Abstract
Cystic fibrosis (CF) is a life-shortening genetic disorder that affects the cystic fibrosis transmembrane conductance regulator (CFTR) protein. In the gastrointestinal (GI) tract, CFTR dysfunction results in low intestinal pH, thick and inspissated mucus, a lack of endogenous pancreatic enzymes, and reduced motility. These mechanisms, combined with antibiotic therapies, drive GI inflammation and significant alteration of the GI microbiota (dysbiosis). Dysbiosis and inflammation are key factors in systemic inflammation and GI complications including malignancy. The following review examines the potential for probiotic and prebiotic therapies to provide clinical benefits through modulation of the microbiome. Evidence from randomised control trials suggest probiotics are likely to improve GI inflammation and reduce the incidence of CF pulmonary exacerbations. However, the highly variable, low-quality data is a barrier to the implementation of probiotics into routine CF care. Epidemiological studies and clinical trials support the potential of dietary fibre and prebiotic supplements to beneficially modulate the microbiome in gastrointestinal conditions. To date, limited evidence is available on their safety and efficacy in CF. Variable responses to probiotics and prebiotics highlight the need for personalised approaches that consider an individual’s underlying microbiota, diet, and existing medications against the backdrop of the complex nutritional needs in CF.
1. Introduction
Cystic fibrosis (CF) is a genetic condition of autosomal recessive inheritance related to mutations in the gene coding for the cystic fibrosis transmembrane conductance regulator (CFTR) protein [1]. The CFTR protein affects the fluid secretion and mucus hydration of epithelial cells in the airway, pancreas, intestines, and hepatobiliary tracts [2]. Chronic suppurative respiratory disease arising due to impaired clearance of dehydrated airway secretions is typically the principal cause of morbidity and mortality. However, the majority (>90%) of patients with CF also suffer from gastrointestinal (GI) symptoms and complications [3,4]. Dysfunction of the CFTR protein in the GI system results in low intestinal pH, thick and inspissated mucus, a lack of endogenous pancreatic enzymes, reduced motility, and possibly an impaired innate immunity [5,6,7] (Figure 1). These mechanisms are proposed drivers of local GI inflammation and contribute to a range of intestinal morbidities, including an increased risk of early-onset adult GI cancer [8,9,10,11]. GI dysfunction combined with antibiotic therapies also drives significant alteration (dysbiosis) of the GI microbiota (Figure 1). Altered CF microbiota is likely to compound the proinflammatory effects of the underlying disease.
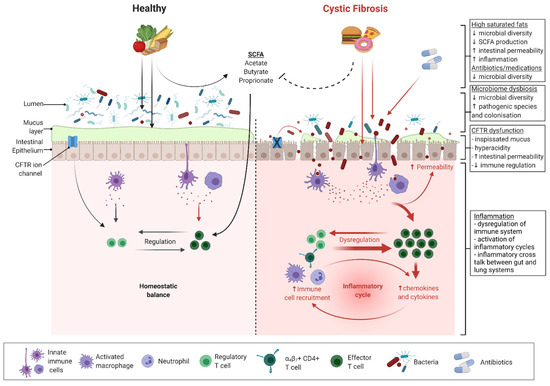
Figure 1.
Microbiome- and CFTR-related dysfunction and inflammation in cystic fibrosis. Black arrows indicate direction of known homeostatic effects. Red arrows indicate direction of known inflammatory effects. Broken lines indicate proposed mechanisms of inhibition or dysfunction. Figure was created with Biorender.com.
Evidence is accumulating that GI bacterial strains, which occur differentially between CF and healthy controls (HC), are linked to inflammatory [12,13,14] and malignancy processes [7,14]. Supplementation with prebiotics and probiotics are thought to provide clinical benefit by promoting commensal bacteria and biosynthesis of immunomodulatory metabolites. As public awareness and acceptance of probiotics and prebiotics continue to expand, there is a growing interest in the potential clinical benefits of dietary prebiotics and probiotics in CF. A total of 17 probiotic trials, including 12 RCTs, have thus far investigated the safety and efficacy of individual probiotic strains and strain combinations in children and adults with CF [15]. Promising improvements in inflammation [16,17,18], nutritional status [19], and health outcomes [20,21] have been observed. However, due to selective reporting and incomplete outcome data, the certainty of evidence has been evaluated as low. Furthermore, large variations between protocols, probiotic formulas, dosage, and duration of treatments limit the potential for clinical application. Prebiotics are often included in probiotic preparations, but evidence surrounding safety and efficacy for the exclusive use of prebiotics is trailing. There is only one clinical trial investigating the prebiotic high-amylose maize starch (HAMS) in adults with CF [22]. The efficacy of prebiotic supplementation is based on the selective utilisationof substrates (usually indigestible carbohydrates) by beneficial bacteria. Critically, it is not yet known whether the altered CF intestinal microbiota retains the capacity to exploit prebiotic substrates.
This review describes the physiology of the GI tract in CF and the clinical relevance of GI microbiome dysbiosis and inflammation. We discuss the current understanding of probiotic and prebiotic mechanisms of action, provide important examples of clinical studies examining probiotic and prebiotic applications in CF, and discuss considerations for clinical translation.
2. Cystic Fibrosis in the Gastrointestinal Tract
CFTR is an important contributor to the normal physiology of the gastrointestinal (GI) tract; as such, its dysfunction in CF disease has profound impacts on GI homeostasis. CFTR is an epithelial cyclic adenosine monophosphate (cAMP)-dependent anion-selective channel. It primarily secretes bicarbonate and chloride, and therefore exerts great influence on the acidity and viscosity of secretions. In CF, the dysfunction of this ion channel is clearly manifested in the systemic production of hyperacidic and viscid mucus [23,24]. CFTR also plays a role in the maintenance of epithelial tight junctions, modulation of fluid flow, regulation of ion channels (such as sodium, potassium, calcium, and other chloride channels [25,26]), and coordination of gut motility [27,28]. Altogether, disruptions to these normal and vital functions of CFTR culminate in an abnormal GI tract (Figure 1).
The altered GI environment in CF results in various clinical sequelae that can be collectively referred to as “obstructive tubulopathies.” The most common of these is pancreatic insufficiency, which affects as much as 90% of patients with CF [29]. From as early as in utero, the presence of concentrated pancreatic ductal secretions leads to luminal protein precipitation with resultant obstruction and dilation of the pancreatic ducts, culminating in progressive, irreversible destruction and fibrosis of the acinar tissue. The resultant pancreas is dysfunctional and severely impaired in its ability to secrete critical enzymes necessary for the digestion of carbohydrates, fats, and proteins [30,31]. Obstructive tubulopathies are also evident in the intestines in the form of meconium ileus (MI) and distal intestinal obstruction syndrome (DIOS). There are numerous other GI manifestations of CF, including gastroesophageal reflux disease, pancreatitis, and liver disease, which have been detailed elsewhere [32,33,34,35,36]. There are also less clinically obvious, but equally significant, manifestations that arise as a result of the altered GI milieu in CF; namely, alterations to the gut microbiota and intestinal inflammation. These are discussed in detail below.
3. The Human Gut Microbiome
The gut microbiome is a sophisticated, functional environment comprising an abundance of microbes along the GI tract. These microorganisms and their metabolites perform homeostatic functions, including the regulation of the gastrointestinal epithelial barrier, fermentation of dietary starches and fibres, synthesis of amino acids and essential vitamins, and modulation of the immune system locally and distally [37,38]. While a small proportion of the gut microbiota is heritable, it is largely influenced by nongenetic factors [39]. The early development of the gut microbiome in infancy is predominantly shaped by one’s mode of birth and feeding, with the cessation of breastfeeding being the driver of functional maturation into an adultlike microbiota [40,41]. Subsequently, diet plays a primary role in shaping the gut microbiome, as organisms respond to selective pressures from dietary patterns throughout life [42,43]. Numerous other environmental factors can also affect the gut microbiota, but perhaps the most well-established are medications, including antibiotics [41,44,45,46].
One important aspect to spotlight when characterising the gut microbiota is microbial diversity. Microbial diversity refers to species richness (the number of species) and/or evenness (the relative distribution of species). Reduced microbial diversity is broadly associated with ill health, as it is hypothesised that species diversity confers the ability to withstand environmental threats and maintain homeostasis. This is attributable to compensatory functional redundancies enabled by a more robust ecological environment [47,48]. Disruption to the normal composition, physiology, and diversity of the gut microbiota is an increasingly recognised feature of numerous disease processes. Decreased microbial diversity has repeatedly been observed in patients with chronic conditions, including obesity, inflammatory bowel disease (IBD), type 1 and type 2 diabetes mellitus, and asthma. Dysbiosis, the collective term for alterations to the normal balance or composition of gut microbes, is also evident in many of those disease processes [48,49,50,51]. These observations point to the critical involvement of the gut microbiota in health and disease, and solidify the rationale for utilising microbial modulation as a therapeutic target.
4. The CF Gut Microbiome
Given the significant alterations to the intestinal environment resulting from CFTR dysfunction, it is unsurprising that the CF gut microbiome differs from that of the healthy gut from early life onwards. One key features of the CF gut microbiome is decreased species diversity [7,12,52,53,54] (Figure 1). In addition, paediatric studies have demonstrated that the CF gut microbiome diversifies and matures at a significantly slower rate than that of a healthy child [54,55,56]. Compositionally, the CF gut microbiome also differs from that of the healthy gut. Reductions in Bacteroidetes, Ruminococcaceae, Bifidobacterium, and Roseburia have consistently been observed. In contrast, abundances of Enterococcus, Veillonella, and Enterobacter have been shown to be relatively increased in the CF gut [7,12,52,53,54,55,57]. Use of the CFTR modulator ivacaftor is associated with arguably “healthier’’ microbiome profiles, reinforcing the concept that dysbiosis is driven by CFTR dysfunction [58]. Recent advancements in metagenomic methods have enhanced the ability to characterise the functionality of the gut microbiota, thereby elucidating the physiological consequences of dysbiosis. It has been demonstrated that the CF gut microbiome displays an increased capacity to metabolise nutrients, antioxidants, and short-chain fatty acids (SCFAs), as well as a relatively decreased propensity to synthesise fatty acids [7,56,59].
The key drivers of these changes to the gut microbiota involve the downstream effects of CFTR dysfunction. The production of dehydrated mucus, changes to intestinal pH, nutrient malabsorption, and prolonged intestinal transit secondary to intestinal dysmotility all have the potential to exert selective pressure on enteric microorganisms and ultimately alter the microbiome [60,61,62]. Notably, fat malabsorption following exocrine pancreatic insufficiency could also confer survival advantage to certain organisms that adapt well to high-fat intestinal environments [63]. These CFTR-related factors are further compounded by iatrogenic causes. Antibiotic exposure, which is prevalent in CF for the prophylaxis and treatment of respiratory tract infections, may contribute to changes in the gut microbiota. Studies in the CF population have consistently demonstrated an association between antibiotic use and decreased alpha diversity (within-sample species diversity) in the gut [12,53,64,65] (Figure 1). Multiple studies have also highlighted a correlation between antibiotic exposure and relative depletions of the bacterial genus Bifidobacterium [64,65,66,67]. The high-energy and high-fat diet prescribed in CF is another likely contributor (discussed below).
5. Intestinal Inflammation
Disruption to the gut microbiota is associated with intestinal inflammation in CF. Chronic inflammation is a well-recognised feature of the CF intestine, primarily evidenced by elevated faecal inflammatory markers in patients with CF in many studies [68,69,70,71,72,73] (Figure 1). The earliest evidence of GI inflammation was elevated concentrations of inflammatory markers such as interleukin-8, interleukin-1β, neutrophil elastase, and immunoglobulins on whole-gut lavage, reported by Smyth et al. [74]. Imaging techniques including endoscopy and capsule endoscopy have subsequently revealed a high prevalence of mucosal pathologies, including ulcerations and oedema in the CF GI tract [71,75,76].
Gut inflammation in CF is of a multifactorial aetiology. Mucus hyperviscosity and hyperacidity as a result of CFTR dysfunction likely promote gut inflammation [8,77,78]. CFTR itself is also involved in downregulating proinflammatory pathways, and hence its dysfunction in CF may contribute to the altered intestinal milieu [79] (Figure 1). Additionally, inflammation may be precipitated by intestinal dysmotility and the intraluminal pooling of inspissated contents [77,80]. The same iatrogenic factors that contribute to intestinal dysbiosis, namely antibiotic exposure and the high-fat CF diet, have also been shown to be correlated with intestinal inflammation in CF and other contexts [81,82,83] (Figure 1). The mechanisms by which antibiotics may induce inflammation are not well-known. However, it has been demonstrated in animal models that antibiotic administration promotes the translocation of microorganisms through goblet-cell-mediated pathways, subsequently increasing the release of inflammatory cytokines [84].
Notably, the aforementioned dysbiosis is a key contributor to intestinal inflammation in CF. Reductions in the abundances of bacteria with anti-inflammatory properties, including Faecalibacterium prausnitzii, has been widely observed in CF cohorts [7,52,53,66,70,85]. Many of these bacteria are known producers of short-chain fatty acids (SCFAs), the primary metabolites of anaerobic fermentation of dietary fibres and starches. SCFAs perform homeostatic functions, including intestinal epithelial maintenance, colonocyte nourishment, and immunomodulation (Figure 1). Accordingly, a relative depletion of SCFA-producing bacteria and subsequent reductions in SCFA levels may contribute to inflammation [60,86,87]. This is supported by numerous animal models in which SCFAs have been shown to improve epithelial integrity and ameliorate intestinal inflammation [88,89,90,91,92,93]. However, the interactions between the microbiota and inflammation are also bidirectional. Chronic inflammation results in the release of reactive oxygen and nitrogen species that supply terminal electron acceptors required for anaerobic respiration. This exerts selective pressure on gut microbes and may contribute to dysbiosis, as organisms with the ability to efficiently perform anaerobic respiration have a growth advantage [94]. For example, intestinal inflammation is associated with the proliferation of Enterobacteriaceae, a bacterial family that has high nitrate reductase activity and can undergo efficient nitrate respiration [95,96]. Indeed, organisms within the Enterobacteriaceae family (i.e., the Enterobacter genus) are relatively more abundant in the CF gut [7,12,64,85]. Many of the mechanistic aspects of the relationship between the gut microbiota and inflammation remain unknown, highlighting the intricacy of these complex interactions.
6. Nutritional Management in CF
In 1988, Corey et al. [97] published a landmark study that led to pivotal paradigm shifts in CF nutritional optimisation. It has since been established that energy requirements are increased in CF due to increased energy expenditure from chronic lung inflammation and increased work of breathing, as well as malabsorption secondary to exocrine pancreatic insufficiency and gastrointestinal disease [29,98,99]. Patients with CF are also at risk of deficiencies in fat-soluble vitamins due to fat and bile acid malabsorption, which often necessitates supplementation [100,101]. Good nutritional status beginning in childhood is now well-documented to be associated with better pulmonary function and survival in CF [102,103,104,105,106]. Body mass index (BMI) is positively correlated with forced expiratory volume in 1 second (FEV1) [107,108,109,110], and a low BMI at the age of 10 years is a risk factor for lung transplantation in adulthood [111]. Greater weight-for-age percentile at the age of 4 years is also associated with better pulmonary function and survival through to 18 years, as well as a reduced likelihood of subsequent pulmonary exacerbations, hospitalisations, or CF-related diabetes [106].
Today, patients with CF are recommended a high-energy diet (110–200% of the age- and sex-appropriate recommended daily energy intake) to maintain growth. While macronutrient targets are individual-specific, the current consensus generally advises that 15–20% of total energy intake be derived from protein, 40–45% from carbohydrates, and up to 35–40% from fat [29,100,112]. Despite the clear benefits of nutritional optimisation, it has become increasingly evident that patients with CF tend to overconsume “energy-dense, nutrient-poor” foods high in salt, sugar, and saturated fat (i.e., junk foods) in order to meet daily macronutrient requirements [113,114,115,116,117]. The proportion of patients with CF who are overweight or obese is increasing. While patients who are overweight or obese are reported to have better lung function than their normal weight or underweight counterparts in some studies, this finding may be confounded by the fact that these patients are also more likely to be pancreatic sufficient and have milder disease genotypes [118,119,120,121,122]. Additionally, weight gain and increased BMI, fat mass, and fat-free mass are reported outcomes of CFTR modulator therapies that need to be taken into account as modulator therapies gradually become the cornerstone of CF treatment [123,124,125,126]. Importantly, both high-fat diets and obesity may exacerbate existing alterations in gut microbial composition and chronic intestinal inflammation, with important clinical implications for individuals with CF [81,82,83,127,128] (Figure 1).
7. Clinical Significance of the CF Gut Microbiome
Intestinal dysbiosis and inflammation have been demonstrated to be significantly associated with clinical outcomes. In a recent study, Hayden et al. [55] observed a distinctly more marked dysbiosis in infants with CF who had low length compared to infants with CF who had normal length. Notably, the gut microbiome of infants with low length exhibited a reduced abundance of Bacteroidetes and relatively delayed maturation compared to that of infants with normal length [55]. Coffey et al. [7] had also previously reported a positive correlation between Ruminococcaceae UCG 014 and BMI. Additionally, the CF intestinal microbiota contains a comparatively lower prevalence of proteins that facilitate carbohydrate transport, metabolism, and conversion, which may impact nutrient utilisation and thus adversely affect growth [85]. It has also been demonstrated that faecal calprotectin levels are inversely correlated with weight and height z-scores, and elevated calprotectin levels are associated with underweight BMI (<18.5 kg/m2) [13,70,72].
Additionally, there is a growing body of evidence that suggests that the intestinal microbiome is related to lung function. It has been reported that patients with lower FEV1 have reduced intestinal microbial diversity compared to their counterparts with better pulmonary function [12]. Positive correlations between intestinal bacterial genera such as Ruminococcaceae NK4A214 and FEV1 have also previously been documented [7]. Furthermore, one study reported an association between microbial diversity in the gut microbiota and pulmonary exacerbation events [57]. Some studies have also demonstrated associations between gut inflammation and lower FEV1, although this has not yet been widely validated [68,72]. It is postulated that these associations reflect a physiological phenomenon termed the “gut–lung axis.” Along this axis, the intestinal and respiratory microbiota engage in cross-talk to regulate immunity and homeostasis in both the enteric and pulmonary environments [129]. In the intestinal compartment, this is achieved by gut-microbiota-derived metabolites, including SCFAs, which coordinate immune cell signaling cascades that ultimately involve the lungs through G-protein coupled receptor (GPCR)-mediated pathways and histone deacetylase inhibition [130,131,132]. In support of this, Hoen et al. [133] observed in a paediatric CF cohort that pulmonary colonisation with the pathogen Pseudomonas aeruginosa, a known contributor to declining lung function, was preceded by a reduction in the abundance of Parabacteroides in the gut. Notably, Parabacteroides is associated with immunomodulation and anti-inflammatory properties [134]. While there remain many unknowns with regard to the mechanistic aspects of the gut–lung axis, these findings suggested that the intestinal microbiome is a site of therapeutic potential that could be manipulated to optimise lung function.
Intestinal dysbiosis and inflammation have been linked to a number of serious morbidities. Firstly, while patients with CF do not typically present with overt GI symptoms similar to those of inflammatory bowel disease (IBD), elevated faecal calprotectin levels are correlated with a worse quality of life [4,135,136]. Elevated calprotectin has also been highlighted as a predictive factor of GI-related hospitalisations for infants with CF in their first year of life [136,137]. Importantly, intestinal dysbiosis and inflammation may be contributors to the increased risk of GI malignancies that is evident in the CF population [138]. While a clear causative mechanism has yet to be established in the context of CF, it is well recognised from studies pertaining to IBD that chronic inflammation poses a significant risk for the development of GI cancers [139,140]. This is largely due to oxidative stress and the resultant DNA damage, culminating in epigenetic disturbances to the expression of tumour-suppressive regulatory proteins, transcription factors, and signalling molecules [141,142]. Furthermore, the inflamed gut may confer a growth advantage to genotoxic organisms, especially E. coli [143]. Indeed, the relative abundance of E. coli is increased in CF, as well as in IBD and colorectal cancer [144,145]. The depletion of SCFA-producing organisms in the CF gut may also be a key factor, as SCFAs exhibit tumour-suppressive properties [146,147]. For example, in an animal model of colitis-associated colorectal cancer, SCFAs have been shown to mediate reductions in proinflammatory cytokine release and tumour size and incidence [148]. All in all, while intestinal dysbiosis and inflammation tend to be clinically silent in CF, they may be associated with serious complications. This emphasises the importance of optimising gut health in the management of CF.
8. Microbiome Modulation with Probiotics
Improving gut health through microbiome modulation is gaining traction in GI and respiratory diseases [149,150]. The microbiome can be modulated through administration of a single or combination of commensal strains (probiotics), indigestible carbohydrates to promote the expansion of commensal strains (prebiotics), or a combination of both (synbionts). Probiotics were first described in 1907, and have been utilised as a beneficial dietary supplement since. In 2002, a consensus was reached by a joint FAO/WHO working group on the definition of probiotics: “Live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” [151]. Existing probiotic preparations are based primarily on strains from the genus lactobacilli, bifidobacterial, and other lactic acid-producing bacteria (LAB) isolated from fermented dairy products and faecal microbiome samples [152]. However, rapidly expanding research into host–microbe interactions is increasing the impetus for the development of next-generation probiotics from beneficial microbes including Akkermansia, Eubacterium, Propionibacterium, Faecalibacterium, and Roseburia species [153,154,155].
Probiotics have reported beneficial effects in diseases with links to a GI dysbiosis, inflammation, and respiratory function [15,152,156]. However, knowledge gaps exist related to robust evidence-based probiotic use as a result of the significant heterogeneity between studies and variability in the probiotic strains studied. Specific probiotic strains have been indicated in the reduction in necrotizing enterocolitis (NEC) incidence [157] and the management of Clostridium difficile [158,159], though the quality of evidence remains low [160]. The rise of in vitro, animal, and cell culture research has expanded our understanding of prosed mechanisms of action, and include direct interaction with commensal gut microbiota, modulation of the immune system, production of organic acids, colonization resistance, improved barrier function, production of hormones and other small molecules with systemic effects, and probiotic–host interactions mediated by cell surface structures [149].
8.1. Mechanisms of Action
Interaction with microbiome. The direct interaction with the microbiome is mediated through the increasing microbial stability [161,162,163], cross-feeding [164], substrate formation, and antagonistic action through competition and production of antimicrobials and bacteriocins [165,166,167]. Competitive exclusion and inhibition of pathogenic species is a primary function of probiotics. In 2007, Collado et al. [168] tested 12 probiotic strains against 8 pathogenic strains in a pig intestinal mucosa model, and found that all probiotic strains tested were able to inhibit and displace pathogenic species of Bacterioides, Clostridium, Staphylococcus and Enterobacter. Another in vitro assay demonstrated that B. animalis subsp. lactis BB-12 and Lactobacillus reuteri DSM 17938 inhibited the growth of pathogenic bacteria E. coli [169]. Likewise, Lactobacillus paracasei FJ861111.1 has demonstrated significant inhibition against several common intestinal pathogens including Shigella dysenteriae, Escherichia coli, and Candida albicans [159].
Modulate immune system. The interaction between microbiota and the immune system, reviewed in [170], has impacts systemwide. Probiotics have been shown to modulate immune function through an increase in anti-inflammatory cytokines [171,172], a reduction in proinflammatory cytokines [149,152,173,174], and augmentation of vaccines and antibody response [175,176,177]. The most common species to demonstrate immune modulation include Lactobacillus, Bacillus, and Bifidobacterium, and the genus Saccharomyces [178]. The modulation of the immune system through probiotics is not consistent across species or strains, and exhibits variability between hosts [149,172]. Yet recently, Sanders et al. [179] identified that some immune modulatory mechanisms related to cell surface infrastructure were conserved across species and even genera.
Production of organic acids. Probiotic species belonging to the Lactobacillus and Bifidobacterium genera produce lactic and acetic acids as end products of carbohydrate metabolism. These organic acids can reduce colonic pH, discouraging the growth of pathogens. Fredua-Agyeman et al. demonstrated that commercial cocultures of Bifidobacterium and Lactobacillus strains inhibited Clostridioides difficile growth in a pH-dependent manner [180]. Through the process of cross-feeding commensal bacterial species such as Faecalibacterium, Lactobacillus, and Bifodobacterium, probiotics can also increase levels of beneficial short-chain fatty acids (SCFA), including butyrate. SCFA have demonstrated anti-inflammatory and antitumour properties [86,181,182].
Improve barrier function. Tight junctions are critical to epithelial cell function, preventing translocation of microbial species and proinflammatory metabolites [183]. Probiotic Lactobacillus and Bifidobacterium strains have been shown to increase the expression of tight junction proteins [184,185] and reduce the severity of acute gastroenteritis in children through fortification of tight junctions [186]. Several Lactobacillus probiotic strains have also demonstrated regulatory effects on the epithelial mucus layer [153,187,188,189]. The demonstrated upregulation of mucin production genes and enhanced mucin secretion improves barrier function, inhibiting pathogen binding to epithelial cells [185].
Production of small molecules with systemic effects. Probiotic strains have been implicated in the production of a range of small molecules and hormones that influence systemic function. Interestingly, these include neurotransmitters such as cortisol, serotonin, tryptamine, noradrenaline gamma-aminobutyric acid (GABA), and dopamine, highlighting the potential of probiotics to modulate the gut–brain axis [190,191]. A range of satiety hormones and enzymes that can aid digestion are also produced by some probiotic strains. For example, Streptococcus thermophilus can facilitate lactose digestion through the production of microbial β-galactosidase [192].
Probiotic–host interactions mediated by cell surface structures. The cell surface architecture of probiotic strains is critical to probiotic–host cell interactions. Many Gram-positive probiotic strains share cell surface macromolecules that mediate these interactions, including surface layer associated proteins (SLAPS), mucin-binding proteins (MUBs), fibronectin binding proteins, and pili that interact directly with the intestinal epithelium, mucus, and gastrointestinal mucosa receptors. These demonstrated interactions reviewed in [179] can improve host barrier integrity, intestinal motility, and binding to intestinal and vaginal cells.
8.2. Probiotics in CF
The use of probiotics in CF has been investigated in 17 clinical trials. Of those 17 trials, 12 were RCTs, with 8 trials including children and 4 trials including both children and adults [15]. The number of subjects within the RCTs ranged from 22 to 81, and the trial duration ranged from 1 month to 12 months. The probiotic formulations varied in dosage from 108 CFU/day to 1011 CFU/day. Strain formulations with six of the trials utilised a single Lactobacillus strain L rhamnosus GG [16,19,193,194] or L. reuteri [18,20,195], two trials utilised multistrain formulations with fructooligosaccharides (FOS) [17,66], and three trials utilised a multistrain without FOS [18,21,196]. Results from individual trials have cited a reduction in in inflammation [16,17,18], nutritional status [19], and pulmonary health outcomes [20,21] (Table 1).

Table 1.
Evidence for the use of probiotics in CF from RCTs.
From 2016 to 2021, six systematic reviews have attempted to synthesise the expanding evidence for probiotics in CF [15,197,198,199,200,201]. The first review in 2016 [201] examined a total of nine trials with a total of 275 subjects, and found that probiotics were likely to decrease gut dysbiosis and improve gut maturity and function. In 2017, three more reviews were published that were broadened to include evidence on pulmonary exacerbations and quality-of-life indicators [197,199,200]. The latest and most comprehensive systematic review was based on data from the 12 RCTs only [15]. Combined data from four trials (225 participants) found that probiotics may reduce pulmonary exacerbations when administered over a four-to-12-month period mean difference (MD) of −0.32 episodes per participant (95% confidence interval (CI) −0.68 to 0.03; p = 0.07)). The 95% confidence intervals included the possibility of both an increased and deceased number of exacerbations. The combined data from four trials (177 participants) also indicated that probiotics may reduce faecal calprotectin, MD −47.4 µg/g (95% CI −93.28 to −1.54; p = 0.04). Due to (i) a high risk of bias due to selective reporting; (ii) a high risk of bias due to incomplete outcome data; and (iii) a lack of generalisability, the evidence for these results was evaluated as low certainty [15].
The results from other biomarkers and health outcomes including lung function (forced expiratory volume at one second (FEV1)% predicted) (five trials, 284 participants); duration of antibiotic therapy (two trials, 127 participants); hospitalisation rates (two trials, 115 participants); height, weight, or body mass index (two trials, 91 participants); and reported health-related quality of life scores (1 trial, 37 participants) did not demonstrate any difference between placebo and treatment groups, (all low-certainty evidence). Only two studies included a microbial analysis, and insufficient data was available to analyse in the systematic review. Likewise, there was insufficient evidence to evaluate gastrointestinal symptoms. The probiotics evaluated in the RCTs were associated with four adverse events, including vomiting, diarrhoea, and allergic reactions [15].
Results from individual trials and systematic reviews have consistently indicated that probiotics are likely to have beneficial effects in CF, especially for inflammation and pulmonary exacerbations. However, all systematic reviews cited a limited amount of low-quality data as a barrier to justifying the inclusion of probiotics in current CF treatment protocols [15,197,198,199,200,201]. Furthermore, a high variation between trial protocols, probiotic formulation, dose, duration of therapy, and clinical outcomes measured make predictions about effective strains and dosages and clinical translation difficult. To address data quality, Coffey et al. [15] recommended multicentre RCTs of at least 12 months duration to best assess the efficacy and safety of probiotics for children and adults with CF.
9. Microbiome Modulation with Prebiotics
Modulation of the microbiome can also be targeted through the administration of generally non-digestible compounds known as prebiotics. Prebiotics provide health benefits by promoting the proliferation of commensal gut species and subsequent production of beneficial metabolites [202]. Prebiotics were first defined in 1995, with an updated definition published in 2017 as “a substrate that is selectively utilized by host microorganisms conferring a health benefit” [203]. Prebiotics occur naturally in foods such as breads, cereals, onions, garlics, and artichokes [204] but are also available as dietary supplements. The most established and well documented prebiotics include inulin, fructo-oligosaccharides (FOS), oligofructose, galacto-oligosaccharides (GOS), and lactulose [205,206]. Other potential prebiotics with expanding evidence of effect include resistant starch, high amylose maize starch (HMAS), glucans, arabinoxylan oligosaccharides, xylooligosaccharides, soybean oligosaccharides, isomalto-oligosaccharides, and pectin [206,207].
9.1. Mechanisms of Action
The underlying hypothesis of prebiotics is that the additional fermentable substrates drive the proliferation of keystone commensal bacteria, and subsequently the production of beneficial metabolites such as SCFA [152]. Commensal bacteria and SCFA metabolites then directly and indirectly improve host health through colonisation inhibition, increased barrier integrity, and immune modulation [183]. Currently, the mechanisms of action postulated for prebiotics are primarily based on in vitro models. Validation of proposed mechanisms and demonstrated effects in human models is limited.
Interaction and modulation of microbiome. Prebiotics in the form of undigestible carbohydrates promote proliferation of beneficial bacteria. As mentioned above, subsequent health benefits are a direct result of increased beneficial bacteria such as colonisation inhibition, or an indirect result of increased beneficial metabolite production such as improved barrier function and immune modulation [183]. In a randomised, double-blind, placebo-controlled clinical trial, a prebiotic intervention with GOS reduced intestinal permeability in obese adults. The probiotic intervention of Bifidobacterium adolescentis also reduced intestinal permeability, but interestingly, no synergistic effect was observed when the two were combined [208].
Defence against pathogens. As with probiotics, the modulation of the microbiome through prebiotics results in the generation of organic acids, reducing luminal pH, which inhibits growth of pathogens. The increase in commensal species also reduces nutrient availability for invasive species as described above. There is also evidence to suggest that GOS prebiotics can directly interfere with E. coli adhesion to tissue culture cells [209].
Metabolic effects. The metabolic effects of prebiotics have been the subject of several meta-analyses [210,211,212]. The evidence suggests that GOS and inulin can reduce high sensitivity C-reactive protein, plasma cholesterol, triglycerides, and fasting plasma insulin associated with obesity and diabetes. The exact mechanisms of action, duration of effects, or results from long-term consumption has not yet been established [211].
Immune modulation. Prebiotic immune modulation is primarily activated through microbial fermentation and subsequent production of SCFA metabolites. However, some prebiotics have been demonstrated to bind directly to some immune cell receptors [183,213]. Immune modulation is not consistent between probiotic categories or conditions, or even within conditions. A RCT with 259 infants concluded that GOS and long-chain FOS administered in formula may regulate immune function in infants, with a 50% reduction in atopic dermatitis, wheezing, and urticaria to when compared to non-prebiotic formula-fed infants [214]. Yet, a subsequent multicentre RCT with 365 infants found that while GOS supplementation altered faecal frequency and consistency, there was no effect on incidence of infection or allergic manifestation during the first year of life [215]. Likewise, conflicting studies in elderly individuals have proposed prebiotic GOS supplementation may have either no effect on immune function [216], or may increase immune function through enhanced phagocytic activity and activity of natural killer cells [217,218].
9.2. Prebiotics in CF
Prebiotics have been combined with probiotics in synbiotic preparations for use in CF probiotic trials [17,66,219]. However, only one study has investigated the exclusive use of prebiotics in CF for GI microbiome modulation [22]. Effective clinical use of prebiotics assumes a selective utilisation of the supplemented substrate by the recipient’s microbiota. It is not yet known if the disrupted microbiota in CF that is depleted in key SCFA-producing organisms, has the functional capacity to utilise prebiotic substrates. In a pilot study, Wang et al. [22] used a combination of metagenomic sequencing, invitro fermentation, amplicon sequencing, and metabolomics to investigate the HAMS fermentation capacity of 19 adults with CF and 16 non-CF controls. They demonstrated that despite low abundances of common taxa attributed to fermentation of HMAS (Faecalibacterium, Roseburia and Coprococcus), the production of butyrate and propionate was consistent with healthy control slurries, while the production of acetate was reduced. In the absence of Faecalibacterium, the CF SCFA biosynthesis was attributed to Clostridium ss1. Importantly, in a subset of CF patients, the presence of HAMS led to enterococcal overgrowth and the accumulation of lactate [22]. Likewise, a murine study found that supplementation with purified prebiotics inulin, fructooligosaccharides, or pectin may result in hepatocellular carcinoma in mice with pre-existing perturbed microbial communities [220]. These results demonstrated the potential for variable responses to prebiotics, dependent on the underlying microbiome [220].
In the absence of further CF-specific research, we examined evidence of prebiotics in GI disorders with an inflammatory link, including ulcerative colitis (UC), Crohn’s disease, colorectal cancer (CRC), and chronic respiratory disease including Psuedomonas aeruginosa infections, asthma, and emphysema (Table 2). In colitis animal models, a range of prebiotics including 2-fructosyl lactose [221], barley leaf insoluble dietary fibre (BLIDF) [222], psyllium [223], wheat bran [224], and butyrate [225] have been shown to reduce colitis symptoms and inflammatory markers, while increasing bacterial diversity and SCFA levels. Specific inflammatory markers, bacterial species, and SCFA vary between studies and prebiotic type (Table 2). The effects of inulin-type fructins (ITF) at 7.5 g/day (n = 12) or 15 g/day (n = 13) were tested in a human trial with 25 patients with mild/moderate UC [226]. The high-ITF-dose group showed significantly reduced colitis and calprotectin concentrations, and increased butyrate levels. The bacterial species Bifidobacteriaceae and Lachnospiraceae also increased in the high-dose group, but their abundance was not correlated to improved disease scores. The lack of taxonomic correlation suggested that functional shifts may be more relevant than compositional shifts in UC (Table 2). A RCT involving 140 preoperative patients with CRC investigated the role of prebiotics (fructooligosaccharides, xylooligosaccharides, polydextrose, and resistant dextrin) on immune function and intestinal microbiota. They reported that probiotics led to improved serum immunological markers and abundances of commensal bacterial species before surgeries [227]. Prebiotics did not protect from surgery-related microbial stress observed in both postoperative groups (Table 2).
Table 2.
Evidence of prebiotics and dietary effects on microbiome in chronic inflammatory and respiratory disease.
In 2015, a mouse model study investigated the effects of dietary-pectin-derived acidic oligosaccharides (pAOS) on Pseudomonas (P) aeruginosa, and found that pAOS may limit the number and severity of pulmonary exacerbations chronically infected with P. aeruginosa [232] (Table 2). These results are highly relevant to individuals with CF, but have not been validated in human trials. In another small RCT study, either a placebo or a galactooligosaccharides (GOS) preparation highly selective to Bifidobacterium (B-GOS) was given to 10 adults with asthma-associated, hyperpnoea-induced bronchoconstriction (HIB) and 8 adult controls. Pulmonary function remained unchanged in the control group, but in the HIB group, FEV1 was attenuated by 40%, and the baseline chemokine CC ligand and TNF-a was reduced after B-GOS supplementation [237] (Table 2). As with probiotics, the large number and source of prebiotics limits the ability to effectively compare treatments across studies and conditions.
9.3. Microbiome Modulation with Diet
There is an increasingly apparent link between diet, microbiome modulation, and host health [238,239]. In the first few years of life, the interactions between diet and the microbiome are especially relevant. The oligosaccharides present in breast milk encourage the colonization of Bifidobacterium spp. The subsequent metabolites produced by Bifidobacterium spp. support the expansion of the microbiome through cross-feeding, and promote immune tolerance to other commensal bacteria [240,241]. The progression of the microbiome develops alongside increases in diet diversity, with the introduction of solid food triggering a rapid expansion in the bacterial community and the subsequent quantity and variety of metabolites. Metabolites associated with solid food ingestion, primarily butyrate, have been demonstrated to drive the maturation of the mucosal barrier in Caco-2 cells [242], which is critical to colonization inhibition of pathogens [243]. As mentioned earlier, iatrogenic factors such as antibiotic use can disrupt microbiome progression [41]. More broadly, early life microbiome disruption has been implicated in the development of autoimmune disease in mouse models [244], and has been associated with lasting metabolic and autoimmune disease consequences in observational studies [245,246,247,248].
The progression of the microbiome continues until it establishes near-adult levels of diversity at 3–5 years of age. Once established, the microbiome is more resistant to disruption [249]. However, diet remains a key modulator, with accumulating evidence of the role of dietary components in local inflammation, intestinal barrier function, and host immune dysregulation [220]. High-fat diets promote the translocation of certain bacteria by enhancing intestinal permeability and preferentially increasing the relative abundances of lipopolysaccharide-bearing bacteria [82,83,250,251,252,253]. Dietary fibres microbially fermented in the gut lead to the production of short-chain fatty acid metabolites, which have demonstrated roles in immune regulation [254,255,256], maintenance of epithelial barrier [223,257], and microbiome modification [258]. Low-fibre diets also have long-term implications for cancer [259] and metabolic and autoimmune diseases [260,261].
9.4. Diet in CF
Consistent with the general population, early-life microbiome development in CF is correlated to infant breastfeeding [262] and the initiation of solid foods [133]. Infant breastfeeding and solid food intake are further linked to the respiratory microbiome [262] and health outcome indicators, supporting the concept of a gut–lung axis in CF [133]. The progression of the CF microbiome is disrupted in early life [53,54,59], characterized by reduced taxonomic and functional profiles (dysbiosis) [7] that persist into adulthood [12]. No dietary fibre interventions have been trialled in CF, but epidemiological studies and preclinical and clinical trials support the potential of fibre to modulate the microbiome structure [234] and improve function in chronic gastrointestinal [263] and respiratory conditions [235,264].
A low-fat, high-fibre diet in individuals with ulcerative colitis improved the quality of life (QoL) as quantified through the QoL IBD survey. The intervention led to increased acetate and tryptophan levels and modulated the microbiome, increasing the abundance of Bacteroidetes and Faecalibacterium [228] (Table 2). A high-fibre diet also reduced inflammation and attenuated pathological changes associated with emphysema in an emphysema mouse model exposed to cigarette smoke. Alveolar destruction and inflammatory cytokines in bronchoalveolar lavage fluid (BALF) were reduced, while SCFA were increased [235] (Table 2). In contrast, saturated fat decreased microbial diversity and increased colitis severity and the TH1 mucosal response in a DSS-induced colitis mouse model [236] (Table 2). In 37 adults with asthma, high-fat intake was also demonstrated to increase inflammation and attenuate both the duration and magnitude of recovery from an aerosol-administered bronchodilator. However, the potential role of the microbiome was not investigated [265]. A range of micronutrients and antioxidant supplements have been trialled in CF to ameliorate fat-soluble vitamin deficiencies, altered fatty-acid synthesis, and increased oxidative stress [266]. In 2020, a meta-analysis concluded that the benefits sometimes observed across 8 antioxidant and 15 essential-fatty-acid supplementation studies was not consistent enough to recommend their routine use in CF [266]. With the exception of an exploratory study correlating vitamin D insufficiency with increases in potential pathogenic species [267], the effect of these supplementations on the microbiome have not been widely explored.
10. Considerations for Clinical Application and Future Studies
A survey from a CF clinic in the USA found that 60% of CF patients currently use some type of probiotic [268], despite the majority of probiotics available for sale having little to no evidence to support effectiveness, dose, or disease specificity [269]. Conditional use of specific probiotic strains in (non-CF) gastrointestinal disorders has been recommended for prevention of antibiotic-associated C. difficile infections, pouchitis, and prevention of necrotizing enterocolitis [160]. While these conditions may impact on individuals with CF, recommendations have not been validated specifically in CF. Evidence from CF-specific RCTs suggest probiotics are likely to improve GI inflammation and reduce the incidence of pulmonary exacerbations. Yet, the highly variable, low-quality data has been insufficient to determine ideal strains, dosage, and treatment duration, constraining the implementation of probiotics into routine CF care.
The evidence for prebiotics lags behind that for probiotics. There was only one clinical trial for the use of prebiotics in CF, and the results were mixed. Some individuals demonstrated successful microbial modulation and an increased production of butyrate and propionate. However, a subset of CF reactions exhibited enterococcal overgrowth, resulting in lactate accumulation and reduced SCFA biosynthesis [22]. The altered microbiome and predisposition to pathogenic overgrowth in CF highlights the need for standardised preparations of well-characterised prebiotics to be investigated specifically within the CF population to adequately evaluate safety and efficacy.
The increased energy needs of individuals with CF are currently being meet through “energy-dense, nutrient-poor” diets with excess saturated fats and inadequate fibre intakes [113]. As previously discussed, high-fibre, low-fat diets are associated with improved inflammation, immune modulation, and gut barrier function outcomes. While simply adding more dietary fibre to existing CF diets is tantalizing, interventional dietary trials have demonstrated that increasing dietary fibre intake does not necessarily translate to increased SCFA production or improvements in disease outcomes [220,270]. This is pertinent to members of the CF population, who are likely to have altered SCFA-generating pathways [7,56,60]. High-quality randomized trials with well-defined dietary components are essential to provide justification for modulating microbiome–host effects through diet. Considering the uptake of highly effective modulator therapies, a re-evaluation of dietary recommendations with a focus on diet quality and individual energy requirements is also recommended.
As outlined in previous reviews, there is a need for large, well-designed, longitudinal and multicentred clinical trials to effectively evaluate the safety and efficacy of probiotics in CF. While this is also true for prebiotics, there is a paucity of prebiotics research in CF and a plethora of substrates with prebiotic potential. The use of organoids, cell lines, and or animal models may be an economic option to demonstrate beneficial effects and mechanisms of action across a variety of compounds before proceeding to clinical trials. Likewise, there are a variety of dietary interventions and supplements that may have beneficial host–microbiome effects. High-quality, randomized studies with well-defined compounds are needed to evaluate safety and efficacy in CF before dietary interventions or supplements can be utilised to modulate the CF microbiome.
11. Conclusions
Although the exact mechanisms are not yet fully elucidated, the host–microbiome interactions in CF are critical to the incidence of GI inflammation and disease.
Targeted diet-based therapies provide an opportunity to modulate the altered CF microbiome to counter the early disruption to microbiome progression, and could transform GI and respiratory disease outcomes. The expansion of metagenomic, proteomic, and transcriptomic analyses continues to illuminate CF specific taxonomic and functional alterations. This knowledge will be critical to the development of next-generation precision probiotics and prebiotics. For now, there is insufficient evidence to support the safe and effective use of prebiotics in CF, but probiotics and a re-evaluation of the CF diet may be beneficial. Critically, there is a need for personalised approaches that understand an individual’s baseline microbiota and can manage potential microbiome-modulating therapies alongside existing medications and complex nutritional needs.
Author Contributions
Conceptualization, J.M.v.D. and C.Y.O.; writing-original draft preparation, J.M.v.D. and R.Y.T.; writing-review and editing J.M.v.D. and C.Y.O.; visualization J.M.v.D.; supervision C.Y.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Riordan, J.R.; Rommens, J.M.; Kerem, B.-S.; Alon, N.; Rozmahel, R.; Grzelczak, Z.; Zielenski, J.; Lok, S.; Plavsic, N.; Chou, J.-L.; et al. Identification of the cystic fibrosis gene: Cloning and characterization of complementary DNA. Science 1989, 245, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Riordan, J.R. CFTR function and prospects for therapy. Annu. Rev. Biochem. 2008, 77, 701–726. [Google Scholar] [CrossRef] [PubMed]
- Rowbotham, N.J.; Smith, S.; A Leighton, P.; Rayner, O.C.; Gathercole, K.; Elliott, Z.C.; Nash, E.F.; Daniels, T.; A Duff, A.J.; Collins, S.; et al. The top 10 research priorities in cystic fibrosis developed by a partnership between people with CF and healthcare providers. Thorax 2018, 73, 388–390. [Google Scholar] [CrossRef]
- Tabori, H.; Arnold, C.; Jaudszus, A.; Mentzel, H.-J.; Renz, D.M.; Reinsch, S.; Lorenz, M.; Michl, R.; Gerber, A.; Lehmann, T.; et al. Abdominal symptoms in cystic fibrosis and their relation to genotype, history, clinical and laboratory findings. PLoS ONE 2017, 12, e0174463. [Google Scholar] [CrossRef]
- Ooi, C.Y.; Durie, P.R. Cystic fibrosis from the gastroenterologist’s perspective. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 175–185. [Google Scholar] [CrossRef]
- Ooi, C.Y.; Pang, T.; Leach, S.T.; Katz, T.; Day, A.S.; Jaffe, A. Fecal Human β-Defensin 2 in Children with Cystic Fibrosis: Is There a Diminished Intestinal Innate Immune Response? Dig. Dis. Sci. 2015, 60, 2946–2952. [Google Scholar] [CrossRef]
- Coffey, M.J.; Nielsen, S.; Wemheuer, B.; Kaakoush, N.O.; Garg, M.; Needham, B.; Pickford, R.; Jaffe, A.; Thomas, T.; Ooi, C.Y. Gut Microbiota in Children With Cystic Fibrosis: A Taxonomic and Functional Dysbiosis. Sci. Rep. 2019, 9, 18593. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Ooi, C.Y. The Enigmatic Gut in Cystic Fibrosis: Linking Inflammation, Dysbiosis, and the Increased Risk of Malignancy. Curr. Gastroenterol. Rep. 2017, 19, 6. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, P.; Marshall, B.C.; Knapp, E.A.; Lowenfels, A.B. Cancer risk in cystic fibrosis: A 20-year nationwide study from the United States. J. Natl. Cancer Inst. 2013, 105, 122–129. [Google Scholar] [CrossRef]
- Maisonneuve, P.; Fitzsimmons, S.C.; Neglia, J.; Campbell, P.W.; Lowenfels, A.B. Cancer risk in nontransplanted and transplanted cystic fibrosis patients: A 10-year study. J. Natl. Cancer Inst. 2003, 95, 381–387. [Google Scholar] [CrossRef]
- Meyer, K.C.; Francois, M.L.; Thomas, H.K.; Radford, K.L.; Hawes, D.S.; Mack, T.L.; Cornwell, R.D.; Maloney, J.D.; De Oliveira, N.C. Colon cancer in lung transplant recipients with CF: Increased risk and results of screening. J. Cyst. Fibros. 2011, 10, 366–369. [Google Scholar] [CrossRef][Green Version]
- Burke, D.; Fouhy, F.; Harrison, M.J.; Rea, M.C.; Cotter, P.D.; O’Sullivan, O.; Stanton, C.; Hill, C.; Shanahan, F.; Plant, B.J.; et al. The altered gut microbiota in adults with cystic fibrosis. BMC Microbiol. 2017, 17, 58. [Google Scholar] [CrossRef]
- Dhaliwal, J.; Leach, S.; Katz, T.; Nahidi, L.; Pang, T.; Lee, J.; Strachan, R.; Day, A.S.; Jaffe, A.; Ooi, C.Y. Intestinal inflammation and impact on growth in children with cystic fibrosis. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 521–526. [Google Scholar] [CrossRef]
- Pang, T.; Leach, S.T.; Katz, T.; Jaffe, A.; Day, A.S.; Ooi, C.Y. Elevated fecal M2-pyruvate kinase in children with cystic fibrosis: A clue to the increased risk of intestinal malignancy in adulthood? J. Gastroenterol. Hepatol. 2015, 30, 866–871. [Google Scholar] [CrossRef]
- Coffey, M.J.; Garg, M.; Homaira, N.; Jaffe, A.; Ooi, C.Y. A systematic cochrane review of probiotics for people with cystic fibrosis. Paediatr. Respir. Rev. 2021, 39, 61–64. [Google Scholar] [CrossRef]
- Bruzzese, E.; Callegari, M.L.; Raia, V.; Viscovo, S.; Scotto, R.; Ferrari, S.; Morelli, L.; Buccigrossi, V.; Vecchio, A.L.; Ruberto, E.; et al. Disrupted intestinal microbiota and intestinal inflammation in children with cystic fibrosis and its restoration with Lactobacillus GG: A randomised clinical trial. PLoS ONE 2014, 9, e87796. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, G.; Motamed, F.; Yousefi, A.; Shafieyoun, A.; Najafi, M.; Khodadad, A.; Farhmand, F.; Ahmadvand, A.; Rezaei, N. The effect of probiotics on fecal calprotectin in patients with cystic fibrosis. Turk. J. Pediatr. 2013, 55, 475–478. [Google Scholar]
- Del Campo, R.; Garriga, M.; Agrimbau, J.; Lamas, A.; Maiz, L.; Canton, R.; Suarez, L. Improvement of intestinal comfort in cystic fibrosis patients after probiotics consumption. J. Cyst. Fibros. 2009, 8, S89. [Google Scholar] [CrossRef]
- Bruzzese, E.; Raia, V.; Spagnuolo, M.I.; Volpicelli, M.; De Marco, G.; Maiuri, L.; Guarino, A. Effect of Lactobacillus GG supplementation on pulmonary exacerbations in patients with cystic fibrosis: A pilot study. Clin. Nutr. 2007, 26, 322–328. [Google Scholar] [CrossRef]
- Di Nardo, G.; Oliva, S.; Menichella, A.; Pistelli, R.; De Biase, R.V.; Patriarchi, F.; Cucchiara, S.; Stronati, L. Lactobacillus reuteri ATCC55730 in cystic fibrosis. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 81–86. [Google Scholar] [CrossRef]
- Jafari, S.-A.; Mehdizadeh-Hakkak, A.; Kianifar, H.-R.; Hebrani, P.; Ahanchian, H.; Abbasnejad, E. Effects of probiotics on quality of life in children with cystic fibrosis: A randomized controlled trial. Iran. J. Pediatrics 2013, 23, 669. [Google Scholar]
- Wang, Y.; Leong, L.E.; Keating, R.L.; Kanno, T.; Abell, G.C.; Mobegi, F.M.; Choo, J.M.; Wesselingh, S.L.; Mason, A.J.; Burr, L.D.; et al. Opportunistic bacteria confer the ability to ferment prebiotic starch in the adult cystic fibrosis gut. Gut Microbes 2019, 10, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Venkatasubramanian, J.; Ao, M.; Rao, M.C. Ion transport in the small intestine. Curr. Opin. Gastroenterol. 2010, 26, 123–128. [Google Scholar] [CrossRef] [PubMed]
- De Lisle, R.C.; Borowitz, D. The cystic fibrosis intestine. Cold Spring Harb. Perspect. Med. 2013, 3, a009753. [Google Scholar] [CrossRef] [PubMed]
- Liou, T.G. The Clinical Biology of Cystic Fibrosis Transmembrane Regulator Protein: Its Role and Function in Extrapulmonary Disease. Chest 2019, 155, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.; Anderson, K.; Singhania, M.; Cormier, R. Cystic Fibrosis, CFTR, and Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 2891. [Google Scholar] [CrossRef]
- Xue, R.; Gu, H.; Qiu, Y.; Guo, Y.; Korteweg, C.; Huang, J.; Gu, J. Expression of Cystic Fibrosis Transmembrane Conductance Regulator in Ganglia of Human Gastrointestinal Tract. Sci. Rep. 2016, 6, 30926. [Google Scholar] [CrossRef] [PubMed]
- Yeh, K.M.; Johansson, O.; Le, H.; Rao, K.; Markus, I.; Perera, D.S.; Lubowski, D.Z.; King, D.W.; Zhang, L.; Chen, H.; et al. Cystic fibrosis transmembrane conductance regulator modulates enteric cholinergic activities and is abnormally expressed in the enteric ganglia of patients with slow transit constipation. J. Gastroenterol. 2019, 54, 994–1006. [Google Scholar] [CrossRef]
- Saxby, N.; Painter, C.; Kench, A.; King, S.; Crowder, T.; van der Haak, N.; The Australian and New Zealand Cystic Fibrosis Nutrition Guideline Authorship Group. Nutrition Guidelines for Cystic Fibrosis in Australia and New Zealand; Bell, S.C., Ed.; Thoracic Society of Australia and New Zealand: Sydney, Australia, 2017. [Google Scholar]
- Li, L.; Somerset, S. Digestive system dysfunction in cystic fibrosis: Challenges for nutrition therapy. Dig. Liver Dis. 2014, 46, 865–874. [Google Scholar] [CrossRef]
- Durie, P.R.; Forstner, G.G. Pathophysiology of the exocrine pancreas in cystic fibrosis. J. R. Soc. Med. 1989, 82 (Suppl. 16), 2–10. [Google Scholar]
- Ooi, C.Y.; Durie, P.R. Cystic fibrosis transmembrane conductance regulator (CFTR) gene mutations in pancreatitis. J. Cyst. Fibros. 2012, 11, 355–362. [Google Scholar] [CrossRef]
- Ooi, C.Y.; Dorfman, R.; Cipolli, M.; Gonska, T.; Castellani, C.; Keenan, K.; Freedman, S.D.; Zielenski, J.; Berthiaume, Y.; Corey, M.; et al. Type of CFTR mutation determines risk of pancreatitis in patients with cystic fibrosis. Gastroenterology 2011, 140, 153–161. [Google Scholar] [CrossRef]
- Wilschanski, M.; Durie, P.R. Patterns of GI disease in adulthood associated with mutations in the CFTR gene. Gut 2007, 56, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Jaclyn, R.B.; Kenneth, J.F.; Simon, C.L.; Rhonda, G.P.; Scott, C.B.; Billy, B.; Giuseppe, C.; Carlo, C.; Marco, C.; Carla, C.; et al. Genetic modifiers of liver disease in cystic fibrosis. JAMA 2009, 302, 1076–1083. [Google Scholar] [CrossRef]
- Gelfond, D.; Borowitz, D. Gastrointestinal complications of cystic fibrosis. Clin. Gastroenterol. Hepatol. 2013, 11, 333–342, quiz e30-1. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Roy, B.C.; Khan, S.A.; Septer, S.; Umar, S. Microbiome, Metabolome and Inflammatory Bowel Disease. Microorganisms 2016, 4, 20. [Google Scholar] [CrossRef]
- Clemente, J.C.; Manasson, J.; Scher, J.U. The role of the gut microbiome in systemic inflammatory disease. BMJ 2018, 360, j5145. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.D.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjölund-Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-term antibiotic treatment has differing long-term impacts on the human throat and gut microbiome. PLoS ONE 2010, 5, e9836. [Google Scholar] [CrossRef]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-level analysis of gut microbiome variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.A.; Goodrich, J.K.; Maxan, M.-E.; Freedberg, D.E.; Abrams, J.A.; Poole, A.; Sutter, J.L.; Welter, D.; Ley, R.; Bell, J.; et al. Proton pump inhibitors alter the composition of the gut microbiota. Gut 2016, 65, 749–756. [Google Scholar] [CrossRef]
- Sommer, F.; Anderson, J.M.; Bharti, R.; Raes, J.; Rosenstiel, P. The resilience of the intestinal microbiota influences health and disease. Nat. Rev. Microbiol. 2017, 15, 630–638. [Google Scholar] [CrossRef]
- Valdes, A.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome—A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef]
- Miragoli, F.; Federici, S.; Ferrari, S.; Minuti, A.; Rebecchi, A.; Bruzzese, E.; Buccigrossi, V.; Guarino, A.; Callegari, M.L. Impact of cystic fibrosis disease on archaea and bacteria composition of gut microbiota. FEMS Microbiol. Ecol. 2017, 93, fiw230. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, P.; Del Chierico, F.; Russo, A.; Majo, F.; Rossitto, M.; Valerio, M.; Casadei, L.; La Storia, A.; De Filippis, F.; Rizzo, C.; et al. Gut microbiota signatures in cystic fibrosis: Loss of host CFTR function drives the microbiota enterophenotype. PLoS ONE 2018, 13, e0208171. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Needham, B.; Leach, S.T.; Day, A.S.; Jaffe, A.; Thomas, T.; Ooi, C. Disrupted progression of the intestinal microbiota with age in children with cystic fibrosis. Sci. Rep. 2016, 6, 24857. [Google Scholar] [CrossRef] [PubMed]
- Hayden, H.S.; Eng, A.; Pope, C.E.; Brittnacher, M.J.; Vo, A.T.; Weiss, E.J.; Hager, K.R.; Martin, B.D.; Leung, D.H.; Heltshe, S.L.; et al. Fecal dysbiosis in infants with cystic fibrosis is associated with early linear growth failure. Nat. Med. 2020, 26, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Eng, A.; Hayden, H.S.; Pope, C.E.; Brittnacher, M.J.; Vo, A.T.; Weiss, E.J.; Hager, K.R.; Leung, D.H.; Heltshe, S.L.; Raftery, D.; et al. Infants with cystic fibrosis have altered fecal functional capacities with potential clinical and metabolic consequences. BMC Microbiol. 2021, 21, 247. [Google Scholar] [CrossRef]
- Antosca, K.M.; Chernikova, D.A.; Price, C.E.; Ruoff, K.L.; Li, K.; Guill, M.F.; Sontag, N.R.; Morrison, H.G.; Hao, S.; Drumm, M.L.; et al. Altered Stool Microbiota of Infants with Cystic Fibrosis Shows a Reduction in Genera Associated with Immune Programming from Birth. J. Bacteriol. 2019, 201, e00274-19. [Google Scholar] [CrossRef] [PubMed]
- Ooi, C.Y.; Syed, S.A.; Rossi, L.; Garg, M.; Needham, B.; Avolio, J.; Young, K.; Surette, M.G.; Gonska, T. Impact of CFTR modulation with Ivacaftor on Gut Microbiota and Intestinal Inflammation. Sci. Rep. 2018, 8, 17834. [Google Scholar] [CrossRef] [PubMed]
- Manor, O.; Levy, R.; Pope, C.E.; Hayden, H.S.; Brittnacher, M.J.; Carr, R.; Radey, M.C.; Hager, K.R.; Heltshe, S.L.; Ramsey, B.W.; et al. Metagenomic evidence for taxonomic dysbiosis and functional imbalance in the gastrointestinal tracts of children with cystic fibrosis. Sci. Rep. 2016, 6, 22493. [Google Scholar] [CrossRef]
- Thavamani, A.; Salem, I.; Sferra, T.; Sankararaman, S. Impact of Altered Gut Microbiota and Its Metabolites in Cystic Fibrosis. Metabolites 2021, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.R.; Pope, C.E.; Hayden, H.S.; Heltshe, S.; Levy, R.; McNamara, S.; Jacobs, M.A.; Rohmer, L.; Radey, M.; Ramsey, B.W.; et al. Escherichia coli dysbiosis correlates with gastrointestinal dysfunction in children with cystic fibrosis. Clin. Infect. Dis. 2014, 58, 396–399. [Google Scholar] [CrossRef]
- Meeker, S.M.; Mears, K.S.; Sangwan, N.; Brittnacher, M.J.; Weiss, E.J.; Treuting, P.M.; Tolley, N.; Pope, C.E.; Hager, K.R.; Vo, A.T.; et al. CFTR dysregulation drives active selection of the gut microbiome. PLoS Pathog. 2020, 16, e1008251. [Google Scholar] [CrossRef]
- Matamouros, S.; Hayden, H.S.; Hager, K.R.; Brittnacher, M.J.; Lachance, K.; Weiss, E.J.; Pope, C.E.; Imhaus, A.-F.; McNally, C.P.; Borenstein, E.; et al. Adaptation of commensal proliferating Escherichia coli to the intestinal tract of young children with cystic fibrosis. Proc. Natl. Acad. Sci. USA 2018, 115, 1605–1610. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, M.; Prevaes, S.M.; Kalkman, G.; Tramper-Stranders, G.A.; Hasrat, R.; de Winter-de Groot, K.M.; Janssens, H.M.; Tiddens, H.A.; van Westreenen, M.; Sanders, E.A.; et al. Development of the gut microbiota in early life: The impact of cystic fibrosis and antibiotic treatment. J. Cyst. Fibros. 2020, 19, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Loman, B.R.; Shrestha, C.L.; Thompson, R.; Groner, J.A.; Mejias, A.; Ruoff, K.L.; O’Toole, G.A.; Bailey, M.T.; Kopp, B.T. Age and environmental exposures influence the fecal bacteriome of young children with cystic fibrosis. Pediatr. Pulmonol. 2020, 55, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, M.B.; Moreira, E.A.M.; Tomio, C.; Moreno, Y.M.F.; Daltoé, F.; Barbosa, E.; Neto, N.L.; Buccigrossi, V.; Guarino, A. Altered intestinal microbiota composition, antibiotic therapy and intestinal inflammation in children and adolescents with cystic fibrosis. PLoS ONE 2018, 13, e0198457. [Google Scholar] [CrossRef] [PubMed]
- Duytschaever, G.; Huys, G.; Bekaert, M.; Boulanger, L.; De Boeck, K.; Vandamme, P. Dysbiosis of bifidobacteria and Clostridium cluster XIVa in the cystic fibrosis fecal microbiota. J. Cyst. Fibros. 2013, 12, 206–215. [Google Scholar] [CrossRef]
- Adriaanse, M.P.M.; Van Der Sande, L.J.T.M.; Neucker, A.M.V.D.; Menheere, P.P.C.A.; Dompeling, E.; Buurman, W.A.; Vreugdenhil, A.C.E. Evidence for a Cystic Fibrosis Enteropathy. PLoS ONE 2015, 10, e0138062. [Google Scholar] [CrossRef]
- Ellemunter, H.; Engelhardt, A.; Schüller, K.; Steinkamp, G. Fecal Calprotectin in Cystic Fibrosis and Its Relation to Disease Parameters: A Longitudinal Analysis for 12 Years. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 438–442. [Google Scholar] [CrossRef]
- Enaud, R.; Hooks, K.B.; Barre, A.; Barnetche, T.; Hubert, C.; Massot, M.; Bazin, T.; Clouzeau, H.; Bui, S.; Fayon, M.; et al. Intestinal Inflammation in Children with Cystic Fibrosis Is Associated with Crohn’s-Like Microbiota Disturbances. J. Clin. Med. 2019, 8, 645. [Google Scholar] [CrossRef]
- Flass, T.; Tong, S.; Frank, D.N.; Wagner, B.; Robertson, C.; Kotter, C.V.; Sokol, R.J.; Zemanick, E.; Accurso, F.; Hoffenberg, E.; et al. Intestinal lesions are associated with altered intestinal microbiome and are more frequent in children and young adults with cystic fibrosis and cirrhosis. PLoS ONE 2015, 10, e0116967. [Google Scholar] [CrossRef]
- Parisi, G.F.; Papale, M.; Rotolo, N.; Aloisio, D.; Tardino, L.; Scuderi, M.G.; Di Benedetto, V.; Nenna, R.; Midulla, F.; Leonardi, S. Severe disease in Cystic Fibrosis and fecal calprotectin levels. Immunobiology 2017, 222, 582–586. [Google Scholar] [CrossRef]
- Rumman, N.; Sultan, M.; El-Chammas, K.; Goh, V.; Salzman, N.; Quintero, D.; Werlin, S. Calprotectin in cystic fibrosis. BMC Pediatr. 2014, 14, 133. [Google Scholar] [CrossRef]
- Smyth, R.L.; Croft, N.M.; O’Hea, U.; Marshall, T.G.; Ferguson, A. Intestinal inflammation in cystic fibrosis. Arch. Dis. Child. 2000, 82, 394–399. [Google Scholar] [CrossRef]
- Werlin, S.L.; Benuri-Silbiger, I.; Kerem, E.; Adler, S.N.; Goldin, E.; Zimmerman, J.; Malka, N.; Cohen, L.; Armoni, S.; Yatzkan-Israelit, Y.; et al. Evidence of intestinal inflammation in patients with cystic fibrosis. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Brecelj, J.; Zidar, N.; Jeruc, J.; Orel, R. Morphological and Functional Assessment of Oesophageal Mucosa Integrity in Children With Cystic Fibrosis. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Munck, A. Cystic fibrosis: Evidence for gut inflammation. Int. J. Biochem. Cell Biol. 2014, 52, 180–183. [Google Scholar] [CrossRef]
- Lisle, R.C.D.; Roach, E.; Jansson, K. Effects of laxative and N-acetylcysteine on mucus accumulation, bacterial load, transit, and inflammation in the cystic fibrosis mouse small intestine. Am. J. Physiol. -Gastrointest. Liver Physiol. 2007, 293, G577–G584. [Google Scholar] [CrossRef]
- Vij, N.; Mazur, S.; Zeitlin, P.L. CFTR is a negative regulator of NFkappaB mediated innate immune response. PLoS ONE 2009, 4, e4664. [Google Scholar] [CrossRef]
- Henen, S.; Denton, C.; Teckman, J.; Borowitz, D.; Patel, D. Review of Gastrointestinal Motility in Cystic Fibrosis. J. Cyst. Fibros. 2021, 20, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Debray, M.; El Mourabit, H.; Merabtene, F.; Brot, L.; Ulveling, D.; Chrétien, Y.; Rainteau, M.; Moszer, I.; Wendum, M.; Sokol, H.; et al. Diet-Induced Dysbiosis and Genetic Background Synergize With Cystic Fibrosis Transmembrane Conductance Regulator Deficiency to Promote Cholangiopathy in Mice. Hepatol. Commun. 2018, 2, 1533–1549. [Google Scholar] [CrossRef]
- Kim, K.-A.; Gu, W.; Lee, I.-A.; Joh, E.-H.; Kim, D.-H. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the TLR4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef]
- Crawford, M.; Whisner, C.; Al-Nakkash, L.; Sweazea, K.L. Six-Week High-Fat Diet Alters the Gut Microbiome and Promotes Cecal Inflammation, Endotoxin Production, and Simple Steatosis without Obesity in Male Rats. Lipids 2019, 54, 119–131. [Google Scholar] [CrossRef]
- Knoop, K.A.; McDonald, K.G.; Kulkarni, D.H.; Newberry, R.D. Antibiotics promote inflammation through the translocation of native commensal colonic bacteria. Gut 2016, 65, 1100–1109. [Google Scholar] [CrossRef]
- Debyser, G.; Mesuere, B.; Clement, L.; Van de Weygaert, J.; Van Hecke, P.; Duytschaever, G.; Aerts, M.; Dawyndt, P.; De Boeck, K.; Vandamme, P.; et al. Faecal proteomics: A tool to investigate dysbiosis and inflammation in patients with cystic fibrosis. J. Cyst. Fibros. 2016, 15, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. Role Short-Chain Fat. Acids Gut Microbiota Gut-Brain Communication. Front. Endocrinol. (Lausanne) 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Dengler, F.; Kraetzig, A.; Gabel, G. Butyrate Protects Porcine Colon Epithelium from Hypoxia-Induced Damage on a Functional Level. Nutrients 2021, 13, 305. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, B.G.; Kim, J.H.; Chun, J.; Im, J.P.; Kim, J.S. Sodium butyrate inhibits the NF-kappa B signaling pathway and histone deacetylation, and attenuates experimental colitis in an IL-10 independent manner. Int. Immunopharmacol. 2017, 51, 47–56. [Google Scholar] [CrossRef]
- Yap, Y.A.; McLeod, K.H.; I McKenzie, C.; Gavin, P.G.; Davalos-Salas, M.; Richards, J.L.; Moore, R.J.; Lockett, T.J.; Clarke, J.M.; Eng, V.V.; et al. An acetate-yielding diet imprints an immune and anti-microbial programme against enteric infection. Clin. Transl. Immunol. 2021, 10, e1233. [Google Scholar] [CrossRef] [PubMed]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 4. [Google Scholar] [CrossRef]
- Liu, L.; Sun, D.; Mao, S.; Zhu, W.; Liu, J. Infusion of sodium butyrate promotes rumen papillae growth and enhances expression of genes related to rumen epithelial VFA uptake and metabolism in neonatal twin lambs. J. Anim. Sci. 2019, 97, 909–921. [Google Scholar] [CrossRef]
- Scales, B.S.; Dickson, R.P.; Huffnagle, G.B. A tale of two sites: How inflammation can reshape the microbiomes of the gut and lungs. J. Leukoc. Biol. 2016, 100, 943–950. [Google Scholar] [CrossRef]
- Winter, S.E.; Baumler, A.J. Dysbiosis in the inflamed intestine: Chance favors the prepared microbe. Gut Microbes 2014, 5, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Lupp, C.; Robertson, M.L.; Wickham, M.; Sekirov, I.; Champion, O.L.; Gaynor, E.C.; Finlay, B.B. Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe 2007, 2, 204. [Google Scholar] [CrossRef]
- Corey, M.; McLaughlin, F.; Williams, M.; Levison, H. A comparison of survival, growth, and pulmonary function in patients with cystic fibrosis in Boston and Toronto. J. Clin. Epidemiol. 1988, 41, 583–591. [Google Scholar] [CrossRef]
- Culhane, S.; George, C.; Pearo, B.; Spoede, E. Malnutrition in cystic fibrosis: A review. Nutr. Clin. Pract. 2013, 28, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Gaskin, K.J. Nutritional care in children with cystic fibrosis: Are our patients becoming better? Eur. J. Clin. Nutr. 2013, 67, 558–564. [Google Scholar] [CrossRef]
- Turck, D.; Braegger, C.P.; Colombo, C.; Declercq, D.; Morton, A.; Pancheva, R.; Robberecht, E.; Stern, M.; Strandvik, B.; Wolfe, S.; et al. ESPEN-ESPGHAN-ECFS guidelines on nutrition care for infants, children, and adults with cystic fibrosis. Clin. Nutr. 2016, 35, 557–577. [Google Scholar] [CrossRef] [PubMed]
- Borowitz, D.; Baker, R.D.; Stallings, V. Consensus report on nutrition for pediatric patients with cystic fibrosis. J. Pediatr. Gastroenterol. Nutr. 2002, 35, 246–259. [Google Scholar] [CrossRef]
- Sanders, D.B.; Zhang, Z.; Farrell, P.M.; Lai, H.J. Early life growth patterns persist for 12years and impact pulmonary outcomes in cystic fibrosis. J. Cyst. Fibros. 2018, 17, 528–535. [Google Scholar] [CrossRef]
- Steinkamp, G.; Wiedemann, B. Relationship between nutritional status and lung function in cystic fibrosis: Cross sectional and longitudinal analyses from the German CF quality assurance (CFQA) project. Thorax 2002, 57, 596–601. [Google Scholar] [CrossRef]
- Peterson, M.L.; Jacobs, D.R., Jr.; Milla, C.E. Longitudinal changes in growth parameters are correlated with changes in pulmonary function in children with cystic fibrosis. Pediatrics 2003, 112 Pt 1, 588–592. [Google Scholar] [CrossRef]
- Konstan, M.W.; Butler, S.M.; Wohl, M.E.B.; Stoddard, M.; Matousek, R.; Wagener, J.S.; Johnson, C.A.; Morgan, W.J. Growth and nutritional indexes in early life predict pulmonary function in cystic fibrosis. J. Pediatr. 2003, 142, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Yen, E.H.; Quinton, H.; Borowitz, D. Better nutritional status in early childhood is associated with improved clinical outcomes and survival in patients with cystic fibrosis. J. Pediatr. 2013, 162, 530–535.e1. [Google Scholar] [CrossRef]
- Hanna, R.M.; Weiner, D.J. Overweight and obesity in patients with cystic fibrosis: A center-based analysis. Pediatr. Pulmonol. 2015, 50, 35–41. [Google Scholar] [CrossRef]
- Forrester, D.L.; Knox, A.J.; Smyth, A.R.; Fogarty, A.W. Measures of body habitus are associated with lung function in adults with cystic fibrosis: A population-based study. J. Cyst. Fibros. 2013, 12, 284–289. [Google Scholar] [CrossRef]
- Forte, G.C.; Pereira, J.S.; Drehmer, M.; Simon, M.I.S.D.S. Anthropometric and dietary intake indicators as predictors of pulmonary function in cystic fibrosis patients. J. Bras. Pneumol. 2012, 38, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Pedreira, C.; Robert, R.; Dalton, V.; Oliver, M.; Carlin, J.; Robinson, P.; Cameron, F. Association of body composition and lung function in children with cystic fibrosis. Pediatr. Pulmonol. 2005, 39, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, M.; Nathan, N.; Sarouk, I.; Bar Aluma, B.E.; Dagan, A.; Bezalel, Y.; Keler, S.; Vilozni, D.; Efrati, O. Nutritional Status in Childhood as a Prognostic Factor in Patients with Cystic Fibrosis. Lung 2019, 197, 371–376. [Google Scholar] [CrossRef]
- Stallings, V.A.; Stark, L.J.; Robinson, K.A.; Feranchak, A.P.; Quinton, H. Evidence-based practice recommendations for nutrition-related management of children and adults with cystic fibrosis and pancreatic insufficiency: Results of a systematic review. J. Am. Diet. Assoc. 2008, 108, 832–839. [Google Scholar] [CrossRef]
- Sutherland, R.; Katz, T.; Liu, V.; Quintano, J.; Brunner, R.; Tong, C.W.; Collins, C.E.; Ooi, C.Y. Dietary intake of energy-dense, nutrient-poor and nutrient-dense food sources in children with cystic fibrosis. J. Cyst. Fibros. 2018, 17, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Poulimeneas, D.; Grammatikopoulou, M.G.; Devetzi, P.; Petrocheilou, A.; Kaditis, A.G.; Papamitsou, T.; Doudounakis, S.E.; Vassilakou, T. Adherence to Dietary Recommendations, Nutrient Intake Adequacy and Diet Quality among Pediatric Cystic Fibrosis Patients: Results from the GreeCF Study. Nutrients 2020, 12, 3126. [Google Scholar] [CrossRef]
- Calvo-Lerma, J.; Hulst, J.; Boon, M.; Martins, T.; Ruperto, M.; Colombo, C.; Fornés-Ferrer, V.; Woodcock, S.; Claes, I.; Asseiceira, I.; et al. The Relative Contribution of Food Groups to Macronutrient Intake in Children with Cystic Fibrosis: A European Multicenter Assessment. J. Acad. Nutr. Diet. 2019, 119, 1305–1319. [Google Scholar] [CrossRef] [PubMed]
- Woestenenk, J.; Castelijns, S.; Van Der Ent, C.; Houwen, R. Dietary intake in children and adolescents with cystic fibrosis. Clin. Nutr. 2014, 33, 528–532. [Google Scholar] [CrossRef]
- Smith, C.; Winn, A.; Seddon, P.; Ranganathan, S. A fat lot of good: Balance and trends in fat intake in children with cystic fibrosis. J. Cyst. Fibros. 2012, 11, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulou, P.; Fotoulaki, M.; Nikolaou, A.; Nousia-Arvanitakis, S.; Maria, F. Prevalence of malnutrition and obesity among cystic fibrosis patients. Pediatr. Int. 2014, 56, 89–94. [Google Scholar] [CrossRef]
- Stephenson, A.L.; A Mannik, L.; Walsh, S.; Brotherwood, M.; Robert, R.; Darling, P.B.; Nisenbaum, R.; Moerman, J.; Stanojevic, S. Longitudinal trends in nutritional status and the relation between lung function and BMI in cystic fibrosis: A population-based cohort study. Am. J. Clin. Nutr. 2013, 97, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Gramegna, A.; Aliberti, S.; Contarini, M.; Savi, D.; Sotgiu, G.; Majo, F.; Saderi, L.; Lucidi, V.; Amati, F.; Pappalettera, M.; et al. Overweight and obesity in adults with cystic fibrosis: An Italian multicenter cohort study. J. Cyst. Fibros. 2021; in press. [Google Scholar] [CrossRef]
- Bonhoure, A.; Boudreau, V.; Litvin, M.; Colomba, J.; Bergeron, C.; Mailhot, M.; Tremblay, F.; Lavoie, A.; Rabasa-Lhoret, R. Overweight, obesity and significant weight gain in adult patients with cystic fibrosis association with lung function and cardiometabolic risk factors. Clin. Nutr. 2020, 39, 2910–2916. [Google Scholar] [CrossRef]
- Harindhanavudhi, T.; Wang, Q.; Dunitz, J.; Moran, A.; Moheet, A. Prevalence and factors associated with overweight and obesity in adults with cystic fibrosis: A single-center analysis. J. Cyst. Fibros. 2020, 19, 139–145. [Google Scholar] [CrossRef]
- Bass, R.; Brownell, J.N.; Stallings, V.A. The Impact of Highly Effective CFTR Modulators on Growth and Nutrition Status. Nutrients 2021, 13, 2907. [Google Scholar] [CrossRef]
- Stallings, V.A.; Sainath, N.; Oberle, M.; Bertolaso, C.; Schall, J.I. Energy Balance and Mechanisms of Weight Gain with Ivacaftor Treatment of Cystic Fibrosis Gating Mutations. J. Pediatr. 2018, 201, 229–237.e4. [Google Scholar] [CrossRef]
- Wainwright, C.E.; Elborn, J.S.; Ramsey, B.W. Lumacaftor-Ivacaftor in Patients with Cystic Fibrosis Homozygous for Phe508del CFTR. N. Engl. J. Med. 2015, 373, 1783–1784. [Google Scholar] [CrossRef]
- Rowe, S.M.; Heltshe, S.L.; Gonska, T.; Donaldson, S.H.; Borowitz, D.; Gelfond, D.; Sagel, S.D.; Khan, U.; Mayer-Hamblett, N.; Van Dalfsen, J.M.; et al. Clinical mechanism of the cystic fibrosis transmembrane conductance regulator potentiator ivacaftor in G551D-mediated cystic fibrosis. Am. J. Respir. Crit. Care Med. 2014, 190, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Chi, M.M.; Scull, B.P.; Rigby, R.; Schwerbrock, N.M.J.; Magness, S.; Jobin, C.; Lund, P.K. High-fat diet: Bacteria interactions promote intestinal inflammation which precedes and correlates with obesity and insulin resistance in mouse. PLoS ONE 2010, 5, e12191. [Google Scholar] [CrossRef]
- Gulhane, M.; Murray, L.; Lourie, R.; Tong, H.; Sheng, Y.H.; Wang, R.; Kang, A.; Schreiber, V.; Wong, K.Y.; Magor, G.; et al. High Fat Diets Induce Colonic Epithelial Cell Stress and Inflammation that is Reversed by IL-22. Sci. Rep. 2016, 6, 28990. [Google Scholar] [CrossRef] [PubMed]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front. Cell. Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut-lung axis. Mucosal. Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef]
- Hoen, A.G.; Li, J.; Moulton, L.A.; O’Toole, G.A.; Housman, M.L.; Koestler, D.C.; Guill, M.F.; Moore, J.; Hibberd, P.L.; Morrison, H.; et al. Associations between gut microbial colonization in early life and respiratory outcomes in cystic fibrosis. J. Pediatrics 2015, 167, 138–147.e3. [Google Scholar] [CrossRef]
- Lai, H.-C.; Lin, T.-L.; Chen, T.-W.; Kuo, Y.-L.; Chang, C.-J.; Wu, T.-R.; Shu, C.-C.; Tsai, Y.-H.; Swift, S.; Lu, C.-C. Gut microbiota modulates COPD pathogenesis: Role of anti-inflammatory Parabacteroides goldsteinii lipopolysaccharide. Gut 2022, 71, 309–321. [Google Scholar] [CrossRef]
- Bolia, R.; Ooi, C.Y.; Lewindon, P.; Bishop, J.; Ranganathan, S.; Harrison, J.; Ford, K.; Van Der Haak, N.; Oliver, M.R. Practical approach to the gastrointestinal manifestations of cystic fibrosis. J. Paediatr. Child. Health 2018, 54, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Beaufils, F.; Mas, E.; Mittaine, M.; Addra, M.; Fayon, M.; Delhaes, L.; Clouzeau, H.; Galode, F.; Lamireau, T.; Bui, S.; et al. Increased Fecal Calprotectin Is Associated with Worse Gastrointestinal Symptoms and Quality of Life Scores in Children with Cystic Fibrosis. J. Clin. Med. 2020, 9, 4080. [Google Scholar] [CrossRef] [PubMed]
- Sathe, M.; Huang, R.; Heltshe, S.; Eng, A.; Borenstein, E.; Miller, S.I.; Hoffman, L.; Gelfond, D.; Leung, D.H.; Borowitz, D.; et al. Gastrointestinal Factors Associated With Hospitalization in Infants With Cystic Fibrosis: Results from the BONUS Study. J. Pediatr. Gastroenterol. Nutr. 2021, 73, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.; Komaki, Y.; Komaki, F.; Micic, D.; Zullow, S.; Sakuraba, A. Risk of gastrointestinal cancers in patients with cystic fibrosis: A systematic review and meta-analysis. Lancet Oncol. 2018, 19, 758–767. [Google Scholar] [CrossRef]
- Bernstein, C.N.; Blanchard, J.F.; Kliewer, E.; Wajda, A. Cancer risk in patients with inflammatory bowel disease: A population-based study. Cancer 2001, 91, 854–862. [Google Scholar] [CrossRef]
- Rutter, M.; Saunders, B.; Wilkinson, K.; Rumbles, S.; Schofield, G.; Kamm, M.; Williams, C.; Price, A.; Talbot, I.; Forbes, A. Severity of inflammation is a risk factor for colorectal neoplasia in ulcerative colitis. Gastroenterology 2004, 126, 451–459. [Google Scholar] [CrossRef]
- Ullman, T.A.; Itzkowitz, S.H. Intestinal inflammation and cancer. Gastroenterology 2011, 140, 1807–1816. [Google Scholar] [CrossRef]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef]
- Arthur, J.C.; Perez-Chanona, E.; Mühlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.-J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Pleguezuelos-Manzano, C.; Puschhof, J.; Rosendahl Huber, A.; Van Hoeck, A.; Wood, H.M.; Nomburg, J.; Gurjao, C.; Manders, F.; Dalmasso, G.; Stege, P.B.; et al. Mutational signature in colorectal cancer caused by genotoxic pks(+) E coli. Nature 2020, 580, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.M.; Campbell, B.J.; Hart, C.; Mpofu, C.; Nayar, M.; Singh, R.; Englyst, H.; Williams, H.F.; Rhodes, J.M. Enhanced Escherichia coli adherence and invasion in Crohn’s disease and colon cancer. Gastroenterology 2004, 127, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chen, Y.; Jiang, H.; Robbins, G.T.; Nie, D. G-protein-coupled receptor for short-chain fatty acids suppresses colon cancer. Int. J. Cancer 2011, 128, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Holley, D.; Collins, L.B.; Montgomery, S.A.; Whitmore, A.C.; Hillhouse, A.; Curry, K.P.; Renner, S.W.; Greenwalt, A.; Ryan, E.P.; et al. A gnotobiotic mouse model demonstrates that dietary fiber protects against colorectal tumorigenesis in a microbiota- and butyrate-dependent manner. Cancer Discov. 2014, 4, 1387–1397. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, Q.; Sun, L.; Ye, Y.; Ji, G. Short-chain fatty acids administration is protective in colitis-associated colorectal cancer development. J. Nutr. Biochem. 2018, 57, 103–109. [Google Scholar] [CrossRef]
- Milner, E.; Stevens, B.; An, M.; Lam, V.; Ainsworth, M.; Dihle, P.; Stearns, J.; Dombrowski, A.; Rego, D.; Segars, K. Utilizing Probiotics for the Prevention and Treatment of Gastrointestinal Diseases. Front. Microbiol. 2021, 12, 689958. [Google Scholar] [CrossRef]
- Santo, C.E.; Caseiro, C.; Martins, M.; Monteiro, R.; Brandão, I. Gut microbiota, in the halfway between nutrition and lung function. Nutrients 2021, 13, 1716. [Google Scholar] [CrossRef]
- Araya, M.; Morelli, L.; Reid, G.; Sanders, M.E.; Stanton, C.; Pineiro, M. Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; World Health Organization: London, Ontario; Food and Agriculture Organization of the United Nations: Quebec City, QC, Canada, 2002. [Google Scholar]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Author Correction: Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 642. [Google Scholar] [CrossRef]
- Coffey, M.J.; Garg, M.; Homaira, N.; Jaffé, A.; Ooi, C.Y. Probiotics for people with cystic fibrosis. Cochrane Database Syst. Rev. 2018. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- Arrieta, M.-C.; Stiemsma, L.T.; Amenyogbe, N.; Brown, E.M.; Finlay, B. The Intestinal Microbiome in Early Life: Health and Disease. Front. Immunol. 2014, 5, 427. [Google Scholar] [CrossRef]
- Bergmann, K.R.; Liu, S.X.L.; Tian, R.; Kushnir, A.; Turner, J.R.; Li, H.L.; Chou, P.M.; Weber, C.R.; De Plaen, I.G. Bifidobacteria Stabilize Claudins at Tight Junctions and Prevent Intestinal Barrier Dysfunction in Mouse Necrotizing Enterocolitis. Am. J. Pathol. 2013, 182, 1595–1606. [Google Scholar] [CrossRef]
- Goldenberg, J.Z.; Yap, C.; Lytvyn, L.; Lo, C.K.-F.; Beardsley, J.; Mertz, D.; Johnston, B.C. Probiotics for the prevention of Clostridium difficile-associated diarrhea in adults and children. Cochrane Database Syst. Rev. 2017, 12, Cd006095. [Google Scholar] [CrossRef]
- Deng, K.; Chen, T.; Wu, Q.; Xin, H.; Wei, Q.; Hu, P.; Wang, X.; Wang, X.; Wei, H.; Shah, N.P. In vitro and in vivo examination of anticolonization of pathogens by Lactobacillus paracasei FJ861111.1. J. Dairy Sci. 2015, 98, 6759–6766. [Google Scholar] [CrossRef]
- Su, G.; Ko, C.W.; Bercik, P.; Falck-Ytter, Y.; Sultan, S.; Weizman, A.V.; Morgan, R.L. AGA Clinical Practice Guidelines on the Role of Probiotics in the Management of Gastrointestinal Disorders. Gastroenterology 2020, 159, 697–705. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Maldonado-Gómez, M.X.; Martínez, I.; Bottacini, F.; O’Callaghan, A.; Ventura, M.; van Sinderen, D.; Hillmann, B.; Vangay, P.; Knights, D.; Hutkins, R.W.; et al. Stable Engraftment of Bifidobacterium longum AH1206 in the Human Gut Depends on Individualized Features of the Resident Microbiome. Cell Host Microbe 2016, 20, 515–526. [Google Scholar] [CrossRef]
- Van Baarlen, P.; Wells, J.M.; Kleerebezem, M. Regulation of intestinal homeostasis and immunity with probiotic lactobacilli. Trends Immunol. 2013, 34, 208–215. [Google Scholar] [CrossRef]
- Rivière, A.; Gagnon, M.; Weckx, S.; Roy, D.; De Vuyst, L. Mutual Cross-Feeding Interactions between Bifidobacterium longum subsp. longum NCC2705 and Eubacterium rectale ATCC 33656 Explain the Bifidogenic and Butyrogenic Effects of Arabinoxylan Oligosaccharides. Appl. Environ. Microbiol. 2015, 81, 7767–7781. [Google Scholar] [CrossRef]
- Hegarty, J.W.; Guinane, C.; Ross, R.P.; Hill, C.; Cotter, P.D. Bacteriocin production: A relatively unharnessed probiotic trait? F1000Res 2016, 5, 2587. [Google Scholar] [CrossRef]
- Bali, V.; Panesar, P.S.; Bera, M.B.; Kennedy, J.F. Bacteriocins: Recent Trends and Potential Applications. Crit. Rev. Food Sci. Nutr. 2016, 56, 817–834. [Google Scholar] [CrossRef]
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef]
- Collado, M.C.; Grześkowiak, Ł.; Salminen, S. Probiotic strains and their combination inhibit in vitro adhesion of pathogens to pig intestinal mucosa. Curr. Microbiol. 2007, 55, 260–265. [Google Scholar] [CrossRef]
- Fijan, S.; Šulc, D.; Steyer, A. Study of the In Vitro Antagonistic Activity of Various Single-Strain and Multi-Strain Probiotics against Escherichia coli. Int. J. Environ. Res. Public Health 2018, 15, 1539. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Klaenhammer, T.R.; Kleerebezem, M.; Kopp, M.V.; Rescigno, M. The impact of probiotics and prebiotics on the immune system. Nat. Rev. Immunol. 2012, 12, 728–734. [Google Scholar] [CrossRef]
- Sichetti, M.; De Marco, S.; Pagiotti, R.; Traina, G.; Pietrella, D. Anti-inflammatory effect of multistrain probiotic formulation (L. rhamnosus, B. lactis, and B. longum). Nutrition 2018, 53, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Pothoulakis, C. Review article: Anti-inflammatory mechanisms of action of Saccharomyces boulardii. Aliment Pharmacol. Ther. 2009, 30, 826–833. [Google Scholar] [CrossRef]
- Costeloe, K.; Hardy, P.; Juszczak, E.; Wilks, M.; Millar, M.R. Bifidobacterium breve BBG-001 in very preterm infants: A randomised controlled phase 3 trial. Lancet 2016, 387, 649–660. [Google Scholar] [CrossRef]
- Przemska-Kosicka, A.; Childs, C.E.; Enani, S.; Maidens, C.; Dong, H.; Bin Dayel, I.; Tuohy, K.; Todd, S.; Gosney, M.A.; Yaqoob, P. Effect of a synbiotic on the response to seasonal influenza vaccination is strongly influenced by degree of immunosenescence. Immun. Ageing 2016, 13, 6. [Google Scholar] [CrossRef]
- Vitetta, L.; Vitetta, G.; Hall, S. Immunological Tolerance and Function: Associations Between Intestinal Bacteria, Probiotics, Prebiotics, and Phages. Front. Immunol. 2018, 9, 2240. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 178. [Google Scholar] [CrossRef]
- Sanders, M.E.; Benson, A.; Lebeer, S.; Merenstein, D.J.; Klaenhammer, T.R. Shared mechanisms among probiotic taxa: Implications for general probiotic claims. Curr. Opin. Biotechnol. 2018, 49, 207–216. [Google Scholar] [CrossRef]
- Fredua-Agyeman, M.; Stapleton, P.; Basit, A.W.; Beezer, A.E.; Gaisford, S. In vitro inhibition of Clostridium difficile by commercial probiotics: A microcalorimetric study. International J. Pharm. 2017, 517, 96–103. [Google Scholar] [CrossRef]
- Vieira, A.; Galvão, I.; Macia, L.; Sernaglia, M.; Vinolo, M.A.; Garcia, C.; Tavares, L.P.; Amaral, F.A.; Sousa, L.; Martins, F.; et al. Dietary fiber and the short-chain fatty acid acetate promote resolution of neutrophilic inflammation in a model of gout in mice. J. Leukoc. Biol. 2017, 101, 275–284. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Sci. (N. Y.) 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Salinas, E.; Reyes-Pavón, D.; Cortes-Perez, N.G.; Torres-Maravilla, E.; Bitzer-Quintero, O.K.; Langella, P.; Bermúdez-Humarán, L.G. Bioactive Compounds in Food as a Current Therapeutic Approach to Maintain a Healthy Intestinal Epithelium. Microorganisms 2021, 9, 1634. [Google Scholar] [CrossRef]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef]
- La Fata, G.; Weber, P.; Mohajeri, M.H. Probiotics and the Gut Immune System: Indirect Regulation. Probiotics Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef]
- Kluijfhout, S.; Trieu, T.V.; Vandenplas, Y. Efficacy of the Probiotic Probiotical Confirmed in Acute Gastroenteritis. Pediatr. Gastroenterol. Hepatol. Nutr. 2020, 23, 464–471. [Google Scholar] [CrossRef]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. 1999, 276, G941–G950. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, P.; Zhang, X. Probiotics Regulate Gut Microbiota: An Effective Method to Improve Immunity. Molecules 2021, 26, 6076. [Google Scholar] [CrossRef]
- Mujagic, Z.; De Vos, P.; Boekschoten, M.V.; Govers, C.; Pieters, H.-J.H.M.; De Wit, N.J.W.; Bron, P.A.; Masclee, A.A.M.; Troost, F.J. The effects of Lactobacillus plantarum on small intestinal barrier function and mucosal gene transcription; a randomized double-blind placebo controlled trial. Sci. Rep. 2017, 7, 40128. [Google Scholar] [CrossRef]
- Kim, Y.K.; Shin, C. The Microbiota-Gut-Brain Axis in Neuropsychiatric Disorders: Pathophysiological Mechanisms and Novel Treatments. Curr. Neuropharmacol. 2018, 16, 559–573. [Google Scholar] [CrossRef]
- Reid, G. Disentangling What We Know About Microbes and Mental Health. Front. Endocrinol. 2019, 10, 81. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to live yoghurt cultures and improved lactose digestion (ID 1143, 2976) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1763. [Google Scholar] [CrossRef]
- Bruzzese, E.; Raia, V.; Ruberto, E.; Scotto, R.; Giannattasio, A.; Bruzzese, D.; Cavicchi, M.C.; Francalanci, M.; Colombo, C.; Faelli, N.; et al. Lack of efficacy of Lactobacillus GG in reducing pulmonary exacerbations and hospital admissions in children with cystic fibrosis: A randomised placebo controlled trial. J. Cyst. Fibros. 2018, 17, 375–382. [Google Scholar] [CrossRef]
- Di Benedetto, L.; Raia, V.; Pastore, A.; Albano, F.; Spagnuolo, M.I.; De Vizia, B.; Guarino, A. Lactobacillus casei strain gg as adjunctive treatment to children with cystic fibrosis. J. Pediatric Gastroenterol. Nutr. 1998, 26, 542. [Google Scholar] [CrossRef]
- Del Campo, R.; Garriga, M.; Pérez-Aragón, A.; Guallarte, P.; Lamas, A.; Máiz, L.; Bayón, C.; Roy, G.; Cantón, R.; Zamora, J.; et al. Improvement of digestive health and reduction in proteobacterial populations in the gut microbiota of cystic fibrosis patients using a Lactobacillus reuteri probiotic preparation: A double blind prospective study. J. Cyst. Fibros. 2014, 13, 716–722. [Google Scholar] [CrossRef]
- Van Biervliet, S.; Hauser, B.; Verhulst, S.; Stepman, H.; Delanghe, J.; Warzee, J.-P.; Pot, B.; Vandewiele, T.; Wilschanski, M. Probiotics in cystic fibrosis patients: A double blind crossover placebo controlled study: Pilot study from the ESPGHAN Working Group on Pancreas/CF. Clin. Nutr. ESPEN 2018, 27, 59–65. [Google Scholar] [CrossRef]
- Anderson, J.L.; Miles, C.; Tierney, A.C. Effect of probiotics on respiratory, gastrointestinal and nutritional outcomes in patients with cystic fibrosis: A systematic review. J. Cyst. Fibros. 2017, 16, 186–197. [Google Scholar] [CrossRef]
- Neri, L.C.L.; Taminato, M.; Filho, L.S. Systematic Review of Probiotics for Cystic Fibrosis Patients: Moving Forward. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 394–399. [Google Scholar] [CrossRef]
- Nikniaz, Z.; Nikniaz, L.; Bilan, N.; Somi, M.H.; Faramarzi, E. Does probiotic supplementation affect pulmonary exacerbation and intestinal inflammation in cystic fibrosis: A systematic review of randomized clinical trials. World J. Pediatrics 2017, 13, 307–313. [Google Scholar] [CrossRef]
- Van Biervliet, S.; Declercq, D.; Somerset, S. Clinical effects of probiotics in cystic fibrosis patients: A systematic review. Clin. Nutr. ESPEN 2017, 18, 37–43. [Google Scholar] [CrossRef]
- Ananthan, A.; Balasubramanian, H.; Rao, S.; Patole, S. Probiotic supplementation in children with cystic fibrosis-a systematic review. Eur. J. Pediatr. 2016, 175, 1255–1266. [Google Scholar] [CrossRef]
- Peredo-Lovillo, A.; Romero-Luna, H.E.; Jiménez-Fernández, M. Health promoting microbial metabolites produced by gut microbiota after prebiotics metabolism. Food Res. Int. 2020, 136, 109473. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Rastall, R.A. Prebiotics in foods. Curr. Opin. Biotechnol. 2012, 23, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Hess, J.M.; Gould, T.J.; Slavin, J.L. Prebiotic Dietary Fiber and Gut Health: Comparing the in Vitro Fermentations of Beta-Glucan, Inulin and Xylooligosaccharide. Nutrients 2017, 9, 1361. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef]
- Krumbeck, J.A.; Rasmussen, H.E.; Hutkins, R.W.; Clarke, J.; Shawron, K.; Keshavarzian, A.; Walter, J. Probiotic Bifidobacterium strains and galactooligosaccharides improve intestinal barrier function in obese adults but show no synergism when used together as synbiotics. Microbiome 2018, 6, 121. [Google Scholar] [CrossRef]
- Shoaf, K.; Mulvey, G.L.; Armstrong, G.D.; Hutkins, R.W. Prebiotic galactooligosaccharides reduce adherence of enteropathogenic Escherichia coli to tissue culture cells. Infect. Immun. 2006, 74, 6920–6928. [Google Scholar] [CrossRef] [PubMed]
- Beserra, B.T.S.; Fernandes, R.; do Rosario, V.A.; Mocellin, M.C.; Kuntz, M.G.F.; Trindade, E.B.S.M. A systematic review and meta-analysis of the prebiotics and synbiotics effects on glycaemia, insulin concentrations and lipid parameters in adult patients with overweight or obesity. Clin. Nutr. 2015, 34, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.J.; Guo, J.; Wang, Q.; Wang, L.; Wang, Y.; Zhang, F.; Huang, W.-J.; Zhang, W.; Liu, W.J.; Wang, Y. Probiotics, prebiotics, and synbiotics for the improvement of metabolic profiles in patients with chronic kidney disease: A systematic review and meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2021, 61, 577–598. [Google Scholar] [CrossRef]
- Fernandes, R.; Rosario, V.A.D.; Mocellin, M.C.; Kuntz, M.G.; Trindade, E.B. Effects of inulin-type fructans, galacto-oligosaccharides and related synbiotics on inflammatory markers in adult patients with overweight or obesity: A systematic review. Clin. Nutr. 2017, 36, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Chethan, G.E.; Garkhal, J.; Sircar, S.; Malik, Y.P.S.; Mukherjee, R.; Sahoo, N.R.; Agarwal, R.K.; De, U.K. Immunomodulatory potential of β-glucan as supportive treatment in porcine rotavirus enteritis. Vet. Immunol. Immunopathol. 2017, 191, 36–43. [Google Scholar] [CrossRef]
- Moro, G.; Arslanoglu, S.; Stahl, B.; Jelinek, J.; Wahn, U.; Boehm, G. A mixture of prebiotic oligosaccharides reduces the incidence of atopic dermatitis during the first six months of age. Arch. Dis. Child. 2006, 91, 814–819. [Google Scholar] [CrossRef]
- Sierra, C.; Bernal, M.-J.; Blasco-Alonso, J.; Martínez, R.; Dalmau, J.; Ortuño, I.; Espín, B.; Vasallo, M.-I.; GIL Ortega, D.; Vidal, M.-L.; et al. Prebiotic effect during the first year of life in healthy infants fed formula containing GOS as the only prebiotic: A multicentre, randomised, double-blind and placebo-controlled trial. Eur. J. Nutr. 2015, 54, 89–99. [Google Scholar] [CrossRef]
- Wilms, E.; An, R.; Smolinska, A.; Stevens, Y.; Weseler, A.R.; Elizalde, M.; Drittij, M.-J.; Ioannou, A.; van Schooten, F.J.; Smidt, H.; et al. Galacto-oligosaccharides supplementation in prefrail older and healthy adults increased faecal bifidobacteria, but did not impact immune function and oxidative stress. Clin. Nutr. 2021, 40, 3019–3031. [Google Scholar] [CrossRef]
- Vulevic, J.; Juric, A.; Tzortzis, G.; Gibson, G.R. A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J. Nutr. 2013, 143, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Vulevic, J.; Juric, A.; Walton, G.E.; Claus, S.; Tzortzis, G.; Toward, R.E.; Gibson, G.R. Influence of galacto-oligosaccharide mixture (B-GOS) on gut microbiota, immune parameters and metabonomics in elderly persons. Br. J. Nutr. 2015, 114, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Maretti, C.; Cavallini, G. The association of a probiotic with a prebiotic (Flortec, Bracco) to improve the quality/quantity of spermatozoa in infertile patients with idiopathic oligoasthenoteratospermia: A pilot study. Andrology 2017, 5, 439–444. [Google Scholar] [CrossRef]
- Wolter, M.; Grant, E.T.; Boudaud, M.; Steimle, A.; Pereira, G.V.; Martens, E.C.; Desai, M.S. Leveraging diet to engineer the gut microbiome. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 885–902. [Google Scholar] [CrossRef] [PubMed]
- Grabinger, T.; Garzon, J.F.G.; Hausmann, M.; Geirnaert, A.; Lacroix, C.; Hennet, T. Alleviation of Intestinal Inflammation by Oral Supplementation With 2-Fucosyllactose in Mice. Front. Microbiol. 2019, 10, 1385. [Google Scholar] [CrossRef]
- Tian, M.; Li, D.; Ma, C.; Feng, Y.; Hu, X.; Chen, F. Barley Leaf Insoluble Dietary Fiber Alleviated Dextran Sulfate Sodium-Induced Mice Colitis by Modulating Gut Microbiota. Nutrients 2021, 13, 846. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, S.R.; Britton, G.J.; Contijoch, E.J.; Vennaro, O.H.; Mortha, A.; Colombel, J.-F.; Grinspan, A.; Clemente, J.C.; Merad, M.; Faith, J.J. Interactions Between Diet and the Intestinal Microbiota Alter Intestinal Permeability and Colitis Severity in Mice. Gastroenterology 2018, 154, 1037–1046.e2. [Google Scholar] [CrossRef]
- Chen, H.; Chen, D.; Qin, W.; Liu, Y.; Che, L.; Huang, Z.; Luo, Y.; Zhang, Q.; Lin, D.; Liu, Y.; et al. Wheat bran components modulate intestinal bacteria and gene expression of barrier function relevant proteins in a piglet model. Int. J. Food Sci. Nutr. 2017, 68, 65–72. [Google Scholar] [CrossRef]
- Zhang, T.; Ding, C.; Zhao, M.; Dai, X.; Yang, J.; Li, Y.; Gu, L.; Wei, Y.; Gong, J.; Zhu, W.; et al. Sodium Butyrate Reduces Colitogenic Immunoglobulin A-Coated Bacteria and Modifies the Composition of Microbiota in IL-10 Deficient Mice. Nutrients 2016, 8, 728. [Google Scholar] [CrossRef]
- Valcheva, R.; Koleva, P.; Martínez, I.; Walter, J.; Gänzle, M.G.; Dieleman, L.A. Inulin-type fructans improve active ulcerative colitis associated with microbiota changes and increased short-chain fatty acids levels. Gut Microbes 2019, 10, 334–357. [Google Scholar] [CrossRef]
- Xie, X.; He, Y.; Li, H.; Yu, D.; Na, L.; Sun, T.; Zhang, D.; Shi, X.; Xia, Y.; Jiang, T.; et al. Effects of prebiotics on immunologic indicators and intestinal microbiota structure in perioperative colorectal cancer patients. Nutrition 2019, 61, 132–142. [Google Scholar] [CrossRef]
- Fritsch, J.; Garces, L.; Quintero, M.A.; Pignac-Kobinger, J.; Santander, A.M.; Fernández, I.; Ban, Y.J.; Kwon, D.; Phillips, M.C.; Knight, K.; et al. Low-Fat, High-Fiber Diet Reduces Markers of Inflammation and Dysbiosis and Improves Quality of Life in Patients With Ulcerative Colitis. Clin. Gastroenterol. Hepatol. 2021, 19, 1189–1199.e30. [Google Scholar] [CrossRef] [PubMed]
- Laffin, M.; Perry, T.; Park, H.; Hotte, N.; Fedorak, R.N.; Thiesen, A.; Dicken, B.; Madsen, K.L. Prebiotic Supplementation Following Ileocecal Resection in a Murine Model is Associated With a Loss of Microbial Diversity and Increased Inflammation. Inflamm. Bowel. Dis. 2017, 24, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Palacios, A.; Harding, A.; Menghini, P.; Himmelman, C.; Retuerto, M.; Nickerson, K.P.; Lam, M.; Croniger, C.M.; McLean, M.; Durum, S.K.; et al. The Artificial Sweetener Splenda Promotes Gut Proteobacteria, Dysbiosis, and Myeloperoxidase Reactivity in Crohn’s Disease-Like Ileitis. Inflamm. Bowel. Dis. 2018, 24, 1005–1020. [Google Scholar] [CrossRef]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef] [PubMed]
- Bernard, H.; Desseyn, J.-L.; Bartke, N.; Kleinjans, L.; Stahl, B.; Belzer, C.; Knol, J.; Gottrand, F.; Husson, M.-O. Dietary pectin-derived acidic oligosaccharides improve the pulmonary bacterial clearance of Pseudomonas aeruginosa lung infection in mice by modulating intestinal microbiota and immunity. J. Infect. Dis. 2015, 211, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef]
- Jang, Y.O.; Kim, O.-H.; Kim, S.J.; Lee, S.H.; Yun, S.; Lim, S.E.; Yoo, H.J.; Shin, Y.; Lee, S.W. High-fiber diets attenuate emphysema development via modulation of gut microbiota and metabolism. Sci. Rep. 2021, 11, 7008. [Google Scholar] [CrossRef]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.C.; Johnson, M.A.; Shaw, D.; Spendlove, I.; Vulevic, J.; Sharpe, G.R.; Hunter, K.A. A prebiotic galactooligosaccharide mixture reduces severity of hyperpnoea-induced bronchoconstriction and markers of airway inflammation. Br. J. Nutr. 2016, 116, 798–804. [Google Scholar] [CrossRef]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet–microbiota interactions and personalized nutrition. Nat. Rev. Microbiol. 2019, 17, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Al Khatib, H.; et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef]
- Lawson, M.A.E.; O’Neill, I.J.; Kujawska, M.; Javvadi, S.G.; Wijeyesekera, A.; Flegg, Z.; Chalklen, L.; Hall, L.J. Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J. 2020, 14, 635–648. [Google Scholar] [CrossRef]
- Fanning, S.; Hall, L.J.; Cronin, M.; Zomer, A.; Mac Sharry, J.; Goulding, D.; O’Connell Motherway, M.; Shanahan, F.; Nally, K.; Dougan, G.; et al. Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. USA 2012, 109, 2108–2113. [Google Scholar] [CrossRef]
- Beaumont, M.; Paës, C.; Mussard, E.; Knudsen, C.; Cauquil, L.; Aymard, P.; Barilly, C.; Gabinaud, B.; Zemb, O.; Fourre, S.; et al. Gut microbiota derived metabolites contribute to intestinal barrier maturation at the suckling-to-weaning transition. Gut Microbes 2020, 11, 1268–1286. [Google Scholar] [CrossRef] [PubMed]
- Zegarra-Ruiz, D.F.; El Beidaq, A.; Iñiguez, A.J.; Di Ricco, M.; Manfredo Vieira, S.; Ruff, W.E.; Mubiru, D.; Fine, R.L.; Sterpka, J.; Greiling, T.M.; et al. A Diet-Sensitive Commensal Lactobacillus Strain Mediates TLR7-Dependent Systemic Autoimmunity. Cell Host Microbe 2019, 25, 113–127.e6. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, T.I.; Wang, J.; Karnell, J.L.; Wang, Q.; Wang, S.; Naiman, B.; Gross, P.; Brohawn, P.Z.; Morehouse, C.; Aoyama, J.; et al. Autoimmune manifestations in aged mice arise from early-life immune dysregulation. Sci. Transl. Med. 2016, 8, 361ra137. [Google Scholar] [CrossRef]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, D.; et al. Altering the Intestinal Microbiota during a Critical Developmental Window Has Lasting Metabolic Consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef]
- AL Nabhani, Z.; Dulauroy, S.; Marques, R.; Cousu, C.; Al Bounny, S.; Déjardin, F.; Sparwasser, T.; Bérard, M.; Cerf-Bensussan, N.; Eberl, G. A Weaning Reaction to Microbiota Is Required for Resistance to Immunopathologies in the Adult. Immunity 2019, 50, 1276–1288.e5. [Google Scholar] [CrossRef] [PubMed]
- Kalbermatter, C.; Trigo, N.F.; Christensen, S.; Ganal-Vonarburg, S.C. Maternal Microbiota, Early Life Colonization and Breast Milk Drive Immune Development in the Newborn. Front. Immunol. 2021, 12, 1768. [Google Scholar] [CrossRef] [PubMed]
- Vatanen, T.; Franzosa, E.A.; Schwager, R.; Tripathi, S.; Arthur, T.D.; Vehik, K.; Lernmark, Å.; Hagopian, W.A.; Rewers, M.J.; She, J.-X.; et al. The human gut microbiome in early-onset type 1 diabetes from the TEDDY study. Nature 2018, 562, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of high-fat diet on gut microbiota: A driving force for chronic disease risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515–520. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, N.; Yoon, H.; Nam, R.H.; Lee, D.H. Microbial Changes and Host Response in F344 Rat Colon Depending on Sex and Age Following a High-Fat Diet. Front. Microbiol. 2018, 9, 2236. [Google Scholar] [CrossRef]
- Tomas, J.; Mulet, C.; Saffarian, A.; Cavin, J.-B.; Ducroc, R.; Regnault, B.; Tan, C.K.; Duszka, K.; Burcelin, R.; Wahli, W.; et al. High-fat diet modifies the PPAR-gamma pathway leading to disruption of microbial and physiological ecosystem in murine small intestine. Proc. Natl. Acad. Sci. USA 2016, 113, E5934–E5943. [Google Scholar] [CrossRef]
- Anitha, M.; Reichardt, F.; Tabatabavakili, S.; Nezami, B.G.; Chassaing, B.; Mwangi, S.; Vijay-Kumar, M.; Gewirtz, A.; Srinivasan, S. Intestinal dysbiosis contributes to the delayed gastrointestinal transit in high-fat diet fed mice. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 328–339. [Google Scholar] [CrossRef]
- Thorburn, A.N.; Macia, L.; Mackay, C.R. Diet, metabolites, and "western-lifestyle" inflammatory diseases. Immunity 2014, 40, 833–842. [Google Scholar] [CrossRef]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.l.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef]
- Ruff, W.E.; Greiling, T.M.; Kriegel, M.A. Host–microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 2020, 18, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J.D.; Li, J.V.; Lahti, L.; Ou, J.; Carbonero, F.; Mohammed, K.; Posma, J.M.; Kinross, J.; Wahl, E.; Ruder, E.; et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 2015, 6, 6342. [Google Scholar] [CrossRef]
- Berer, K.; Martínez, I.; Walker, A.; Kunkel, B.; Schmitt-Kopplin, P.; Walter, J.; Krishnamoorthy, G. Dietary non-fermentable fiber prevents autoimmune neurological disease by changing gut metabolic and immune status. Sci. Rep. 2018, 8, 10431. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Chen, H.; Faas, M.M.; De Haan, B.J.; Li, J.; Xiao, P.; Zhang, H.; Diana, J.; De Vos, P.; Sun, J. Specific inulin-type fructan fibers protect against autoimmune diabetes by modulating gut immunity, barrier function, and microbiota homeostasis. Mol. Nutr. Food Res. 2017, 61, 1601006. [Google Scholar] [CrossRef] [PubMed]
- Madan, J.C.; Koestler, D.C.; Stanton, B.A.; Davidson, L.; Moulton, L.A.; Housman, M.L.; Moore, J.H.; Guill, M.F.; Morrison, H.G.; Sogin, M.L.; et al. Serial analysis of the gut and respiratory microbiome in cystic fibrosis in infancy: Interaction between intestinal and respiratory tracts and impact of nutritional exposures. MBio 2012, 3, e00251-e12. [Google Scholar] [CrossRef]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Halnes, I.; Baines, K.J.; Berthon, B.S.; MacDonald-Wicks, L.K.; Gibson, P.G.; Wood, L.G. Soluble Fibre Meal Challenge Reduces Airway Inflammation and Expression of GPR43 and GPR41 in Asthma. Nutrients 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Garg, M.L.; Gibson, P.G. A high-fat challenge increases airway inflammation and impairs bronchodilator recovery in asthma. J. Allergy Clin. Immunol. 2011, 127, 1133–1140. [Google Scholar] [CrossRef]
- Simon, M.I.S.D.S.; Molle, R.D.; Silva, F.M.; Rodrigues, T.W.; Feldmann, M.; Forte, G.C.; Marostica, P.J.C. Antioxidant Micronutrients and Essential Fatty Acids Supplementation on Cystic Fibrosis Outcomes: A Systematic Review. J. Acad. Nutr. Diet 2020, 120, 1016–1033.e1. [Google Scholar] [CrossRef]
- Kanhere, M.; He, J.; Chassaing, B.; Ziegler, T.R.; Alvarez, J.A.; Ivie, E.A.; Hao, L.; Hanfelt, J.; Gewirtz, A.T.; Tangpricha, V. Bolus weekly Vitamin D3 supplementation impacts gut and airway microbiota in adults with cystic fibrosis: A double-blind, randomized, placebo-controlled clinical trial. J. Clin. Endocrinol. Metab. 2018, 103, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J. Probiotic use and antibiotic associated gastrointestinal symptoms in pediatric patients with cystic fibrosis. Pediatric Pulmonol. 2015, 50 (Suppl. 41), 407. [Google Scholar]
- De Simone, C. The Unregulated Probiotic Market. Clin. Gastroenterol. Hepatol. 2019, 17, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S.; et al. Gut-microbiota-targeted diets modulate human immune status. Cell 2021, 184, 4137–4153.e14. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).