
The Steroidal Alkaloid Tomatidine and Tomatidine-Rich Tomato Leaf Extract Suppress the Human Gastric Cancer-Derived 85As2 Cells In Vitro and In Vivo via Modulation of Interferon-Stimulated Genes



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of TRTLE
2.2. Cell Culture
2.3. Cancer Model Induced by Implantation of 85As2 Cells
2.4. Microarray Analysis
2.5. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Analysis
2.6. Cell Proliferation and Cytotoxicity Assays
2.7. Small Interfering RNA (siRNA) Transfection
2.8. Statistical Analysis
3. Results
3.1. HPLC Analysis of TRTLE
3.2. Tomatidine and TRTLE Inhibit Tumor Growth in a Syngeneic Mouse Model
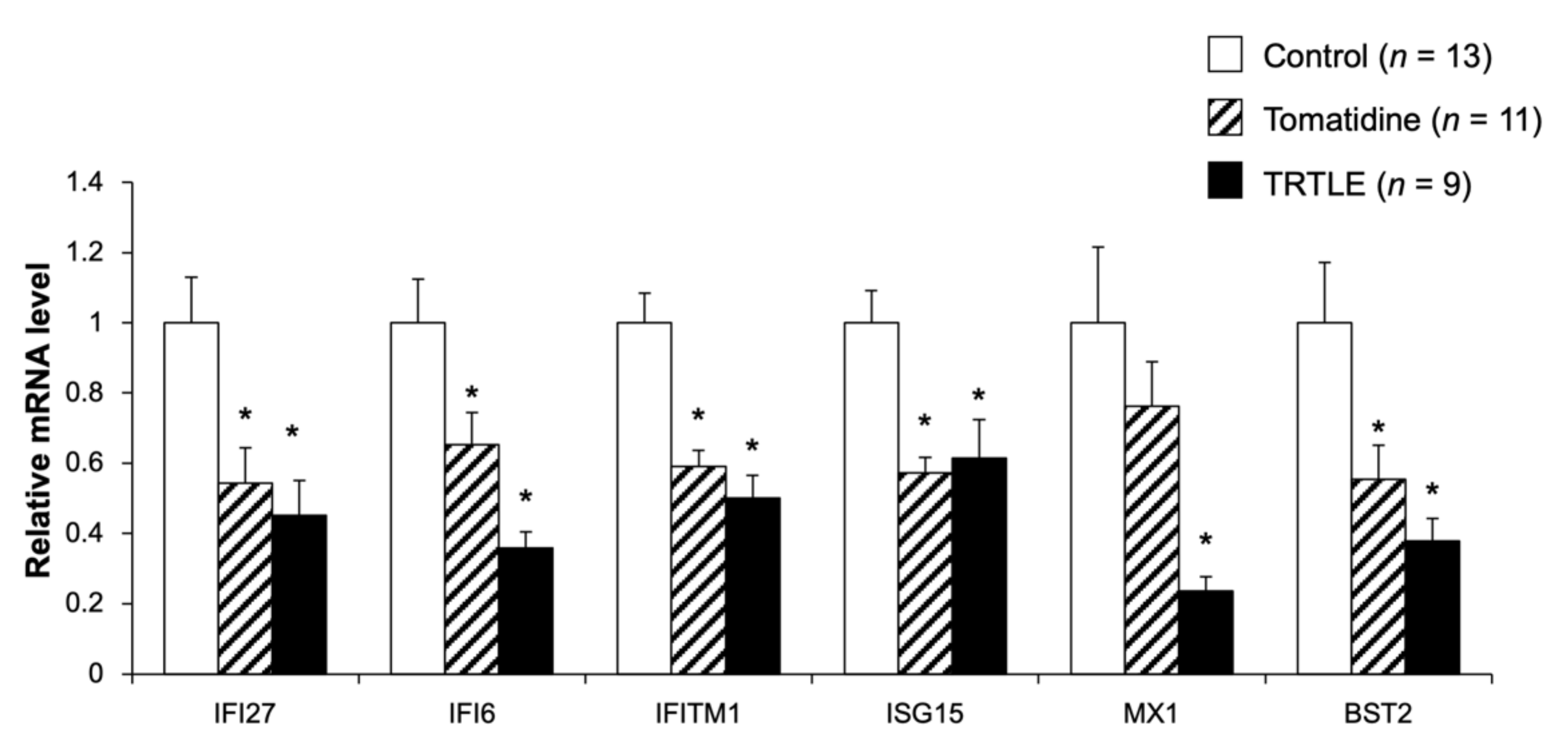
3.3. Tomatidine Suppresses the Expression Levels of Type I Interferon-Stimulated Genes
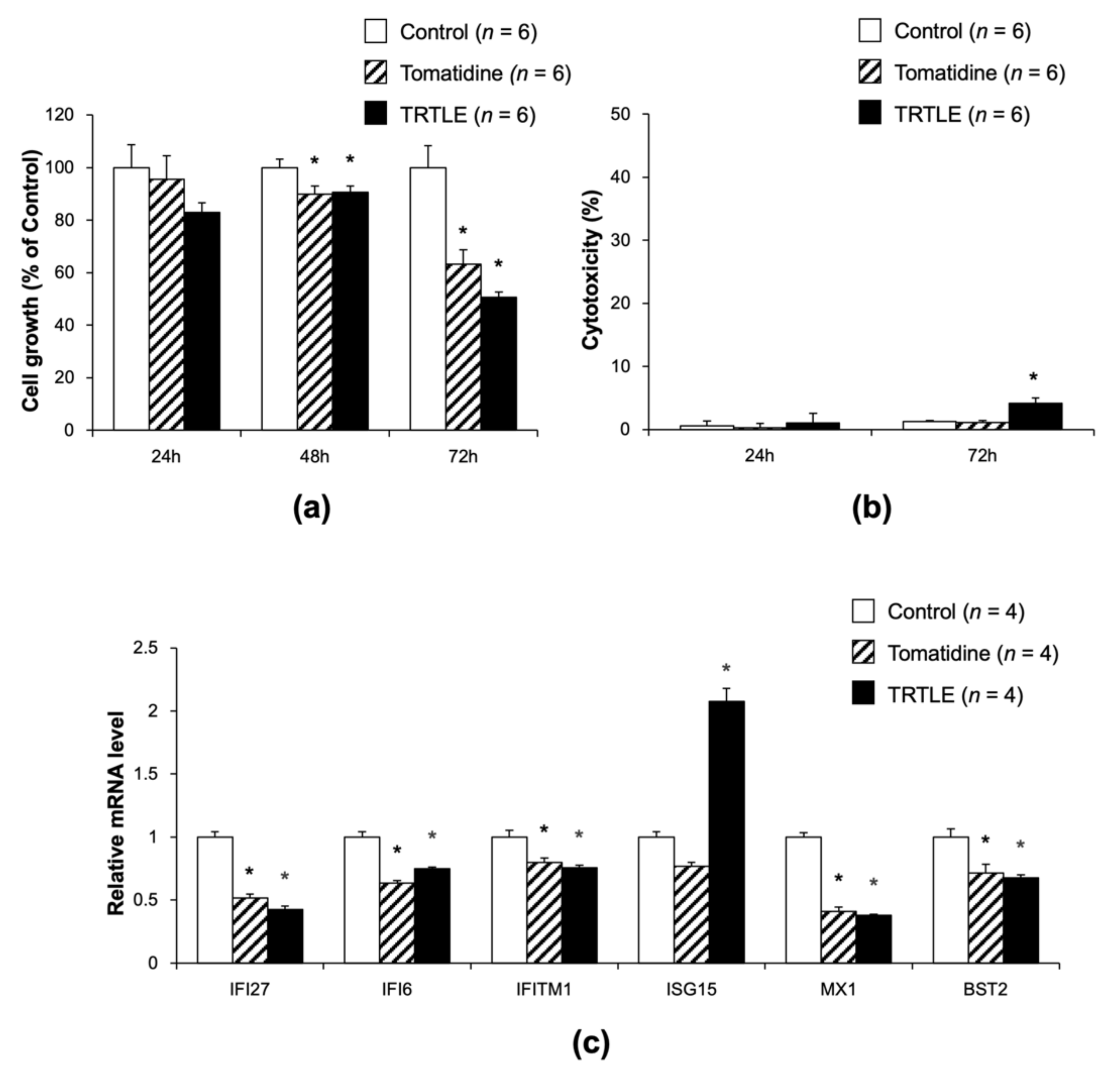
3.4. Tomatidine Inhibits the Proliferation of 85As2 Cells without Cytotoxicity
3.5. Knocking Down of IFI27 Inhibits the Proliferation of 85As2 Cells
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Shikata, K.; Kiyohara, Y.; Kubo, M.; Yonemoto, K.; Ninomiya, T.; Shirota, T.; Tanizaki, Y.; Doi, Y.; Tanaka, K.; Oishi, Y.; et al. A prospective study of dietary salt intake and gastric cancer incidence in a defined Japanese population: The Hisayama study. Int. J. Cancer 2006, 119, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Joossens, J.V.; Hill, M.J.; Elliott, P.; Stamler, R.; Lesaffre, E.; Dyer, A.; Nichols, R.; Kesteloot, H. Dietary salt, nitrate, and stomach cancer mortality in 24 countries. European Cancer Prevention (ECP) and the INTERSALT Cooperative Research Group. Int. J. Epidemiol. 1996, 25, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Bergkvist, L.; Wolk, A. Fruit and vegetable consumption and incidence of gastric cancer: A prospective study. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1998–2001. [Google Scholar] [CrossRef] [Green Version]
- Riboli, E.; Norat, T. Epidemiologic evidence of the protective effect of fruit and vegetables on cancer risk. Am. J. Clin. Nutr. 2003, 78, 559S–569S. [Google Scholar] [CrossRef] [Green Version]
- Bahrami, A.; Ferns, A.G. Effect of curcumin and its derivates on gastric cancer: Molecular mechanisms. Nutr. Cancer 2021, 73, 1553–1569. [Google Scholar] [CrossRef]
- Kim, S.; Kim, W.; Kim, D.H.; Jang, J.H.; Kim, S.J.; Park, S.A.; Hahn, H.; Han, B.W.; Na, H.K.; Chun, K.S.; et al. Resveratrol suppresses gastric cancer cell proliferation and survival through inhibition of PIM-1 kinase activity. Arch. Biochem. Biophys. 2020, 689, 108413. [Google Scholar] [CrossRef]
- Friedman, M. Tomato glycoalkaloids: Role in the plant and in the diet. J. Agric. Food Chem. 2002, 50, 5751–5780. [Google Scholar] [CrossRef]
- Thorne, H.V.; Clarke, G.F.; Skuce, R. The inactivation of herpes simplex virus by some Solanaceae glycoalkaloids. Antivir. Res. 1985, 5, 335–343. [Google Scholar] [CrossRef]
- Zhao, B.; Zhou, B.; Bao, L.; Yang, Y.; Guo, K. Alpha-tomatine exhibits anti-inflammatory activity in lipopolysaccharide-activated macrophages. Inflammation 2015, 38, 1769–1776. [Google Scholar] [CrossRef]
- Huang, H.; Chen, X.; Li, D.; He, Y.; Li, Y.; Du, Z.; Zhang, K.; DiPaola, R.; Goodin, S.; Zheng, X. Combination of α-tomatine and curcumin inhibits growth and induces apoptosis in human prostate cancer cells. PLoS ONE 2015, 10, e0144293. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.W.; Chen, C.H.; Chang, Y.L.; Teng, C.M.; Pan, S.L. α-Tomatine-mediated anti-cancer activity in vitro and in vivo through cell cycle- and caspase-independent pathways. PLoS ONE 2012, 7, e44093. [Google Scholar] [CrossRef]
- Kim, S.P.; Nam, S.H.; Friedman, M. The tomato glycoalkaloid α-tomatine induces caspase-independent cell death in mouse colon cancer CT-26 cells and transplanted tumors in mice. J. Agric. Food Chem. 2015, 63, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Diosa-Toro, M.; Troost, B.; van de Pol, D.; Heberle, A.M.; Urcuqui-Inchima, S.; Thedieck, K.; Smit, J.M. Tomatidine, a novel antiviral compound towards dengue virus. Antivir. Res. 2019, 161, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Chiu, F.L.; Lin, J.K. Tomatidine inhibits iNOS and COX-2 through suppression of NF-κB and JNK pathways in LPS-stimulated mouse macrophages. FEBS Lett. 2008, 582, 2407–2412. [Google Scholar] [CrossRef] [Green Version]
- Koduru, S.; Grierson, D.S.; Van De Venter, M.; Afolayan, A.J. Anticancer activity of steroid alkaloids isolated from Solanum aculeastrum. Pharm. Biol. 2007, 45, 137. [Google Scholar] [CrossRef]
- Chiang, C.T.; Way, T.D.; Tsai, S.J.; Lin, J.K. Diosgenin, a naturally occurring steroid, suppresses fatty acid synthase expression in HER2-overexpressing breast cancer cells through modulating Akt, mTOR and JNK phosphorylation. FEBS Lett. 2007, 581, 5735–5742. [Google Scholar] [CrossRef]
- Choi, S.H.; Ahn, J.B.; Kozukue, N.; Kim, H.J.; Nishitani, Y.; Zhang, L.; Mizuno, M.; Levin, C.E.; Friedman, M. Structure–activity relationships of α-, β1-, γ-, and δ-tomatine and tomatidine against human breast (MDA-MB-231), gastric (KATO-III), and prostate (PC3) cancer cells. J. Agric. Food Chem. 2012, 60, 3891–3899. [Google Scholar] [CrossRef]
- Friedman, M.; Levin, C.E.; Lee, S.U.; Kim, H.J.; Lee, I.S.; Byun, J.O.; Kozukue, N. Tomatine-containing green tomato extracts inhibit growth of human breast, colon, liver, and stomach cancer cells. J. Agric. Food Chem. 2009, 57, 5727–5733. [Google Scholar] [CrossRef]
- Taveira, M.; Ferreres, F.; Gil-Izquierdo, A.; Oliveira, L.; Valentão, P.; Andrade, P.B. Fast determination of bioactive compounds from Lycopersicon esculentum Mill. leaves. Food Chem. 2012, 135, 748–755. [Google Scholar] [CrossRef]
- Yanagihara, K.; Takigahira, M.; Mihara, K.; Kubo, T.; Morimoto, C.; Morita, Y.; Terawaki, K.; Uezono, Y.; Seyama, T. Inhibitory effects of isoflavones on tumor growth and cachexia in newly established cachectic mouse models carrying human stomach cancers. Nutr. Cancer 2013, 65, 578–589. [Google Scholar] [CrossRef]
- Watkins, D.N.; Berman, D.M.; Burkholder, S.G.; Wang, B.; Beachy, P.A.; Baylin, S.B. Hedgehog signalling within airway epithelial progenitors and in small-cell lung cancer. Nature 2003, 422, 313–317. [Google Scholar] [CrossRef]
- Wu, S.J.; Huang, W.C.; Yu, M.C.; Chen, Y.L.; Shen, S.C.; Yeh, K.W.; Liou, C.J. Tomatidine ameliorates obesity-induced nonalcoholic fatty liver disease in mice. J. Nutr. Biochem. 2021, 91, 108602. [Google Scholar] [CrossRef] [PubMed]
- Kusu, H.; Yoshida, H.; Kudo, M.; Okuyama, M.; Harada, N.; Tsuji-Naito, K.; Akagawa, M. Tomatidine reduces palmitate-induced lipid accumulation by activating AMPK via vitamin D receptor-mediated signaling in human HepG2 hepatocytes. Mol. Nutr. Food Res. 2019, 63, e1801377. [Google Scholar] [CrossRef]
- Yamashoji, S.; Onoda, E. Detoxification and function of immature tomato. Food Chem. 2016, 209, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Sucha, L.; Hroch, M.; Rezacova, M.; Rudolf, E.; Havelek, R.; Sispera, L.; Cmielova, J.; Kohlerova, R.; Bezrouk, A.; Tomsik, P. The cytotoxic effect of α-tomatine in MCF-7 human adenocarcinoma breast cancer cells depends on its interaction with cholesterol in incubation media and does not involve apoptosis induction. Oncol. Rep. 2013, 30, 2593–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervantes-Badillo, M.G.; Paredes-Villa, A.; Gómez-Romero, V.; Cervantes-Roldán, R.; Arias-Romero, L.E.; Villamar-Cruz, O.; González-Montiel, M.; Barrios-García, T.; Cabrera-Quintero, A.J.; Rodríguez-Gómez, G.; et al. IFI27/ISG12 downregulates estrogen receptor α transactivation by facilitating its interaction with CRM1/XPO1 in breast cancer cells. Front. Endocrinol. 2020, 11, 568375. [Google Scholar] [CrossRef]
- Wang, H.; Qiu, X.; Lin, S.; Chen, X.; Wang, T.; Liao, T. Knockdown of IFI27 inhibits cell proliferation and invasion in oral squamous cell carcinoma. World J. Surg. Oncol. 2018, 16, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, K.C.; Huang, S.T.; Wu, R.C.; Huang, S.C.; Yeh, T.S.; Chen, M.H.; Hsu, J.T.; Chen, L.W.; Kuo, S.F.; Chueh, H.Y.; et al. Interferon α-inducible protein 27 is an oncogene and highly expressed in cholangiocarcinoma patients with poor survival. Cancer Manag. Res. 2019, 11, 1893–1905. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Gu, S.; Lu, T.; Wu, K.; Li, L.; Dong, C.; Zhou, Y. IFI6 depletion inhibits esophageal squamous cell carcinoma progression through reactive oxygen species accumulation via mitochondrial dysfunction and endoplasmic reticulum stress. J. Exp. Clin. Cancer Res. 2020, 39, 144. [Google Scholar] [CrossRef]
- Yan, J.; Jiang, Y.; Lu, J.; Wu, J.; Zhang, M. Inhibiting of proliferation, migration, and invasion in lung cancer induced by silencing interferon-induced transmembrane protein 1 (IFITM1). BioMed Res. Int. 2019, 2019, 9085435. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Ren, M.; Xu, H.; Xia, H.; Tang, Q.; Liu, M. Inhibition of ISG15 enhances the anti-cancer effect of trametinib in colon cancer cells. OncoTargets Ther. 2019, 12, 10239–10250. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Cao, Y.; Guan, Y.; Zheng, C. BST2 promotes cell proliferation, migration and induces NF-κB activation in gastric cancer. Biotechnol. Lett. 2018, 40, 1015–1027. [Google Scholar] [CrossRef]
- Kim, Y.S.; Do Hwan, J.; Bae, S.; Bae, D.H.; Ahn Shick, W. Identification of differentially expressed genes using an annealing control primer system in stage III serous ovarian carcinoma. BMC Cancer 2010, 10, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Xie, Y.; Zhang, W.; Gao, J.; Wang, M.; Zheng, G.; Yin, X.; Xia, H.; Tao, X. Interferon alpha-inducible protein 27 promotes epithelial–mesenchymal transition and induces ovarian tumorigenicity and stemness. J. Surg. Res. 2015, 193, 255–264. [Google Scholar] [CrossRef]
- Budhu, A.; Chen, Y.; Kim, J.W.; Forgues, M.; Valerie, K.; Harris, C.C.; Wang, X.W. Induction of a unique gene expression profile in primary human hepatocytes by hepatitis C virus core, NS3 and NS5A proteins. Carcinogenesis 2007, 28, 1552–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, U.B.; Wolf, C.; Mattel, M.G.; Chenard, M.P.; Bellocq, J.P. Identification of a new interferon-α-inducible gene (p27) on human chromosome 14q32 and its expression in breast carcinoma. Cancer Res. 1993, 53, 4096–4101. [Google Scholar]
- Lao, M.; Zhang, X.; Ma, T.; Xu, J.; Yang, H.; Duan, Y.; Ying, H.; Zhang, X.; Guo, C.; Qiu, J.; et al. Regulator of calcineurin 1 gene isoform 4 in pancreatic ductal adenocarcinoma regulates the progression of tumor cells. Oncogene 2021, 40, 3136–3151. [Google Scholar] [CrossRef]
- Huang, S.; Zhao, J.; Song, J.; Li, Y.; Zuo, R.; Sa, Y.; Ma, Z.; OuYang, H. Interferon alpha-inducible protein 27 (IFI27) is a prognostic marker for pancreatic cancer based on comprehensive bioinformatics analysis. Bioengineered 2021, 12, 8515–8528. [Google Scholar] [CrossRef]
- Liu, N.; Zuo, C.; Wang, X.; Chen, T.; Yang, D.; Wang, J.; Zhu, H. miR-942 decreases TRAIL-induced apoptosis through ISG12a downregulation and is regulated by AKT. Oncotarget 2014, 5, 4959–4971. [Google Scholar] [CrossRef] [Green Version]
- Erdal, E.; Haider, S.; Rehwinkel, J.; Harris, A.L.; McHugh, P.J. A prosurvival DNA damage-induced cytoplasmic interferon response is mediated by end resection factors and is limited by Trex1. Genes Dev. 2017, 31, 353–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Liang, H.; Fu, S.; Weichselbaum, R.R.; Fu, Y.X. From DNA damage to nucleic acid sensing: A strategy to enhance radiation therapy. Clin. Cancer Res. 2016, 22, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheon, H.J.; Holvey-Bates, E.G.; McGrail, D.J.; Stark, G.R. PD-L1 sustains chronic, cancer cell-intrinsic responses to type I interferon, enhancing resistance to DNA damage. Proc. Natl. Acad. Sci. USA 2021, 118, e2112258118. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPLC | |
|---|---|
| Column | CAPCELL PAK C18-ACR column 4.6 × 250 mm (OSAKA SODA, Osaka, Japan) |
| Guard column | CAPCELL PAK C18-ACR guard column 4.0 × 10 mm (OSAKA SODA) |
| Wavelength | 205 nm |
| Mobile phase A | 25 mM Triethylammonium phosphate |
| Mobile phase B | Acetonitrile |
| Gradient | B% = 20–45–55–57–20 (0–12, 12–17, 17–20, 20–21, 21 min) |
| Column temperature | 25 °C |
| Flow rate | 0.8 mL/min |
| Ingredients | Control | Tomatidine | TRTLE |
|---|---|---|---|
| g/100 g (Except Water) | |||
| Tomatidine (hydrochloride) | - | 0.05 | - |
| Tomatidine-rich tomato leaf extract (TRTLE) | - | - | 0.077 |
| L(-)-Cystine (Wako) | 0.30 | 0.30 | 0.30 |
| AIN93 Vitamin mix (Without choline bitartrate) (Oriental Yeast, Tokyo, Japan) | 1.00 | 1.00 | 1.00 |
| AIN93 Mineral mix (Oriental Yeast) | 3.50 | 3.50 | 3.50 |
| Cellulose (Oriental Yeast) | 5.00 | 5.00 | 5.00 |
| Casein (Oriental Yeast) | 20.00 | 20.00 | 20.00 |
| α-Starch (Oriental Yeast) | 66.20 | 66.20 | 66.20 |
| Safflower oil (Benihana Olein Ichiban Shibori) (Benibana foods, Tokyo, Japan) | 4.00 | 4.00 | 4.00 |
| Gene | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| β-actin | TGGCACCCAGCACAATGA | CTAAGTCATAGTCCGCCTAGAAGCA |
| GAPDH | TGGACCTGACCTGCCGTCTAG | GTGGGTGTCGCTGTTGAAGTC |
| IFI27 | TGCTCTCACCTCATCAGCAGT | CACAACTCCTCCAATCACAACT |
| IFI6 | GATGAGCTGGTCTGCGATCC | TCGAGATACTTGTGGGTGGC |
| IFITM1 | TCGCCTACTCCGTGAAGTCTA | TGTCACAGAGCCGAATACCAG |
| ISG15 | TGTCCCTGAGCAGCTCCATG | TGTCCTGCAGCGCCACACC |
| MX1 | GCCAGGACCAGGTATACAG | GCCTGCGTCAGCCGTGC |
| BST2 | GAGCTTGAGGGAGAGATCACTAC | ATTCTCACGCTTAAGACCTGGTT |
| siRNA | Sense (5′–3′) | Antisense (5′–3′) |
|---|---|---|
| GFP | GCAGCACGACUUCUUCAAGdTdT | CUUGAAGAAGUCGUGCUGCdTdT |
| IFI27 #1 | GUGAAAUAUACCAAAUUCUdTdT | AGAAUUUGGUAUAUUUCACdCdC |
| IFI27 #2 | GAAAUAAAGAUGAAUUGUUdTdT | AACAAUUCAUCUUUAUUUCdTdT |
| Biological Process (Gene Ontology) | |||
|---|---|---|---|
| GO-Term | Description | Count in Network | p-Value |
| GO:0060337 | Type I interferon signaling pathway | 6 of 67 | 0.0062 |
| GO:0045069 | Regulation of viral genome replication | 6 of 99 | 0.0114 |
| GO:1903900 | Regulation of viral life cycle | 7 of 153 | 0.0114 |
| GO:0045071 | Negative regulation of viral genome replication | 5 of 61 | 0.0154 |
| Biological Process (Gene Ontology) | |||
|---|---|---|---|
| GO-Term | Description | Count in Network | p-Value |
| GO:0040029 | Regulation of gene expression, epigenetic | 12 of 202 | 4.63 × 10−6 |
| GO:0060968 | Regulation of gene silencing | 10 of 137 | 1.12 × 10−5 |
| GO:0045814 | Negative regulation of gene expression, epigenetic | 9 of 103 | 1.28 × 10−5 |
| GO:0097549 | Chromatin organization involved in negative regulation of transcription | 9 of 108 | 1.42 × 10−5 |
| GO:0006334 | Nucleosome assembly | 9 of 135 | 5.89 × 10−5 |
| GO:0045653 | Negative regulation of megakaryocyte differentiation | 5 of 18 | 0.00012 |
| GO:0060337 | Type I interferon signaling pathway | 7 of 67 | 0.00012 |
| GO:0006323 | DNA packaging | 10 of 215 | 0.00012 |
| GO:0051253 | Negative regulation of RNA metabolic process | 23 of 1422 | 0.00012 |
| GO:0051172 | Negative regulation of nitrogen compound metabolic process | 31 of 2429 | 0.00012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujimaki, J.; Sayama, N.; Shiotani, S.; Suzuki, T.; Nonaka, M.; Uezono, Y.; Oyabu, M.; Kamei, Y.; Nukaya, H.; Wakabayashi, K.; et al. The Steroidal Alkaloid Tomatidine and Tomatidine-Rich Tomato Leaf Extract Suppress the Human Gastric Cancer-Derived 85As2 Cells In Vitro and In Vivo via Modulation of Interferon-Stimulated Genes. Nutrients 2022, 14, 1023. https://doi.org/10.3390/nu14051023
Fujimaki J, Sayama N, Shiotani S, Suzuki T, Nonaka M, Uezono Y, Oyabu M, Kamei Y, Nukaya H, Wakabayashi K, et al. The Steroidal Alkaloid Tomatidine and Tomatidine-Rich Tomato Leaf Extract Suppress the Human Gastric Cancer-Derived 85As2 Cells In Vitro and In Vivo via Modulation of Interferon-Stimulated Genes. Nutrients. 2022; 14(5):1023. https://doi.org/10.3390/nu14051023
Chicago/Turabian StyleFujimaki, Junya, Neo Sayama, Shigenobu Shiotani, Takanori Suzuki, Miki Nonaka, Yasuhito Uezono, Mamoru Oyabu, Yasutomi Kamei, Haruo Nukaya, Keiji Wakabayashi, and et al. 2022. "The Steroidal Alkaloid Tomatidine and Tomatidine-Rich Tomato Leaf Extract Suppress the Human Gastric Cancer-Derived 85As2 Cells In Vitro and In Vivo via Modulation of Interferon-Stimulated Genes" Nutrients 14, no. 5: 1023. https://doi.org/10.3390/nu14051023
APA StyleFujimaki, J., Sayama, N., Shiotani, S., Suzuki, T., Nonaka, M., Uezono, Y., Oyabu, M., Kamei, Y., Nukaya, H., Wakabayashi, K., Morita, A., Sato, T., & Miura, S. (2022). The Steroidal Alkaloid Tomatidine and Tomatidine-Rich Tomato Leaf Extract Suppress the Human Gastric Cancer-Derived 85As2 Cells In Vitro and In Vivo via Modulation of Interferon-Stimulated Genes. Nutrients, 14(5), 1023. https://doi.org/10.3390/nu14051023