence of Helicobacter pylori antibiotic resistance. Aliment. Pharmacol. Ther. 2016, 43, 514–533. [Google Scholar] [CrossRef] [Green Version]
- Oršolić, N.; Landeka Jurčević, I.; Đikić, D.; Rogić, D.; Odeh, D.; Balta, V.; Perak Junaković, E.; Terzić, S.; Jutrić, D. Effect of Propolis on diet-Induced hyperlipidemia and atherogenic Indices in mice. Antioxidants 2019, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, Y.; Sakai, H.; Hirata, A.; Yanai, T. Brazilian green propolis suppresses acetaminophen-induced hepatocellular necrosis by modulating inflammation-related factors in rats. J. Toxicol. Pathol. 2018, 31, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Piñeros, A.R.; de Lima, M.H.F.; Rodrigues, T.; Gembre, A.F.; Bertolini, T.B.; Fonseca, M.D.; Berretta, A.A.; Ramalho, L.N.Z.; Cunha, F.Q.; Hori, J.I.; et al. Green propolis increases myeloid suppressor cells and CD4+Foxp3+ cells and reduces Th2 inflammation in the lungs after allergen exposure. J. Ethnopharmacol. 2020, 252, 112496. [Google Scholar] [CrossRef]
- Wang, K.; Jin, X.; Li, Q.; Sawaya, A.C.H.F.; Le Leu, R.K.; Conlon, M.A.; Wu, L.; Hu, F. Propolis from different geographic origins decreases intestinal inflammation and bacteroides spp. populations in a model of DSS-induced colitis. Mol. Nutr. Food Res. 2018, 62, 1800080. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Tezuka, Y.; Adnyana, I.K.; Ishii, E.; Midorikawa, K.; Matsushige, K.; Kadota, S. Hepatoprotective and anti-Helicobacter pylori activities of constituents from Brazilian propolis. Phytomedicine 2001, 8, 16–23. [Google Scholar] [CrossRef]
- Nostro, A.; Cellini, L.; Bartolomeo, S.D.; Cannatelli, M.A.; Campli, E.D.; Procopio, F.; Grande, R.; Marzio, L.; Alonzo, V. Effects of combining extracts (from propolis or Zingiber officinale) with clarithromycin on Helicobacter pylori. Phytother. Res. 2006, 20, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Baltas, N.; Karaoglu, S.A.; Tarakci, C.; Kolayli, S. Effect of propolis in gastric disorders: Inhibition studies on the growth of Helicobacter pylori and production of its urease. J. Enzyme Inhib. Med. Chem. 2016, 31, 46–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyanova, L.; Derejian, S.; Koumanova, R.; Katsarov, N.; Gergova, G.; Mitov, I.; Nikolov, R.; Krastev, Z. Inhibition of Helicobacter pylori growth in vitro by Bulgarian propolis: Preliminary report. J. Med. Microbiol. 2003, 52, 417–419. [Google Scholar] [CrossRef]
- Romero, M.; Freire, J.; Pastene, E.; García, A.; Aranda, M.; González, C. Propolis polyphenolic compounds affect the viability and structure of Helicobacter pylori in vitro. Rev. Bras. Farmacogn. 2019, 29, 325–332. [Google Scholar] [CrossRef]
- Roesler, B.M.; Rabelo-Gonçalves, E.M.; Zeitune, J.M. Virulence Factors of Helicobacter pylori: A Review. Clin. Med. Insights Gastroenterol. 2014, 7, 9–17. [Google Scholar] [CrossRef]
- Šterbenc, A.; Jarc, E.; Poljak, M.; Homan, M. Helicobacter pylori virulence genes. World J. Gastroenterol. 2019, 25, 4870–4884. [Google Scholar] [CrossRef]
- Lamb, A.; Chen, L.F. Role of the Helicobacter pylori-induced inflammatory response in the development of gastric cancer. J. Cell. Biochem. 2013, 114, 491–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejati, S.; Karkhah, A.; Darvish, H.; Validi, M.; Ebrahimpour, S.; Nouri, H.R. Influence of Helicobacter pylori virulence factors CagA and VacA on pathogenesis of gastrointestinal disorders. Microb. Pathog. 2018, 117, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, E.; Tomimori, K.; Takamatsu, R.; Ishikawa, C.; Kinjo, F.; Hirayama, T.; Fujita, J.; Mori, N. Helicobacter pylori vacA activates NF-κB in T Cells via the classical but not alternative pathway. Helicobacter 2009, 14, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Lacy, D.B.; Ohi, M.D. The Helicobacter pylori cag type IV secretion system. Trends Microbiol. 2020, 28, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Viala, J.; Chaput, C.; Boneca, I.G.; Cardona, A.; Girardin, S.E.; Moran, A.P.; Athman, R.; Mémet, S.; Huerre, M.R.; Coyle, A.J. Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat. Immunol. 2004, 5, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Waskito, L.A.; Salama, N.R.; Yamaoka, Y. Pathogenesis of Helicobacter pylori infection. Helicobacter 2018, 23, e12516. [Google Scholar] [CrossRef] [Green Version]
- McGee, D.J.; Mobley, H.L. Pathogenesis of Helicobacter pylori infection. Curr. Opin. Gastroenterol. 2000, 16, 24–31. [Google Scholar] [CrossRef]
- Pignatelli, B.; Bancel, B.; Esteve, J.; Malaveille, C.; Calmels, S.; Correa, P.; Patricot, L.; Laval, M.; Lyandrat, N.; Ohshima, H. Inducible nitric oxide synthase, anti-oxidant enzymes and Helicobacter pylori infection in gastritis and gastric precancerous lesions in humans. Eur. J. Cancer Prev. 1998, 7, 439–447. [Google Scholar] [CrossRef]
- Bothmer, C.v.; Edebo, A.; Lönroth, H.; Olbe, L.; Pettersson, A.; Fändriks, L. Helicobacter pylori infection inhibits antral mucosal nitric oxide production in humans. Scand. J. Gastroenterol. 2002, 37, 404–408. [Google Scholar] [CrossRef]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef]
- Fu, S.; Ramanujam, K.S.; Wong, A.; Fantry, G.T.; Drachenberg, C.B.; James, S.P.; Meltzer, S.J.; Wilson, K.T. Increased expression and cellular localization of inducible nitric oxide synthase and cyclooxygenase 2 in Helicobacter pylori gastritis. Gastroenterology 1999, 116, 1319–1329. [Google Scholar] [CrossRef]
- Saaed, H.K.; Chiggiato, L.; Webb, D.L.; Rehnberg, A.S.; Rubio, C.A.; Befrits, R.; Hellström, P.M. Elevated gaseous luminal nitric oxide and circulating IL-8 as features of Helicobacter pylori-induced gastric inflammation. UPS J. Med. Sci. 2021, 126, e8116. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.-T.; Oh, S.-Y.; Ahn, B.-W.; Kim, Y.; Jang, D.; Yang, K.; Hahm, K.; Kim, D. Decreased Helicobacter pylori associated gastric carcinogenesis in mice lacking inducible nitric oxide synthase. Gut 2004, 53, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Lamb, A.; Chen, L.-F. The many roads traveled by Helicobacter pylori to NF-κB activation. Gut Microbes 2010, 1, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Saha, K.; Sarkar, D.; Khan, U.; Karmakar, B.C.; Paul, S.; Mukhopadhyay, A.K.; Dutta, S.; Bhattacharya, S. Capsaicin inhibits inflammation and gastric damage during H pylori infection by targeting NF-kB–miRNA axis. Pathogens 2022, 11, 641. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identification | Gene | Primer Sequence (5′ to 3′) | |
|---|---|---|---|
| Specific for H. pylori | 16S rRNA | Forward | CTC ATT GCG AAG GCG ACC T |
| Reverse | TCT AAT CCT GTT TGC TCC CCA | ||
| SS1 | Forward | CTT AAC CAT AGA ACT GCA TTT GAA ACT AC | |
| Reverse | GGT CGC CTT CGC AAT GAG TA | ||
| ureA | Forward | AGG AAA CAT CGC TTC AAT ACC | |
| Reverse | AGG AAA CAT CGC TTC AAT ACC | ||
| SSA | Forward | TGG CGT GTC TAT TGA CAG CGA GC | |
| Reverse | CCT GCT GGG CAT ACT TCA CCA TG | ||
| napA | Forward | TCC TTT CAG CGA GAT CGT CA | |
| Reverse | GAA TGT GAA AGG CAC CGA TT | ||
| Specific for inflammation | IL-8 | Forward | TCC TTG TTC CAC TGT GCC TTG |
| Reverse | TGC TTC CAC ATG TCC TCA CAA | ||
| TNF-α | Forward | TCA GAG GGC CTG TAC CTC AT | |
| Reverse | GGA AGA CCC CTC CCA GAT AG | ||
| IL-1β | Forward | TTA AAG CCC GCC TGA CAG A | |
| Reverse | GCG AAT GAC AGA GGG TTT CTT | ||
| A1a | Forward | TCC ACA AGA GCA GAT TGC CCT G | |
| Reverse | GCC AGC CAG ATT TGG GTT CAA AC | ||
| A20 | Forward | AGC AAG TGC AGG AAA GCT GGC T | |
| Reverse | GCT TTC GCA GAG GCA GTA ACA G | ||
| c-myc | Forward | GCT GTT TGA AGG CTG GAT TTC | |
| Reverse | GAT GAA ATA GGG CTG TAC GGA G | ||
| Internal control | 18s rRNA | Forward | GCA ATT ATT CCC CAT GAA CG |
| Reverse | GGC CTC ACT AAA CCA TCC AA | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, M.-Y.; Lee, D.-Y.; Han, Y.-M.; Kim, E.-H. Anti-Inflammatory Effect of Korean Propolis on Helicobacter pylori-Infected Gastric Mucosal Injury Mice Model. Nutrients 2022, 14, 4644. https://doi.org/10.3390/nu14214644
Song M-Y, Lee D-Y, Han Y-M, Kim E-H. Anti-Inflammatory Effect of Korean Propolis on Helicobacter pylori-Infected Gastric Mucosal Injury Mice Model. Nutrients. 2022; 14(21):4644. https://doi.org/10.3390/nu14214644
Chicago/Turabian StyleSong, Moon-Young, Da-Young Lee, Young-Min Han, and Eun-Hee Kim. 2022. "Anti-Inflammatory Effect of Korean Propolis on Helicobacter pylori-Infected Gastric Mucosal Injury Mice Model" Nutrients 14, no. 21: 4644. https://doi.org/10.3390/nu14214644
APA StyleSong, M.-Y., Lee, D.-Y., Han, Y.-M., & Kim, E.-H. (2022). Anti-Inflammatory Effect of Korean Propolis on Helicobacter pylori-Infected Gastric Mucosal Injury Mice Model. Nutrients, 14(21), 4644. https://doi.org/10.3390/nu14214644