
NAPRT, but Not NAMPT, Provides Additional Support for NAD Synthesis in Esophageal Precancerous Lesions

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Collection
2.2. Laboratory Measurements
2.3. Statistical Analysis
3. Results
3.1. General Characteristics of the Subjects
3.2. Relevant Variables for NAD of the Subjects
3.3. Association between Relevant Variables for NAD and Risk of EPL
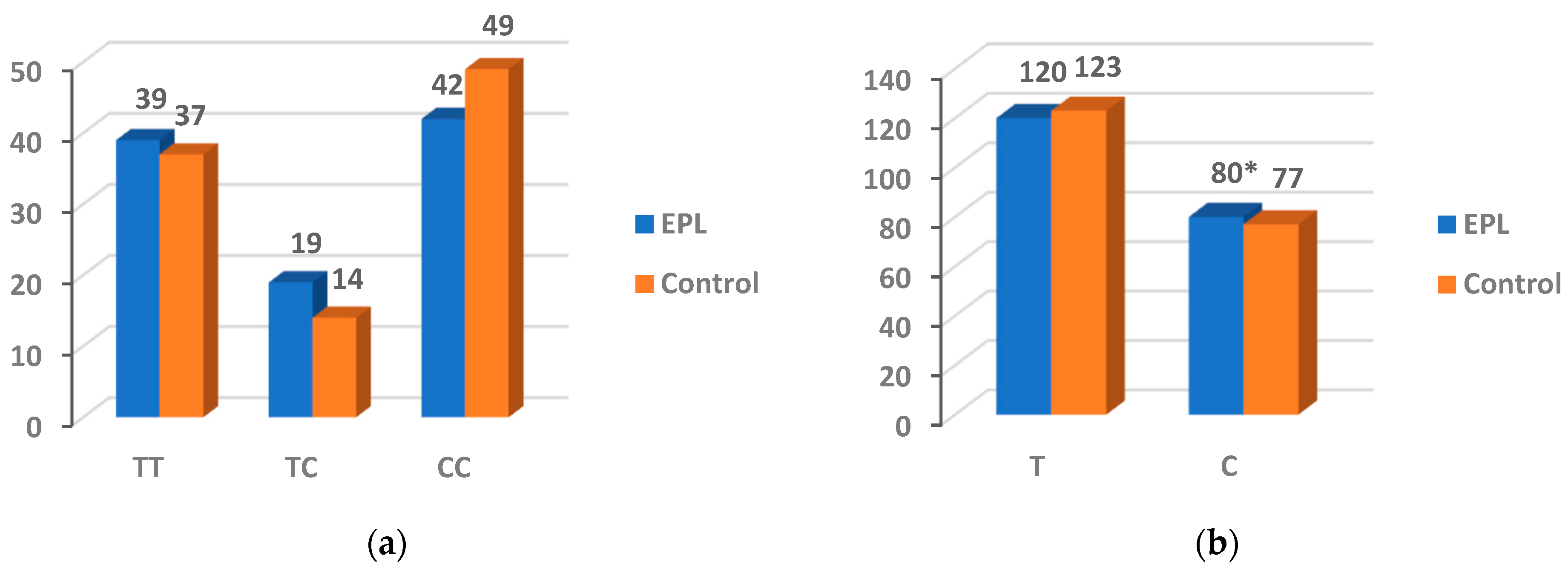
3.4. The Gene Polymorphism of PARP-1 V762A
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morgan, E.; Soerjomataram, I.; Rumgay, H.; Coleman, H.G.; Thrift, A.P.; Vignat, J.; Laversanne, M.; Ferlay, J.; Arnold, M. The Global Landscape of Esophageal Squamous Cell Carcinoma and Esophageal Adenocarcinoma Incidence and Mortality in 2020 and Projections to 2040: New Estimates From GLOBOCAN 2020. Gastroenterology 2022, 163, 649–658.e642. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.W.; Zhao, Z.H.; Wang, L.; Li, P.; Chen, K.S.; Zhang, J.Y.; Li, W.C.; Jiang, G.Z.; Li, X.N. MicroRNA-134 prevents the progression of esophageal squamous cell carcinoma via the PLXNA1-mediated MAPK signalling pathway. EBioMedicine 2019, 46, 66–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Zhu, Y.; Ding, Y.; Niu, T.; Zhang, Y.; Wu, H.; Zhu, L.; Yuan, B.; Qiao, Y.; Lu, J.; et al. DNA polymerase β deficiency promotes the occurrence of esophageal precancerous lesions in mice. Neoplasia 2021, 23, 663–675. [Google Scholar] [CrossRef]
- Cantó, C.; Menzies, K.J.; Auwerx, J. NAD(+) Metabolism and the Control of Energy Homeostasis: A Balancing Act between Mitochondria and the Nucleus. Cell Metab. 2015, 22, 31–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanem, M.S.; Monacelli, F.; Nencioni, A. Advances in NAD-Lowering Agents for Cancer Treatment. Nutrients 2021, 13, 1665. [Google Scholar] [CrossRef] [PubMed]
- Preiss, J.; Handler, P. Biosynthesis of diphosphopyridine nucleotide. I. Identification of intermediates. J. Biol. Chem. 1958, 233, 488–492. [Google Scholar] [CrossRef]
- Preiss, J.; Handler, P. Biosynthesis of diphosphopyridine nucleotide. II. Enzymatic aspects. J. Biol. Chem. 1958, 233, 493–500. [Google Scholar] [CrossRef]
- Preiss, J.; Handler, P. Enzymatic synthesis of nicotinamide mononucleotide. J. Biol. Chem. 1957, 225, 759–770. [Google Scholar] [CrossRef]
- Sharif, T.; Martell, E.; Dai, C.; Ghassemi-Rad, M.S.; Kennedy, B.E.; Lee, P.W.K.; Gujar, S. Regulation of Cancer and Cancer-Related Genes via NAD. Antioxid. Redox Signal. 2019, 30, 906–923. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, H. Reprogramming of glucose, fatty acid and amino acid metabolism for cancer progression. Cell. Mol. Life Sci. CMLS 2016, 73, 377–392. [Google Scholar] [CrossRef]
- Piacente, F.; Caffa, I.; Ravera, S.; Sociali, G.; Passalacqua, M.; Vellone, V.G.; Becherini, P.; Reverberi, D.; Monacelli, F.; Ballestrero, A.; et al. Nicotinic Acid Phosphoribosyltransferase Regulates Cancer Cell Metabolism, Susceptibility to NAMPT Inhibitors, and DNA Repair. Cancer Res. 2017, 77, 3857–3869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhry, S.; Zanca, C.; Rajkumar, U.; Koga, T.; Diao, Y.; Raviram, R.; Liu, F.; Turner, K.; Yang, H.; Brunk, E.; et al. NAD metabolic dependency in cancer is shaped by gene amplification and enhancer remodelling. Nature 2019, 569, 570–575. [Google Scholar] [CrossRef]
- Takahashi, S.; Miura, N.; Harada, T.; Wang, Z.; Wang, X.; Tsubokura, H.; Oshima, Y.; Hasegawa, J.; Inagaki, Y.; Shiota, G. Prognostic impact of clinical course-specific mRNA expression profiles in the serum of perioperative patients with esophageal cancer in the ICU: A case control study. J. Transl. Med. 2010, 8, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kim, H.; Lee, J.E.; Shin, S.J.; Oh, S.; Kwon, G.; Kim, H.; Choi, Y.Y.; White, M.A.; Paik, S.; et al. Selective Cytotoxicity of the NAMPT Inhibitor FK866 Toward Gastric Cancer Cells With Markers of the Epithelial-Mesenchymal Transition, Due to Loss of NAPRT. Gastroenterology 2018, 155, 799–814.e713. [Google Scholar] [CrossRef]
- Nacarelli, T.; Fukumoto, T.; Zundell, J.A.; Fatkhutdinov, N.; Jean, S.; Cadungog, M.G.; Borowsky, M.E.; Zhang, R. NAMPT Inhibition Suppresses Cancer Stem-like Cells Associated with Therapy-Induced Senescence in Ovarian Cancer. Cancer Res. 2020, 80, 890–900. [Google Scholar] [CrossRef] [Green Version]
- Hasmann, M.; Schemainda, I. FK866, a highly specific noncompetitive inhibitor of nicotinamide phosphoribosyltransferase, represents a novel mechanism for induction of tumor cell apoptosis. Cancer Res. 2003, 63, 7436–7442. [Google Scholar] [PubMed]
- Olesen, U.H.; Christensen, M.K.; Björkling, F.; Jäättelä, M.; Jensen, P.B.; Sehested, M.; Nielsen, S.J. Anticancer agent CHS-828 inhibits cellular synthesis of NAD. Biochem. Biophys. Res. Commun. 2008, 367, 799–804. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Cantó, C.; Wanders, R.J.; Auwerx, J. The secret life of NAD+: An old metabolite controlling new metabolic signaling pathways. Endocr. Rev. 2010, 31, 194–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarugi, A.; Dölle, C.; Felici, R.; Ziegler, M. The NAD metabolome—A key determinant of cancer cell biology. Nat. Rev. Cancer 2012, 12, 741–752. [Google Scholar] [CrossRef]
- Malanga, M.; Althaus, F.R. The role of poly(ADP-ribose) in the DNA damage signaling network. Biochem. Cell Biol. 2005, 83, 354–364. [Google Scholar] [CrossRef]
- Yu, H.; Ma, H.; Yin, M.; Wei, Q. Association between PARP-1 V762A polymorphism and cancer susceptibility: A meta-analysis. Genet. Epidemiol. 2012, 36, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, H.; Jo, Y.; Yoon, M.; Hong, S. Thymidine decreases the DNA damage and apoptosis caused by tumor-treating fields in cancer cell lines. Genes Genom. 2021, 43, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Ying, S.; Zhang, C.; Lin, R.; Zheng, J.; Zhang, G.; Tian, D.; Guo, Y.; Du, C.; et al. Genetic Alterations in Esophageal Tissues From Squamous Dysplasia to Carcinoma. Gastroenterology 2017, 153, 166–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Li, Y.; Li, X.; Zhou, W.; Shi, B.; Chen, H.; Yuan, W. PARP-1 Val762Ala polymorphism, CagA+ H. pylori infection and risk for gastric cancer in Han Chinese population. Mol. Biol. Rep. 2009, 36, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zha, Y.; Du, F.; Liu, J.; Li, X.; Zhao, X. Contributions of PARP-1 rs1136410 C>T polymorphism to the development of cancer. J. Cell. Mol. Med. 2020, 24, 14639–14644. [Google Scholar] [CrossRef]
- Qin, Q.; Lu, J.; Zhu, H.; Xu, L.; Cheng, H.; Zhan, L.; Yang, X.; Zhang, C.; Sun, X. PARP-1 Val762Ala polymorphism and risk of cancer: A meta-analysis based on 39 case-control studies. PLoS ONE 2014, 9, e98022. [Google Scholar] [CrossRef]
- Pan, D.; Su, M.; Zhang, T.; Miao, C.; Fu, L.; Yang, L.; Song, G.; Raine, P.J.; Wang, S.; Sun, G. A Distinct Epidemiologic Pattern of Precancerous Lesions of Esophageal Squamous Cell Carcinoma in a High-risk Area of Huai’an, Jiangsu Province, China. Cancer Prev. Res. 2019, 12, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Wang, S. Vitamin B(12) may play a preventive role in esophageal precancerous lesions: A case-control study based on markers in blood and 3-day duplicate diet samples. Eur. J. Nutr. 2021, 60, 3375–3386. [Google Scholar] [CrossRef] [PubMed]
- Shats, I.; Williams, J.G.; Liu, J.; Makarov, M.V.; Wu, X.; Lih, F.B.; Deterding, L.J.; Lim, C.; Xu, X.; Randall, T.A.; et al. Bacteria Boost Mammalian Host NAD Metabolism by Engaging the Deamidated Biosynthesis Pathway. Cell Metab. 2020, 31, 564–579.e567. [Google Scholar] [CrossRef]
- Tempel, W.; Rabeh, W.M.; Bogan, K.L.; Belenky, P.; Wojcik, M.; Seidle, H.F.; Nedyalkova, L.; Yang, T.; Sauve, A.A.; Park, H.W.; et al. Nicotinamide riboside kinase structures reveal new pathways to NAD+. PLoS Biol. 2007, 5, e263. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Su, X.; Quinn, W.J., 3rd; Hui, S.; Krukenberg, K.; Frederick, D.W.; Redpath, P.; Zhan, L.; Chellappa, K.; White, E.; et al. Quantitative Analysis of NAD Synthesis-Breakdown Fluxes. Cell Metab. 2018, 27, 1067–1080.e1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, I.; Hale, M.; Demetriou, A.; Mohamed, A.E. Pathological effects of pellagra on the esophagus. Nutr. Cancer 1990, 14, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, S.; Bidoli, E.; Negri, E.; Zambon, P.; Talamini, R.; Ruol, A.; Parpinel, M.; Levi, F.; Simonato, L.; La Vecchia, C. Role of macronutrients, vitamins and minerals in the aetiology of squamous-cell carcinoma of the oesophagus. Int. J. Cancer 2000, 86, 626–631. [Google Scholar] [CrossRef]
- Jessri, M.; Rashidkhani, B.; Hajizadeh, B.; Jessri, M.; Gotay, C. Macronutrients, vitamins and minerals intake and risk of esophageal squamous cell carcinoma: A case-control study in Iran. Nutr. J. 2011, 10, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hormozdiari, H.; Day, N.E.; Aramesh, B.; Mahboubi, E. Dietary factors and esophageal cancer in the Caspian Littoral of Iran. Cancer Res. 1975, 35, 3493–3498. [Google Scholar]
- Piacente, F.; Caffa, I.; Nencioni, A. Nicotinic acid: A case for a vitamin that moonlights for cancer? Cell Cycle 2017, 16, 1635–1636. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.F.; Henderson, L.M. Pyridinium precursors of pyridine nucleotides in perfused rat kidney and in the testis. J. Biol. Chem. 1972, 247, 8023–8030. [Google Scholar] [CrossRef]
- Hara, N.; Yamada, K.; Shibata, T.; Osago, H.; Hashimoto, T.; Tsuchiya, M. Elevation of cellular NAD levels by nicotinic acid and involvement of nicotinic acid phosphoribosyltransferase in human cells. J. Biol. Chem. 2007, 282, 24574–24582. [Google Scholar] [CrossRef] [Green Version]
- Moffett, J.R.; Arun, P.; Puthillathu, N.; Vengilote, R.; Ives, J.A.; Badawy, A.A.; Namboodiri, A.M. Quinolinate as a Marker for Kynurenine Metabolite Formation and the Unresolved Question of NAD(+) Synthesis During Inflammation and Infection. Front. Immunol. 2020, 11, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelius, M.E.; Wang, T.W.; Jamal, A.; Loretan, C.G.; Neff, L.J. Tobacco Product Use Among Adults—United States, 2019. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 1736–1742. [Google Scholar] [CrossRef]
- Bobrova, N.; West, R.; Malyutina, D.; Malyutina, S.; Bobak, M. Gender differences in drinking practices in middle aged and older Russians. Alcohol Alcohol. 2010, 45, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.; Lim, S.; Lee, S. Why is high-risk drinking more prevalent among men than women? Evidence from South Korea. BMC Public Health 2012, 12, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lande, R.G.; Marin, B.A.; Chang, A.S.; Lande, G.R. Gender differences and alcohol use in the US Army. J. Am. Osteopath. Assoc. 2007, 107, 401–407. [Google Scholar]
- Brennan, P.L.; Schutte, K.K.; Moos, B.S.; Moos, R.H. Twenty-year alcohol-consumption and drinking-problem trajectories of older men and women. J. Stud. Alcohol Drugs 2011, 72, 308–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukocheva, O.A.; Li, B.; Due, S.L.; Hussey, D.J.; Watson, D.I. Androgens and esophageal cancer: What do we know? World J. Gastroenterol. 2015, 21, 6146–6156. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K. Effect of sex hormone on the experimental induction of esophageal cancer. Nihon Geka Gakkai Zasshi 1985, 86, 280–289. [Google Scholar] [PubMed]
- Matsuoka, H.; Sugimachi, K.; Ueo, H.; Kuwano, H.; Nakano, S.; Nakayama, M. Sex hormone response of a newly established squamous cell line derived from clinical esophageal carcinoma. Cancer Res. 1987, 47, 4134–4140. [Google Scholar]
- Asanuma, K.; Iijima, K.; Shimosegawa, T. Gender difference in gastro-esophageal reflux diseases. World J. Gastroenterol. 2016, 22, 1800–1810. [Google Scholar] [CrossRef]
- Gaut, Z.N.; Solomon, H.M. Inhibition of nicotinate phosphoribosyltransferase in human platelet lysate by nicotinic acid analogs. Biochem. Pharmacol. 1971, 20, 2903–2906. [Google Scholar] [CrossRef]
- Franco, J.; Piacente, F. Structure-Based Identification and Biological Characterization of New NAPRT Inhibitors. Pharmaceuticals 2022, 15, 855. [Google Scholar] [CrossRef]
- Ghanem, M.S.; Caffa, I. Identification of NAPRT Inhibitors with Anti-Cancer Properties by In Silico Drug Discovery. Pharmaceuticals 2022, 15, 848. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Wang, H.; Zhou, K.; Li, Y.; Chen, X.; Zhou, G.; Zhu, Y.; Miao, X.; Tan, W.; Wei, Q.; et al. Identification of genetic variants in base excision repair pathway and their associations with risk of esophageal squamous cell carcinoma. Cancer Res. 2004, 64, 4378–4384. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Median (25th–75th) | EPL | Controls | p Value |
|---|---|---|---|
| Niacin (nmol/L) | 34.209 (30.238, 38.756) | 32.488 (29.245, 41.422) | 0.951 |
| Nicotinamide (ng/mL) | 0.613 (0.351, 1.042) | 0.586 (0.371, 1.040) | 0.732 |
| NAPRT (pg/mL) | 194.117 (152.068, 237.732) | 146.972 (115.463, 219.051) | <0.001 * |
| NAMPT (pg/mL) | 164.658 (142.565, 188.002) | 160.528 (137.922, 190.628) | 0.767 |
| PARP-1 (pg/mL) | 879.598 (669.161, 1294.424) | 709.649 (431.848, 979.108) | 0.001 * |
| Q1 | Q2 | Q3 | Q4 | p for Trend | |
|---|---|---|---|---|---|
| OR (95%CI) | OR (95%CI) | OR (95%CI) | OR (95%CI) | ||
| Niacin (nmol/L) | 21.335–29.522 | 29.522–33.858 | 33.858–40.162 | 40.162–395.435 | |
| EPL cases (%) | 46.94 | 42.86 | 69.39 | 40.82 | |
| crude model | 1 | 0.913 (0.505–1.650) | 1.478 (0.871–2.509) | 0.870 (0.478–1.583) | 0.642 |
| adjusted model | 1 | 0.970 (0.534–1.763) | 1.341 (0.780–2.304) | 0.886 (0.484–1.620) | 0.632 |
| Nicotinamide (ng/mL) | 0.070–0.362 | 0.362–0.591 | 0.591–1.039 | 1.039–37.700 | |
| EPL cases (%) | 52.00 | 46.00 | 52.00 | 50.00 | |
| crude model | 1 | 0.885 (0.505–1.550) | 1.000 (0.581–1.722) | 0.962 (0.555–1.665) | 0.999 |
| adjusted model | 1 | 1.063 (0.597–1.894) | 1.071 (0.615–1.866) | 1.090 (0.615–1.931) | 0.804 |
| NAMPT (pg/mL) | 65.119–141.113 | 141.113–161.586 | 161.586–188.603 | 188.603–776.568 | |
| EPL cases (%) | 42.86 | 52.08 | 57.14 | 47.92 | |
| crude model | 1 | 1.215 (0.680–2.171) | 1.333 (0.757–2.348) | 1.118 (0.619–2.020) | 0.819 |
| adjusted model | 1 | 1.221 (0.680–2.190) | 1.347 (0.760–2.387) | 1.192 (0.655–2.170) | 0.641 |
| NAPRT (pg/mL) | 44.12–131.20 | 131.21–171.67 | 171.68–233.12 | 233.132065.36 | |
| EPL cases (%) | 20.41 | 54.16 | 73.47 | 52.08 | |
| crude model | 1 | 2.654 (1.280–5.504) * | 3.600 (1.787–7.254) * | 2.552 (1.226–5.314) * | 0.117 |
| adjusted model | 1 | 2.526 (1.208–5.282) * | 3.268 (1.597–6.685) * | 2.477 (1.187–5.170) * | 0.132 |
| PARP-1 (pg/mL) | 133.53–537.78 | 537.79–790.48 | 790.49–1070.71 | 1070.71–3099.21 | |
| EPL cases (%) | 37.50 | 40.43 | 56.25 | 65.96 | |
| crude model | 1 | 1.078 (0.566–2.054) | 1.500 (0.826–2.723) | 1.759 (0.984–3.144) | 0.033 |
| adjusted model | 1 | 0.987 (0.510–1.908) | 1.376 (0.753–2.517) | 1.816 (1.011–3.262) * | 0.021 * |
| Q1 | Q2 | Q3 | Q4 | p for Trend | |
|---|---|---|---|---|---|
| OR (95%CI) | OR (95%CI) | OR (95%CI) | OR (95%CI) | ||
| Male | |||||
| Niacin (nmol/L) | |||||
| EPL cases (%) | 54.55 | 38.46 | 72.00 | 37.93 | |
| Adjusted OR (95%CI) | 1 | 0.728 (0.306–2.610) | 1.229 (0.579–2.610) | 0.703 (0.301–1.645) | 0.483 |
| Nicotinamide (ng/mL) | |||||
| EPL cases (%) | 50.00 | 44.44 | 44.44 | 58.62 | |
| Adjusted OR (95%CI) | 1 | 0.927 (0.387–2.220) | 0.856 (0.391–1.877) | 1.251 (0.614–2.549) | 0.454 |
| NAMPT (pg/mL) | |||||
| EPL cases (%) | 48.00 | 50.00 | 53.85 | 44.44 | |
| Adjusted OR (95%CI) | 1 | 1.165 (0.513–2.648) | 1.106 (0.499–2.451) | 1.019 (0.444–2.339) | 0.949 |
| NAPRT (pg/mL) | |||||
| EPL cases (%) | 21.74 | 48.15 | 81.48 | 43.48 | |
| Adjusted OR (95%CI) | 1 | 2.373 (0.820–0.869) | 3.464 (1.287–9.319) * | 2.143 (0.723–6.350) | 0.401 |
| PARP-1 (pg/mL) | |||||
| EPL cases (%) | 31.25 | 37.04 | 76.92 | 67.65 | |
| Adjusted OR (95%CI) | 1 | 1.355 (0.452–4.056) | 1.709 (0.600–4.866) | 2.619 (0.972–7.062) | 0.020 * |
| Female | |||||
| Niacin (nmol/L) | |||||
| EPL cases (%) | 40.74 | 47.83 | 66.67 | 45.00 | |
| Adjusted OR (95%CI) | 1 | 1.237 (0.515–2.973) | 1.411 (0.614–3.240) | 1.196 (0.473–3.024) | 0.815 |
| Nicotinamide (ng/mL) | |||||
| EPL cases (%) | 55.00 | 46.88 | 60.87 | 38.10 | |
| Adjusted OR (95%CI) | 1 | 1.304 (0.496–3.432) | 1.503 (0.564–4.010) | 1.025 (0.357–2.942) | 0.783 |
| NAMPT (pg/mL) | |||||
| EPL cases (%) | 37.50 | 50.00 | 60.87 | 52.38 | |
| Adjusted OR (95%CI) | 1 | 1.233 (0.525–2.896) | 1.507 (0.630–3.607) | 1.312 (0.528–3.260) | 0.575 |
| NAPRT (pg/mL) | |||||
| EPL cases (%) | 19.23 | 61.90 | 63.64 | 60.00 | |
| Adjusted OR (95%CI) | 1 | 2.373 (0.820–6.869) | 3.464 (1.287–9.319) * | 2.143 (0.723–6.350) | 0.261 |
| PARP-1 (pg/mL) | |||||
| EPL cases (%) | 40.63 | 45.00 | 60.87 | 61.54 | |
| Adjusted OR (95%CI) | 1 | 0.880 (0.352–2.203) | 1.254 (0.572–2.750) | 1.576 (0.646–3.848) | 0.278 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Pan, D.; Wang, X.; Su, M.; Wang, X.; Yan, Q.; Sun, G.; Wang, S. NAPRT, but Not NAMPT, Provides Additional Support for NAD Synthesis in Esophageal Precancerous Lesions. Nutrients 2022, 14, 4916. https://doi.org/10.3390/nu14224916
Wang N, Pan D, Wang X, Su M, Wang X, Yan Q, Sun G, Wang S. NAPRT, but Not NAMPT, Provides Additional Support for NAD Synthesis in Esophageal Precancerous Lesions. Nutrients. 2022; 14(22):4916. https://doi.org/10.3390/nu14224916
Chicago/Turabian StyleWang, Niannian, Da Pan, Xuemei Wang, Ming Su, Xin Wang, Qingyang Yan, Guiju Sun, and Shaokang Wang. 2022. "NAPRT, but Not NAMPT, Provides Additional Support for NAD Synthesis in Esophageal Precancerous Lesions" Nutrients 14, no. 22: 4916. https://doi.org/10.3390/nu14224916