
Neohesperidin Dihydrochalcone and Neohesperidin Dihydrochalcone-O-Glycoside Attenuate Subcutaneous Fat and Lipid Accumulation by Regulating PI3K/AKT/mTOR Pathway In Vivo and In Vitro


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Studies
2.3. Oral Glucose Tolerance Test
2.4. Biochemical Analysis
2.5. Histological Analysis
2.6. Quantitative Real-Time PCR
2.7. Cell Culture
2.8. Oil Red O Staining
2.9. Cell Viability Assay
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
3.1. Effects of NHDC and GNHDC on Body Weight, Tissue Weight, Lipid Profile, and Cytokines in db/db Mice
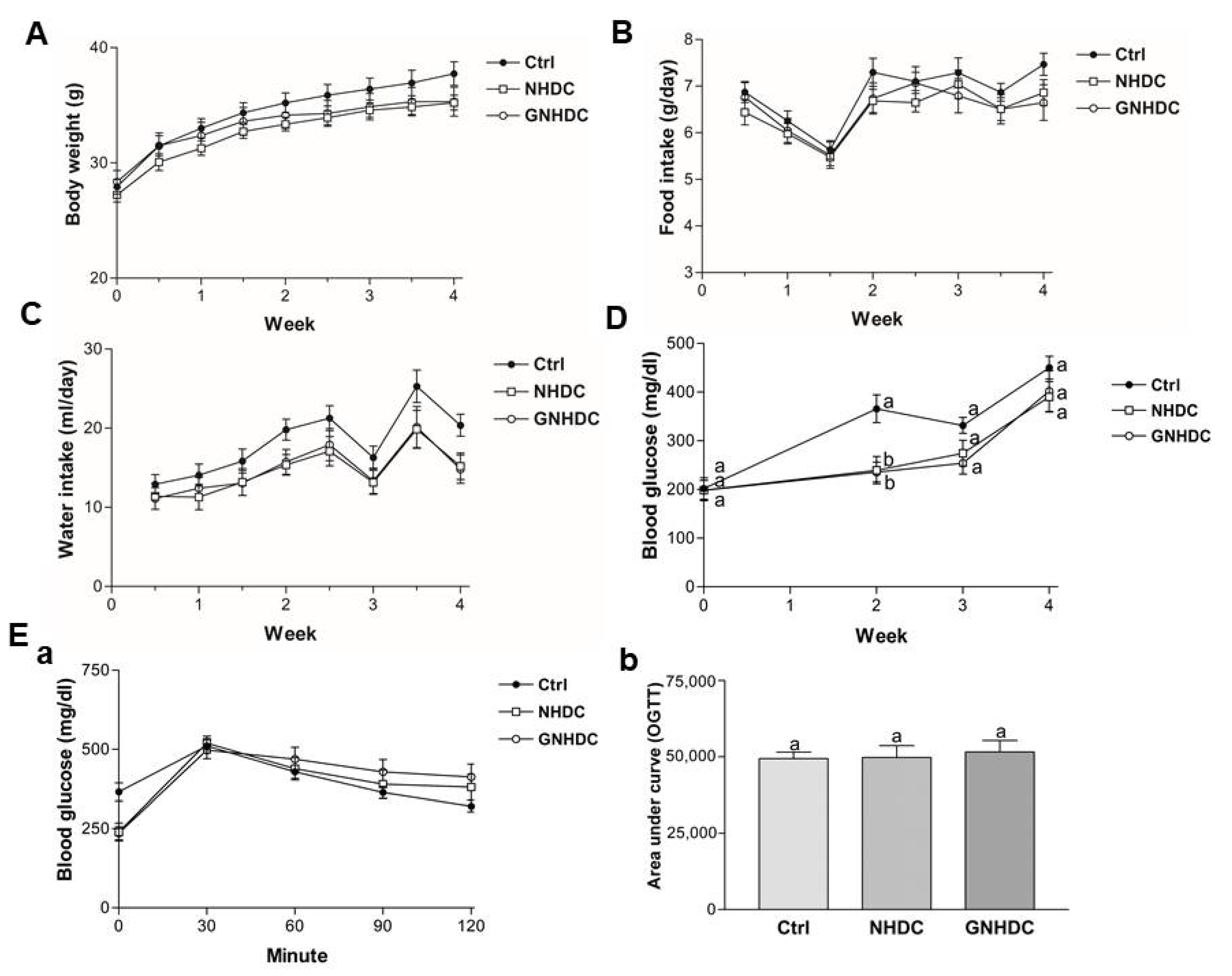
3.2. Effects of NHDC and GNHDC on Body Weight, Food Intake, Water Intake, Fasting Blood Glucose Levels, and OGTT in db/db Mice
3.3. Effects of NHDC and GNHDC on the Subcutaneous Adipocyte Area and the Gene Expressions for Lipid Metabolism in Subcutaneous Adipose Tissues
3.4. Effects of NHDC and GNHDC on Lipid Accumulation and Cell Viability in 3T3-L1 Cells
3.5. Effects of NHDC and GNHDC on Lipogenesis, Adipogenesis, and Proinflammatory Cytokines in 3T3-L1 Cells
3.6. Effects of NHDC and GNHDC on the PI3K/AKT/mTOR Pathway and AMPK in 3T3-L1 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight/ (accessed on 16 December 2020).
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic; World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- Chan, J.M.; Rimm, E.B.; Colditz, G.A.; Stampfer, M.J.; Willett, W.C. Obesity, fat distribution, and weight gain as risk factors for clinical diabetes in men. Diabetes Care 1994, 17, 961–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prospective Studies Collaboration. Body-mass index and cause-specific mortality in 900,000 adults: Collaborative analyses of 57 prospective studies. Lancet 2009, 373, 1083–1096. [Google Scholar] [CrossRef] [Green Version]
- Hamer, M.; Stamatakis, E. Metabolically Healthy Obesity and Risk of All-Cause and Cardiovascular Disease Mortality. J. Clin. Endocrinol. Metab. 2012, 97, 2482–2488. [Google Scholar] [CrossRef] [PubMed]
- Sowers, J.R. Obesity as a cardiovascular risk factor. Am. J. Med. 2003, 115, 37–41. [Google Scholar] [CrossRef]
- El Hafidi, M.; Buelna-Chontal, M.; Sánchez-Muñoz, F.; Carbó, R. Adipogenesis: A Necessary but Harmful Strategy. Int. J. Mol. Sci. 2019, 20, 3657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.M.; Bennett, M.J. Disorders of mitochondrial fatty acid β-oxidation. In Biomarkers in Inborn Errors of Metabolism, 1st ed.; Garg, U., Smith, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 87–93. [Google Scholar] [CrossRef]
- Bartelt, A.; Heeren, J. Adipose tissue browning and metabolic health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [Green Version]
- Wittekind, A.; Walton, J. Worldwide trends in dietary sugars intake. Nutr. Res. Rev. 2014, 27, 330–345. [Google Scholar] [CrossRef] [Green Version]
- Duffey, K.J.; Popkin, B.M. Shifts in Patterns and Consumption of Beverages Between 1965 and 2002. Obesity 2007, 15, 2739–2747. [Google Scholar] [CrossRef]
- Bray, G.A.; Nielsen, S.J.; Popkin, B.M. Consumption of high-fructose corn syrup in beverages may play a role in the epidemic of obesity. Am. J. Clin. Nutr. 2004, 79, 537–543. [Google Scholar] [CrossRef]
- World Health Organization. Executive summary. In Guideline: Sugars Intake for Adults and Children; World Health Organization: Geneva, Switzerland, 2015; pp. 3–7. [Google Scholar]
- Keller, A.; Della Torre, S.B. Sugar-Sweetened Beverages and Obesity among Children and Adolescents: A Review of Systematic Literature Reviews. Child. Obes. 2015, 11, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Malik, V.S.; Pan, A.; Willett, W.C.; Hu, F.B. Sugar-sweetened beverages and weight gain in children and adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2013, 98, 1084–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glória, M. Sweeteners/Others. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Cambridge, UK, 2003; pp. 5696–5699. [Google Scholar]
- Edwards, C.H.; Rossi, M.; Corpe, C.P.; Butterworth, P.J.; Ellis, P.R. The role of sugars and sweeteners in food, diet and health: Alternatives for the future. Trends Food Sci. Technol. 2016, 56, 158–166. [Google Scholar] [CrossRef]
- Food and Drug Administration. GRAS Notice (GRN) No. 902: Neohesperidin Dihydrochalcone; HealthTech BioActives, S.L.U.: Beniel, Spain, 2019; pp. 9–42. [Google Scholar]
- Horowitz, R.M.; Gentili, B. Dihydrochalcone Derivatives and Their Use as Sweetening Agents. U.S. Patent 3,087,821, 30 April 1963. [Google Scholar]
- Hattori, S.; Shimokoriyama, M.; Kanao, M. Studies on Flavanone Glycosides. IV. The Glycosides of Ripe Fruit Peel and Flower Petals of Citrus Aurantium L. J. Am. Chem. Soc. 1952, 74, 3614–3615. [Google Scholar] [CrossRef]
- Horowitz, R.M.; Gentili, B. Taste and structure in phenolic glycosides. J. Agric. Food Chem. 1969, 17, 696–700. [Google Scholar] [CrossRef]
- Harborne, J.B. Biochemistry of phenolic compounds. In Biochemistry of Phenolic Compounds; Academic Press: New York, NY, USA, 1964; pp. 545–571. [Google Scholar]
- O’Brien-Nabors, L. Alternative Sweeteners, 3rd ed.; Marcel Dekker: New York, NY, USA, 2001; pp. 87–96. [Google Scholar] [CrossRef]
- Han, G.E.; Kang, H.-T.; Chung, S.; Lim, C.; Linton, J.A.; Lee, J.-H.; Kim, W.; Kim, S.-H.; Lee, J.H. Novel Neohesperidin Dihydrochalcone Analogue Inhibits Adipogenic Differentiation of Human Adipose-Derived Stem Cells through the Nrf2 Pathway. Int. J. Mol. Sci. 2018, 19, 2215. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Song, X.; Fu, J.; Su, C.; Xia, X.; Song, E.; Song, Y. Artificial sweetener neohesperidin dihydrochalcone showed antioxidative, anti-inflammatory and anti-apoptosis effects against paraquat-induced liver injury in mice. Int. Immunopharmacol. 2015, 29, 722–729. [Google Scholar] [CrossRef]
- Singh, R.G. Chemotaxonomy of Medicinal Plants: Possibilities and Limitations. In Natural Products and Drug Discovery, 1st ed.; Mandal, S., Mandal, V., Konishi, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; p. 127. [Google Scholar] [CrossRef]
- Yang, R.-M.; Liu, F.; He, Z.-D.; Ji, M.; Chu, X.-X.; Kang, Z.-Y.; Cai, D.-Y.; Gao, N.-N. Anti-obesity effect of total phenylpropanoid glycosides from Ligustrum robustum Blume in fatty diet-fed mice via up-regulating leptin. J. Ethnopharmacol. 2015, 169, 459–465. [Google Scholar] [CrossRef]
- Zang, Y.; Zhang, L.; Igarashi, K.; Yu, C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015, 6, 834–841. [Google Scholar] [CrossRef]
- Kerru, N.; Singh-Pillay, A.; Awolade, P.; Singh, P. Current anti-diabetic agents and their molecular targets: A review. Eur. J. Med. Chem. 2018, 152, 436–488. [Google Scholar] [CrossRef]
- Choi, J.-M.; Yoon, B.-S.; Lee, S.-K.; Hwang, J.-K.; Ryang, R. Antioxidant Properties of Neohesperidin Dihydrochalcone: Inhibition of Hypochlorous Acid-Induced DNA Strand Breakage, Protein Degradation, and Cell Death. Biol. Pharm. Bull. 2007, 30, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolivalt, C.; Lee, C.; Beiswenger, K.; Smith, J.; Orlov, M.; Torrance, M.; Masliah, E. Defective insulin signaling pathway and increased glycogen synthase kinase-3 activity in the brain of diabetic mice: Parallels with Alzheimer’s disease and correction by insulin. J. Neurosci. Res. 2008, 86, 3265–3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Forte, T.M.; Taniguchi, S.; Ishida, B.Y.; Oka, K.; Chan, L. The db/db mouse, a model for diabetic dyslipidemia: Molecular characterization and effects of western diet feeding. Metabolism 2000, 49, 22–31. [Google Scholar] [CrossRef]
- Herbreteau, V.; Jittapalapong, S.; Rerkamnuaychoke, W.; Chaval, Y.; Cosson, J.-F.; Morand, S. Protocols for Field and Laboratory Rodent Studies; Kasetsart University: Bangkok, Thailand, 2011; pp. 5–46. [Google Scholar]
- Bagchi, D.P.; MacDougald, O.A. Identification and Dissection of Diverse Mouse Adipose Depots. J. Vis. Exp. 2019, 149. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.; Rudenski, A.; Naylor, B.; Treacher, D.; Turner, R. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Li, L.; Xu, D.; Xia, X.; Pi, R.; Xu, D.; Wang, W.; Du, H.; Song, E.; Song, Y. Protective effects of neohesperidin dihydrochalcone against carbon tetrachloride-induced oxidative damage in vivo and in vitro. Chem. Biol. Interact. 2014, 213, 51–59. [Google Scholar] [CrossRef]
- Suarez, J.; Herrera, M.; Marhuenda, E. Hesperidin and neohesperidin dihydrochalcone on different experimental models of induced gastric ulcer. Phytother. Res. 1996, 10, 616–618. [Google Scholar] [CrossRef]
- Xia, X.; Fu, J.; Song, X.; Shi, Q.; Su, C.; Song, E.; Song, Y. Neohesperidin dihydrochalcone down-regulates MyD88-dependent and -independent signaling by inhibiting endotoxin-induced trafficking of TLR4 to lipid rafts. Free Radic. Biol. Med. 2015, 89, 522–532. [Google Scholar] [CrossRef]
- Lina, B.; der Meulen, H.D.-V.; Leegwater, D. Subchronic (13-week) oral toxicity of neohesperidin dihydrochalcone in rats. Food Chem. Toxicol. 1990, 28, 507–513. [Google Scholar] [CrossRef]
- Henning, S.J. Plasma concentrations of total and free corticosterone during development in the rat. Am. J. Physiol. Metab. 1978, 235, E451–E456. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2005, 28, S5–S10. [Google Scholar]
- Kim, E.; Shin, J.-H.; Seok, P.R.; Kim, M.-S.; Yoo, S.-H.; Kim, Y. Phyllodulcin, a natural functional sweetener, improves diabetic metabolic changes by regulating hepatic lipogenesis, inflammation, oxidative stress, fibrosis, and gluconeogenesis in db/db mice. J. Funct. Foods 2018, 42, 1–11. [Google Scholar] [CrossRef]
- Lim, S.-m.; Kim, E.; Shin, J.-H.; Seok, P.R.; Jung, S.; Yoo, S.-H.; Kim, Y. Xylobiose prevents high-fat diet induced mice obesity by suppressing mesenteric fat deposition and metabolic dysregulation. Molecules 2018, 23, 705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pałasz, A.; Cież, D.; Trzewik, B.; Miszczak, K.; Tynor, G.; Bazan, B. In the search of Glycoside-Based molecules as antidiabetic agents. Top. Curr. Chem. 2019, 377, 19. [Google Scholar] [CrossRef] [Green Version]
- Zang, Y.; Igarashi, K.; Li, Y. Anti-diabetic effects of luteolin and luteolin-7-O-glucoside on KK-A y mice. Biosci. Biotechnol. Biochem. 2016, 80, 1580–1586. [Google Scholar] [CrossRef] [Green Version]
- Wood, I.S.; Trayhurn, P. Glucose transporters (GLUT and SGLT): Expanded families of sugar transport proteins. Br. J. Nutr. 2003, 89, 3–9. [Google Scholar] [CrossRef]
- Kuo, G.-H.; Gaul, M.D.; Liang, Y.; Xu, J.Z.; Du, F.; Hornby, P.; Xu, G.; Qi, J.; Wallace, N.; Lee, S. Synthesis and biological evaluation of benzocyclobutane-C-glycosides as potent and orally active SGLT1/SGLT2 dual inhibitors. Bioorg. Med. Chem. Lett. 2018, 28, 1182–1187. [Google Scholar] [CrossRef]
- Kuroda, S.; Kobashi, Y.; Oi, T.; Kawabe, K.; Shiozawa, F.; Okumura-Kitajima, L.; Sugisaki-Kitano, M.; Io, F.; Yamamoto, K.; Kakinuma, H. Discovery of potent, low-absorbable sodium-dependent glucose cotransporter 1 (SGLT1) inhibitor SGL5213 for type 2 diabetes treatment. Bioorg. Med. Chem. 2019, 27, 394–409. [Google Scholar] [CrossRef]
- Miller, P.E.; Perez, V. Low-calorie sweeteners and body weight and composition: A meta-analysis of randomized con-trolled trials and prospective cohort studies. Am. J. Clin. Nutr. 2014, 100, 765–777. [Google Scholar] [CrossRef] [Green Version]
- Schäffler, A.; Schölmerich, J.; Büchler, C. Mechanisms of Disease: Adipocytokines and visceral adipose tissue—emerging role in nonalcoholic fatty liver disease. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 273–280. [Google Scholar] [CrossRef]
- Maersk, M.; Belza, A.; Stødkilde-Jørgensen, H.; Ringgaard, S.; Chabanova, E.; Thomsen, H.; Pedersen, S.B.; Astrup, A.; Richelsen, B. Sucrose-sweetened beverages increase fat storage in the liver, muscle, and visceral fat depot: A 6-mo randomized intervention study. Am. J. Clin. Nutr. 2011, 95, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Antonio-Villa, N.; Bello-Chavolla, O.Y.; Vargas-Vázquez, A.; Mehta, R.; Aguilar-Salinas, C.; Group, M.S.S. The combina-tion of insulin resistance and visceral adipose tissue estimation improves the performance of metabolic syndrome as a predictor of type 2 diabetes. Diabet. Med. 2020, 37, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.-C.; Fujimoto, W.Y.; Hayashi, T.; Kahn, S.E.; Leonetti, D.L.; Boyko, E.J. Increased Visceral Adipose Tissue Is an Independent Predictor for Future Development of Atherogenic Dyslipidemia. J. Clin. Endocrinol. Metab. 2016, 101, 678–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sam, S.; Haffner, S.; Davidson, M.H.; D’Agostino, R.B.; Feinstein, S.; Kondos, G.; Perez, A.; Mazzone, T. Relationship of Abdominal Visceral and Subcutaneous Adipose Tissue With Lipoprotein Particle Number and Size in Type 2 Diabetes. Diabetes 2008, 57, 2022–2027. [Google Scholar] [CrossRef] [Green Version]
- Adeva-Andany, M.M.; Pérez-Felpete, N.; Fernández-Fernández, C.; Donapetry-García, C.; Pazos-García, C. Liver glucose metabolism in humans. Biosci. Rep. 2016, 36, e00416. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [Green Version]
- Abeyrathna, P.; Su, Y. The critical role of Akt in cardiovascular function. Vasc. Pharmacol. 2015, 74, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Crewe, C.; Zhu, Y.; Paschoal, V.A.; Joffin, N.; Ghaben, A.L.; Gordillo, R.; Oh, D.Y.; Liang, G.; Horton, J.D.; Scherer, P.E. SREBP-regulated adipocyte lipogenesis is dependent on substrate availability and redox modulation of mTORC1. JCI Insight 2019, 5, 129397. [Google Scholar] [CrossRef]
- Kimura, N.; Tokunaga, C.; Dalal, S.; Richardson, C.; Yoshino, K.; Hara, K.; Kemp, B.E.; Witters, L.A.; Mimura, O.; Yonezawa, K. A possible linkage between AMP-activated protein kinase (AMPK) and mammalian target of rapamycin (mTOR) signalling pathway. Genes Cells 2003, 8, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Lee, J.; Hong, M.-G.; Lee, B.-H.; Kim, Y.-M.; Chang, P.-S.; Kim, Y.; Yoo, S.-H. Optimization of leucrose produc-tion by dextransucrase from Streptococcus mutans and its application as an adipogenesis regulator. J. Funct. Foods 2017, 39, 238–244. [Google Scholar] [CrossRef]
- Liu, W.; Huang, S.; Li, Y.; Li, Y.; Li, D.; Wu, P.; Wang, Q.; Zheng, X.; Zhang, K. Glycyrrhizic acid from licorice down-regulates inflammatory responses via blocking MAPK and PI3K/Akt-dependent NF-κB signalling pathways in TPA-induced skin inflammation. Med. Chem. Comm. 2018, 9, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, H.-L.; Jung, Y.; Ahn, K.S.; Kwak, H.J.; Um, J.-Y. Bitter Orange (Citrus aurantium Linné) Improves Obesity by Regulating Adipogenesis and Thermogenesis through AMPK Activation. Nutrients 2019, 11, 1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, P. Transcriptional Regulatory Circuits Controlling Brown Fat Development and Activation. Diabetes 2015, 64, 2369–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.; Montague, C.; Prins, J.; Holder, J.C.; Smith, S.A.; Sanders, L.; Digby, J.E.; Sewter, C.P.; Lazar, M.A.; Chatterjee, V.K.; et al. Activators of peroxisome proliferator-activated receptor gamma have depot-specific effects on human preadipocyte differentiation. J. Clin. Investig. 1997, 100, 3149–3153. [Google Scholar] [CrossRef] [Green Version]
- Sears, I.B.; MacGinnitie, M.A.; Kovacs, L.G.; Graves, R.A. Differentiation-dependent expression of the brown adipocyte uncoupling protein gene: Regulation by peroxisome proliferator-activated receptor gamma. Mol. Cell. Biol. 1996, 16, 3410–3419. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Ward, W.F. PGC-1α: A key regulator of energy metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef]
- Puigserver, P.; Wu, Z.; Park, C.W.; Graves, R.; Wright, M.; Spiegelman, B.M. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 1998, 92, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Nagata, N.; Chen, G.; Nagashimada, M.; Zhuge, F.; Ni, Y.; Sakai, Y.; Kaneko, S.; Ota, T. Empagliflozin reverses obesity and insulin resistance through fat browning and alternative macrophage activation in mice fed a high-fat diet. BMJ Open Diabetes Res. Care 2019, 7, e000783. [Google Scholar] [CrossRef]
- Kang, N.H.; Mukherjee, S.; Yun, J.W. Trans-cinnamic acid stimulates white fat browning and activates brown adipocytes. Nutrients 2019, 11, 577. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Horiuchi, Y.; Hironao, K.-y.; Kitakaze, T.; Yamashita, Y.; Ashida, H. Prevention effect of quercetin and its glycosides on obesity and hyperglycemia through activating AMPKα in high-fat diet-fed ICR mice. J. Clin. Biochem. Nutr. 2020, 67, 74–83. [Google Scholar] [CrossRef]
- Viskupičová, J.; Ondrejovič, M.; Šturdík, E. Bioavailability and metabolism of flavonoids. J. Food Nutr. Res. 2008, 47, 151–162. [Google Scholar]
- European Food Safety Authority. Flavouring Group Evaluation 32 (FGE. 32): Flavonoids (Flavanones and dihydrochalcones) from chemical groups 25 and 30. EFSA J. 2010, 8, 1065. [Google Scholar] [CrossRef]
- Kwon, E.-Y.; Lee, J.; Kim, Y.J.; Do, A.; Choi, J.-Y.; Cho, S.-J.; Jung, U.J.; Lee, M.-K.; Park, Y.B.; Choi, M.-S. Seabuckthorn Leaves Extract and Flavonoid Glycosides Extract from Seabuckthorn Leaves Ameliorates Adiposity, Hepatic Steatosis, Insulin Resistance, and Inflammation in Diet-Induced Obesity. Nutrients 2017, 9, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Hassan, Y.I.; Liu, R.; Mats, L.; Yang, C.; Liu, C.; Tsao, R. Molecular Mechanisms Underlying the Absorption of Aglycone and Glycosidic Flavonoids in a Caco-2 BBe1 Cell Model. ACS Omega 2020, 5, 10782–10793. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | AIN 93G |
|---|---|
| gm | |
| Casein, lactic | 200 |
| L-Cystine | 3 |
| Corn Starch | 397 |
| Sucrose | 100 |
| Dextrose | 132 |
| Cellulose | 50 |
| Soybean Oil | 70 |
| t-Butylhydroquinone | 0.014 |
| AIN-93G Mineral Mix | 35 |
| AIN-93 Vitamin Mix | 10 |
| Choline Bitartrate | 2.5 |
| Total | 1000 |
| Name/GeneID | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) | |
|---|---|---|---|
| Acox1 (Acyl-CoA oxidase 1) | 11430 | TTGGAAACCACTGCCACATA | AGGCATGTAACCCGTAGCAC |
| Acsl1 (Acyl-CoA synthetase long-chain family member 1) | 14081 | TGCCAGAGCTGATTGACATTC | GGCATACCAGAAGGTGGTGAG |
| Cd36 (Cluster of differentiation 36) | 12491 | GTGCTCTCCCTTGATTCTGC | TGAGAATGCCTCCAAACACA |
| C/ebpα (CCAAT/enhancer binding protein alpha) | 12606 | CCAAGAAGTCGGTGGACAAGA | CGGTCATTGTCACTGGTCAACT |
| Cpt1α (Carnitine palmitoyltransferase 1 alpha) | 12894 | AACCCAGTGCCTTAACGATG | GAACTGGTGGCCAATGAGAT |
| Fas (Fatty acid synthase) | 14104 | TGTGAGTGGTTCAGAGGCAT | TTCTGTAGTGCCAGCAAGCT |
| Il-1β (Interleukin 1 beta) | 16176 | ATGGCAACTGTTCCTGAACTCAACT | CAGGACAGGTATAGATTCTTTCCTTT |
| Lpl (Lipoprotein lipase) | 16956 | GAGTTTGACCGCCTTCCG | TCCCGTTACCGTCCATCC |
| Mcp1 (Monocyte chemoattractant protein 1) | 17224 | CTTCTGGGCCTGCTGTTCA | CCAGCCTACTCATTGGGATCA |
| Pgc1α (Peroxisome proliferator-activated receptor gamma coactivator 1 alpha) | 19017 | TCGAGCTGTACTTTTGTGGA | TCATACTTGCTCTTGGTGGA |
| Pparγ (Peroxisome proliferator-activated receptor gamma) | 19016 | GAGCACTTCACAAGAAATTACC | GAACTCCATAGTGGAAGCCT |
| Prdm16 (Positive regulatory domain 16) | 70673 | AGATGAACCAGGCATCCACT | TCTACGTCCTCTGGCTTTGC |
| Srebp1c (Sterol regulatory element-binding protein 1) | 20787 | TAGAGCATATCCCCCAGGTG | GGTACGGGCCACAAGAAGTA |
| Tnfα (Tumor necrosis factor alpha) | 21926 | ATGAGAAGTTCCCAAATGGC | CTCCACTTGGTGGTTTGCTA |
| Ucp1 (Uncoupling protein 1) | 22227 | CCAAGCCAGGATGGTGAAC | CCAGCGGGAAGGTGATGATA |
| Glut4 (Glucose transporter type 4) | 20528 | TGTTCAATCACCTGGTTGCG | CTTGGCTCCCTTCAGTTTGG |
| Gapdh (Glyceraldehyde 3-phosphate dehydrogenase) | 14433 | AACTTTGGCATTGTGGAAGG | TGTGAGGGAGATGCTCAGTG |
| Ctrl | NHDC | GNHDC | p-Value | |
|---|---|---|---|---|
| Final body weight (g) | 37.75 ± 1.02 a | 35.23 ± 0.64 a | 35.32 ± 1.26 a | 0.1551 |
| Body weight gain (g) | 9.80 ± 0.67 a | 8.01 ± 0.69 a | 6.95 ± 0.79 b | 0.0314 |
| Food intake (g/day) | 6.25 ± 0.20 a | 5.90 ± 0.20 a | 6.00 ± 0.27 a | 0.5340 |
| Water intake (mL/day) | 18.31 ± 1.47 a | 14.68 ± 1.65 a | 14.92 ± 1.68 a | 0.2216 |
| Liver (g) | 2.31 ± 0.05 a | 2.13 ± 0.09 a | 2.08 ± 0.12 a | 0.1874 |
| Subcutaneous adipose tissues (g) | 2.46 ± 0.47 a | 1.27 ± 0.16 b | 1.49 ± 0.19 b | 0.0247 |
| Visceral adipose tissues (g) | 1.22 ± 0.07 a | 1.22 ± 0.07 a | 1.12 ± 0.04 a | 0.4228 |
| Total adipose tissues (g) | 5.66 ± 0.49 a | 4.42 ± 0.20 b | 4.57 ± 0.24 b | 0.0299 |
| Total cholesterol (mg/dL) | 159.39 ± 3.60 a | 158.81 ± 2.57 a | 146.17 ± 4.03 b | 0.0193 |
| LDL-cholesterol (mg/dL) | 76.50 ± 4.80 a | 73.47 ± 2.06 a | 61.29 ± 6.37 a | 0.0766 |
| HDL-cholesterol (mg/dL) | 53.54 ± 3.02 a | 53.82 ± 2.10 a | 54.77 ± 5.21 a | 0.9205 |
| Triacylglycerols (mg/dL) | 146.78 ± 3.77 a | 157.61 ± 7.09 a | 146.91 ± 11.81 a | 0.5736 |
| NEFA (mEq/L) | 0.73 ± 0.04 a | 0.63 ± 0.02 a | 0.61 ± 0.04 b | 0.0444 |
| Leptin (ng/dL) | 30.05 ± 1.24 a | 31.27 ± 1.74 a | 25.67 ± 3.37 a | 0.2161 |
| Adiponectin (ng/dL) | 0.54 ± 0.02 ab | 0.50 ± 0.02 a | 0.62 ± 0.04 b | 0.0377 |
| Insulin (ng/dL) | 1.77 ± 0.28 a | 2.95 ± 0.36 b | 1.42 ± 0.24 a | 0.0035 |
| HOMA-IR | 41.28 ± 21.05 ab | 58.86 ± 20.98 a | 29.83 ± 19.39 b | 0.0206 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, M.; Kim, Y.; Lee, J.; Manthey, J.A.; Kim, Y.; Kim, Y. Neohesperidin Dihydrochalcone and Neohesperidin Dihydrochalcone-O-Glycoside Attenuate Subcutaneous Fat and Lipid Accumulation by Regulating PI3K/AKT/mTOR Pathway In Vivo and In Vitro. Nutrients 2022, 14, 1087. https://doi.org/10.3390/nu14051087
Kwon M, Kim Y, Lee J, Manthey JA, Kim Y, Kim Y. Neohesperidin Dihydrochalcone and Neohesperidin Dihydrochalcone-O-Glycoside Attenuate Subcutaneous Fat and Lipid Accumulation by Regulating PI3K/AKT/mTOR Pathway In Vivo and In Vitro. Nutrients. 2022; 14(5):1087. https://doi.org/10.3390/nu14051087
Chicago/Turabian StyleKwon, Minseo, Yerin Kim, Jihye Lee, John A. Manthey, Yang Kim, and Yuri Kim. 2022. "Neohesperidin Dihydrochalcone and Neohesperidin Dihydrochalcone-O-Glycoside Attenuate Subcutaneous Fat and Lipid Accumulation by Regulating PI3K/AKT/mTOR Pathway In Vivo and In Vitro" Nutrients 14, no. 5: 1087. https://doi.org/10.3390/nu14051087