
Is Bitterness Only a Taste? The Expanding Area of Health Benefits of Brassica Vegetables and Potential for Bitter Taste Receptors to Support Health Benefits



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
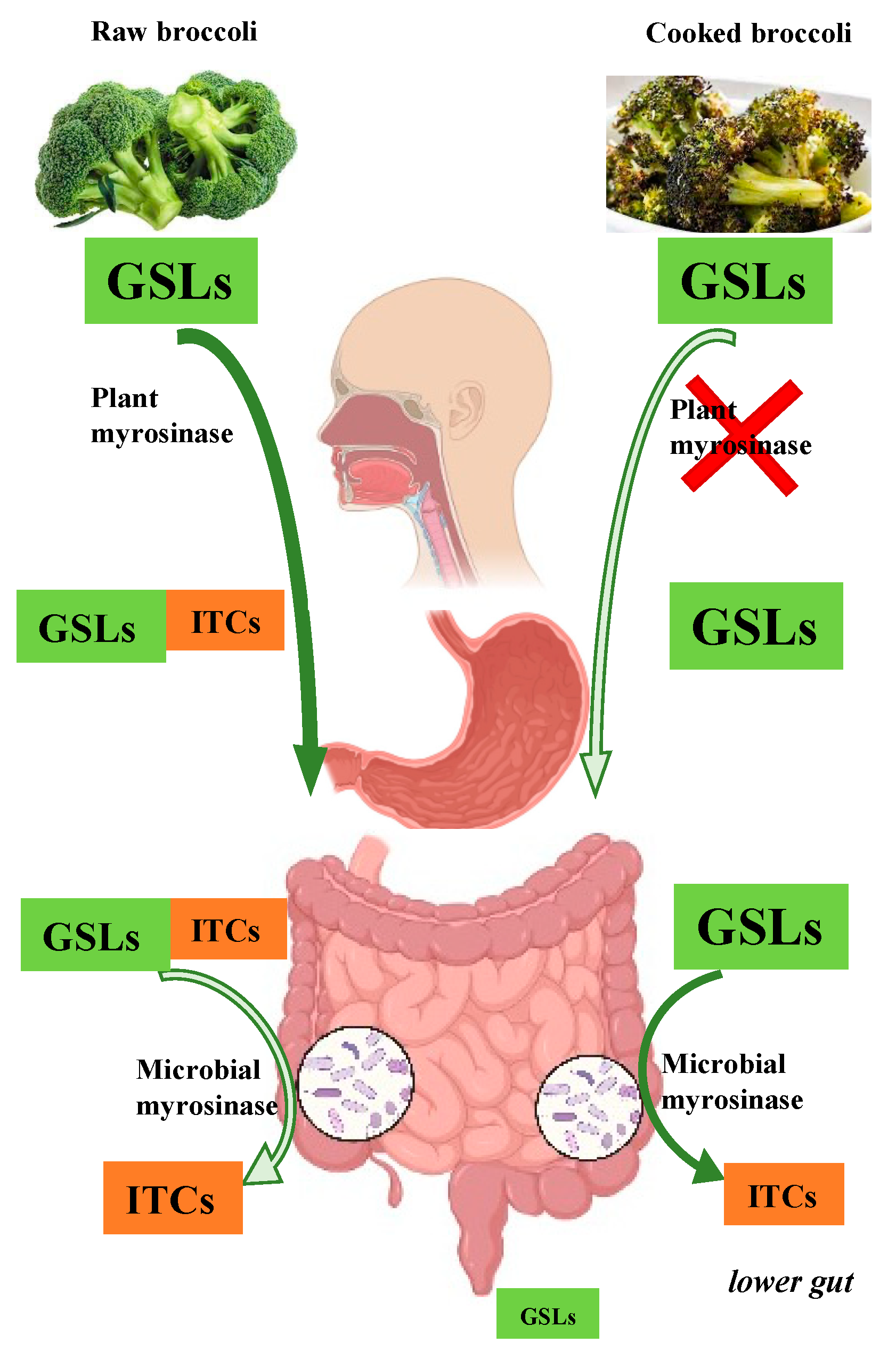
2. Metabolism of Glucosinolates and Resulting ITC Bioavailability: A Key Source of Variability in Health Benefits
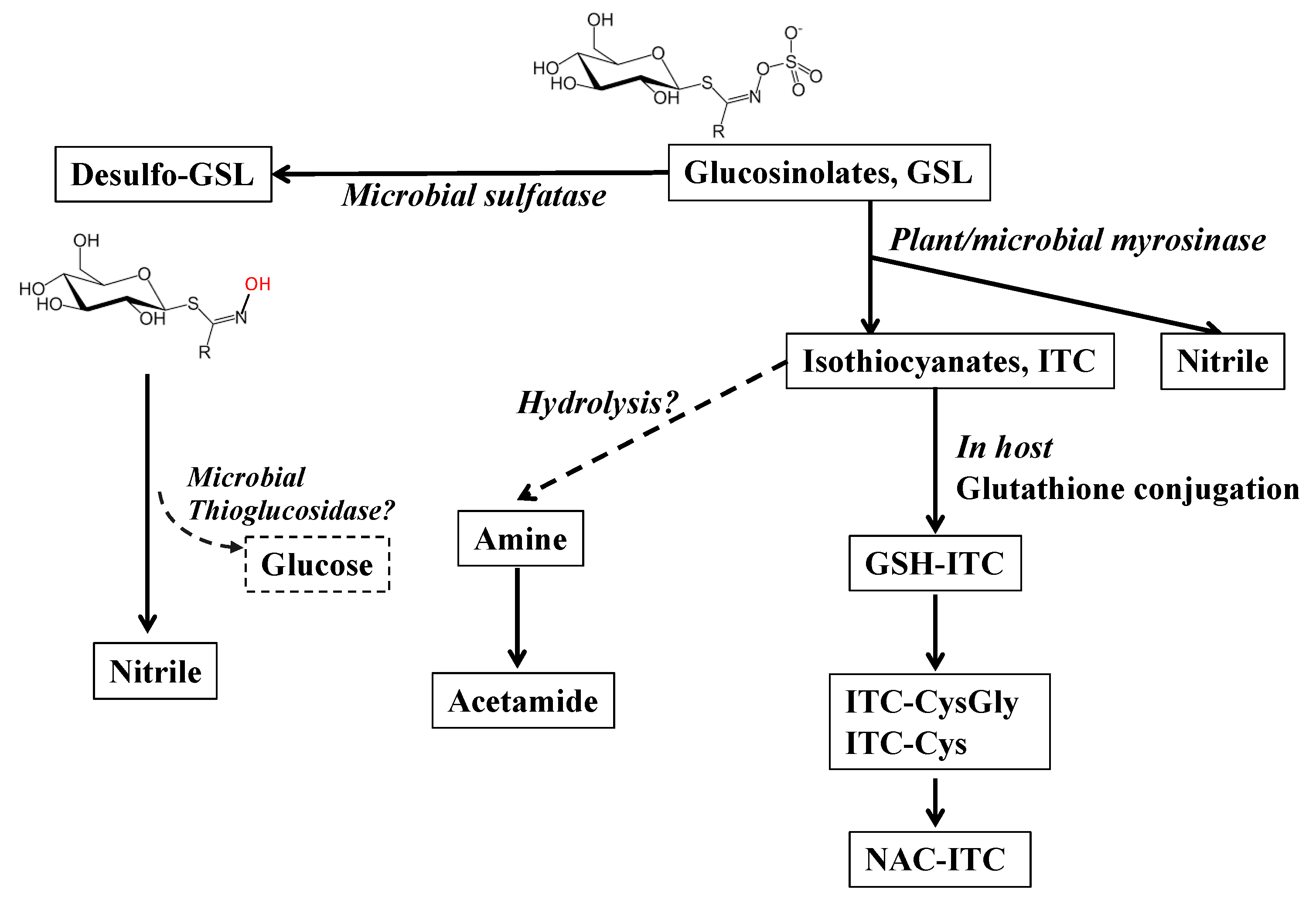
2.1. Metabolic Fate of Glucosinolates
2.2. Microbial Metabolism of Glucosinolates
2.3. Alternative Pathways of Microbial Metabolism of Glucosinolates
3. Health Benefits of Brassica Vegetables and Associated Mechanisms
3.1. Health Benefits of ITCs Working via Nrf2 Pathways
3.1.1. Prevention of Oxidative Stress and Inflammation
3.1.2. Chemopreventive Effects
3.1.3. Metabolic Syndrome
3.2. Health Benefits of ITCs through Mechanisms Other Than Nrf2
4. A Novel Interaction: The Bitter Brassica Vegetables and Intestinal Bitter Taste Receptors
4.1. Glucose Metabolism and Obesity
4.2. Innate Immunity
4.3. Neuroprotection
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sikorska-Zimny, K.; Beneduce, L. The Glucosinolates and Their Bioactive Derivatives in Brassica: A Review on Classification, Biosynthesis and Content in Plant Tissues, Fate during and after Processing, Effect on the Human Organism and Interaction with the Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2020, 61, 2544–2571. [Google Scholar] [CrossRef] [PubMed]
- Favela-González, K.M.; Hernández-Almanza, A.Y.; de la Fuente-Salcido, N.M. The Value of Bioactive Compounds of Cruciferous Vegetables (Brassica) as Antimicrobials and Antioxidants: A Review. J. Food Biochem. 2020, 44, e13414. [Google Scholar] [CrossRef] [PubMed]
- Maina, S.; Misinzo, G.; Bakari, G.; Kim, H.-Y. Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review. Molecules 2020, 25, 3682. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.C.; Frandsen, H.B.; Jensen, S.K.; Kristensen, N.B.; Sørensen, S.; Sørensen, H. Bioavailability and in Vivo Metabolism of Intact Glucosinolates. J. Funct. Foods 2016, 24, 450–460. [Google Scholar] [CrossRef]
- Abdull Razis, A.F.; Bagatta, M.; de Nicola, G.R.; Iori, R.; Ioannides, C. Intact Glucosinolates Modulate Hepatic Cytochrome P450 and Phase II Conjugation Activities and May Contribute Directly to the Chemopreventive Activity of Cruciferous Vegetables. Toxicology 2010, 277, 74–85. [Google Scholar] [CrossRef]
- Angelino, D.; Jeffery, E. Glucosinolate Hydrolysis and Bioavailability of Resulting Isothiocyanates: Focus on Glucoraphanin. J. Funct. Foods 2014, 7, 67–76. [Google Scholar] [CrossRef]
- Gu, H.F.; Mao, X.Y.; Du, M. Metabolism, Absorption, and Anti-Cancer Effects of Sulforaphane: An Update. Crit. Rev. Food Sci. Nutr. 2021, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mi, E.N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Swiergiel, A.H.; Wacław, P. Health Benefits of Plant-Derived Sulfur Compounds, Glucosinolates, and Organosulfur Compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef]
- Sikorska-Zimny, K.; Beneduce, L. The Metabolism of Glucosinolates by Gut Microbiota. Nutrients 2021, 13, 2750. [Google Scholar] [CrossRef]
- Narbad, A.; Rossiter, J.T. Gut Glucosinolate Metabolism and Isothiocyanate Production. Mol. Nutr. Food Res. 2018, 62, 1700991. [Google Scholar] [CrossRef]
- Bouranis, J.A.; Beaver, L.M.; Ho, E. Metabolic Fate of Dietary Glucosinolates and Their Metabolites: A Role for the Microbiome. Front. Nutr. 2021, 8, 748433. [Google Scholar] [CrossRef] [PubMed]
- Angelino, D.; Jeffery, E.H. Editorial: Glucosinolate Metabolites: Bioavailability, Bioactivity and Clinical Variability. Front. Nutr. 2021, 8, 823203. [Google Scholar] [CrossRef] [PubMed]
- Sønderby, I.E.; Geu-Flores, F.; Halkier, B.A. Biosynthesis of Glucosinolates—Gene Discovery and Beyond. Trends Plant Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; Cartea, M.E.; Moreno, D.A.; Tortosa, M.; Francisco, M. Processing and Cooking Effects on Glucosinolates and Their Derivatives. Glucosinolates Prop. Recov. Appl. 2020, 181–212. [Google Scholar] [CrossRef]
- Martínez, S.; Armesto, J.; Gómez-Limia, L.; Carballo, J. Impact of Processing and Storage on the Nutritional and Sensory Properties and Bioactive Components of Brassica spp. A Review. Food Chem. 2020, 313, 126065. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica Foods: Bioavailability in Food and Significance for Human Health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Barba, F.J.; Nikmaram, N.; Roohinejad, S.; Khelfa, A.; Zhu, Z.; Koubaa, M. Bioavailability of Glucosinolates and Their Breakdown Products: Impact of Processing. Front. Nutr. 2016, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Kuchernig, J.C.; Burow, M.; Wittstock, U. Evolution of Specifier Proteins in Glucosinolate-Containing Plants. BMC Evol. Biol. 2012, 12, 127. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Pang, X.; Yang, T. Microwave Cooking Increases Sulforaphane Level in Broccoli. Food Sci. Nutr. 2020, 8, 2052–2058. [Google Scholar] [CrossRef] [Green Version]
- Matusheski, N.V.; Juvik, J.A.; Jeffery, E.H. Heating Decreases Epithiospecifier Protein Activity and Increases Sulforaphane Formation in Broccoli. Phytochemistry 2004, 65, 1273–1281. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Hoeflinger, J.L.; Neme, B.P.; Jeffery, E.H.; Miller, M.J. Dietary Broccoli Alters Rat Cecal Microbiota to Improve Glucoraphanin Hydrolysis to Bioactive Isothiocyanates. Nutrients 2017, 9, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okunade, O.; Niranjan, K.; Ghawi, S.K.; Kuhnle, G.; Methven, L. Supplementation of the Diet by Exogenous Myrosinase via Mustard Seeds to Increase the Bioavailability of Sulforaphane in Healthy Human Subjects after the Consumption of Cooked Broccoli. Mol. Nutr. Food Res. 2018, 62, 1700980. [Google Scholar] [CrossRef] [PubMed]
- Bheemreddy, R.M.; Jeffery, E.H. The Metabolic Fate of Purified Glucoraphanin in F344 Rats. J. Agric. Food Chem. 2007, 55, 2861–2866. [Google Scholar] [CrossRef]
- Egner, P.A.; Chen, J.G.; Wang, J.B.; Wu, Y.; Sun, Y.; Lu, J.H.; Zhu, J.; Zhang, Y.H.; Chen, Y.S.; Friesen, M.D.; et al. Bioavailability of Sulforaphane from Two Broccoli Sprout Beverages: Results of a Short-Term, Cross-over Clinical Trial in Qidong, China. Cancer Prev. Res. 2011, 4, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Fahey, J.W.; Holtzclaw, W.D.; Wehage, S.L.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Sulforaphane Bioavailability from Glucoraphanin-Rich Broccoli: Control by Active Endogenous Myrosinase. PLoS ONE 2015, 10, e0140963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petri, N.; Tannergren, C.; Holst, B.; Mellon, F.A.; Bao, Y.; Plumb, G.W.; Bacon, J.; O’Leary, K.A.; Kroon, P.A.; Knutson, L.; et al. Absorption/metabolism of sulforaphane and quercetin, and regulation of phase ii enzymes, in human jejunum in vivo. Drug Metab. Dispos. 2003, 31, 805–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, J.M.; Hait, W.N. Pharmacology of Drugs That Alter Multidrug Resistance in Cancer. Pharmacol. Rev. 1990, 42, 155–199. [Google Scholar]
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Human Metabolism and Excretion of Cancer Chemoprotective Glucosinolates and Isothiocyanates of Cruciferous Vegetables. Cancer Epidemiol. Prev. Biomark. 1998, 7, 1091–1100. [Google Scholar]
- Rouzaud, G.; Young, S.A.; Duncan, A.J. Hydrolysis of Glucosinolates to Isothiocyanates after Ingestion of Raw or Microwaved Cabbage by Human Volunteers. Cancer Epidemiol. Prev. Biomark. 2004, 13, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jeffery, E.H.; Miller, M.J.; Wallig, M.A.; Wu, Y. Lightly Cooked Broccoli Is as Effective as Raw Broccoli in Mitigating Dextran Sulfate Sodium-Induced Colitis in Mice. Nutrients 2018, 10, 748. [Google Scholar] [CrossRef] [Green Version]
- Albaser, A.; Kazana, E.; Bennett, M.H.; Cebeci, F.; Luang-In, V.; Spanu, P.D.; Rossiter, J.T. Discovery of a Bacterial Glycoside Hydrolase Family 3 (GH3) β-Glucosidase with Myrosinase Activity from a Citrobacter Strain Isolated from Soil. J. Agric. Food Chem. 2016, 64, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- Liou, C.S.; Sirk, S.J.; Diaz, C.A.C.; Klein, A.P.; Fischer, C.R.; Higginbottom, S.K.; Erez, A.; Donia, M.S.; Sonnenburg, J.L.; Sattely, E.S. A Metabolic Pathway for Activation of Dietary Glucosinolates by a Human Gut Symbiont. Cell 2020, 180, 717–728.e19. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Hullar, M.A.J.; Schwarz, Y.; Lampe, J.W. Human Gut Bacterial Communities Are Altered by Addition of Cruciferous Vegetables to a Controlled Fruit- and Vegetable-Free Diet. J. Nutr. 2009, 139, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, J.L.; Liu, X.; Charron, C.S.; Novotny, J.A.; Jeffery, E.H.; Seifried, H.E.; Ross, S.A.; Miller, M.J.; Swanson, K.S.; Holscher, H.D. Broccoli Consumption Affects the Human Gastrointestinal Microbiota. J. Nutr. Biochem. 2019, 63, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zandani, G.; Kaftori-Sandler, N.; Sela, N.; Nyska, A.; Madar, Z. Dietary Broccoli Improves Markers Associated with Glucose and Lipid Metabolism through Modulation of Gut Microbiota in Mice. Nutrition 2021, 90, 111240. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Wehage, S.L.; Holtzclaw, W.D.; Kensler, T.W.; Egner, P.A.; Shapiro, T.A.; Talalay, P. Protection of Humans by Plant Glucosinolates: Efficiency of Conversion of Glucosinolates to Isothiocyanates by the Gastrointestinal Microflora. Physiol. Behav. 2012, 176, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Hullar, M.A.; Beresford, S.A.; Lampe, J.W. Variation of Glucoraphanin Metabolism in Vivo and Ex Vivo by Human Gut Bacteria. Br. J. Nutr. 2011, 106, 408–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beran, F.; Sporer, T.; Paetz, C.; Ahn, S.J.; Betzin, F.; Kunert, G.; Shekhov, A.; Vassão, D.G.; Bartram, S.; Lorenz, S.; et al. One Pathway Is Not Enough: The Cabbage Stem Flea Beetle Psylliodes Chrysocephala Uses Multiple Strategies to Overcome the Glucosinolate-Myrosinase Defense in Its Host Plants. Front. Plant Sci. 2018, 871, 1754. [Google Scholar] [CrossRef]
- Shukla, S.P.; Beran, F. Gut Microbiota Degrades Toxic Isothiocyanates in a Flea Beetle Pest. Mol. Ecol. 2020, 29, 4692–4705. [Google Scholar] [CrossRef]
- Welte, C.U.; de Graaf, R.M.; van den Bosch, T.J.M.; Op den Camp, H.J.M.; van Dam, N.M.; Jetten, M.S.M. Plasmids from the Gut Microbiome of Cabbage Root Fly Larvae Encode SaxA That Catalyses the Conversion of the Plant Toxin 2-Phenylethyl Isothiocyanate. Environ. Microbiol. 2016, 18, 1379–1390. [Google Scholar] [CrossRef]
- Welte, C.U.; Rosengarten, J.F.; de Graaf, R.M.; Jetten, M.S.M. SaxA-Mediated Isothiocyanate Metabolism in Phytopathogenic Pectobacteria. Appl. Environ. Microbiol. 2016, 82, 2372–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.J.; Betzin, F.; Gikonyo, M.W.; Yang, Z.L.; Köllner, T.G.; Beran, F. Identification and Evolution of Glucosinolate Sulfatases in a Specialist Flea Beetle. Sci. Rep. 2019, 9, 15725. [Google Scholar] [CrossRef] [PubMed]
- Falk, K.L.; Gershenzon, J. The Desert Locust, Schistocerca Gregaria, Detoxifies the Glucosinolates of Schouwia Purpurea by Desulfation. J. Chem. Ecol. 2007, 33, 1542–1555. [Google Scholar] [CrossRef] [PubMed]
- Luang-In, V.; Albaser, A.A.; Nueno-Palop, C.; Bennett, M.H.; Narbad, A.; Rossiter, J.T. Glucosinolate and Desulfo-Glucosinolate Metabolism by a Selection of Human Gut Bacteria. Curr. Microbiol. 2016, 73, 442–451. [Google Scholar] [CrossRef]
- Bouranis, J.A.; Beaver, L.M.; Choi, J.; Wong, C.P.; Jiang, D.; Sharpton, T.J.; Stevens, J.F.; Ho, E. Composition of the Gut Microbiome Influences Production of Sulforaphane-Nitrile and Iberin-Nitrile from Glucosinolates in Broccoli Sprouts. Nutrients 2021, 13, 3013. [Google Scholar] [CrossRef]
- Sturm, C.; Wagner, A.E. Brassica-Derived Plant Bioactives as Modulators of Chemopreventive and Inflammatory Signaling Pathways. Int. J. Mol. Sci. 2017, 18, 1890. [Google Scholar] [CrossRef] [Green Version]
- Fofaria, N.M.; Ranjan, A.; Kim, S.H.; Srivastava, S.K. Mechanisms of the Anticancer Effects of Isothiocyanates. Enzymes 2015, 37, 111–137. [Google Scholar] [CrossRef]
- Esteve, M. Mechanisms Underlying Biological Effects of Cruciferous Glucosinolate-Derived Isothiocyanates/Indoles: A Focus on Metabolic Syndrome. Front. Nutr. 2020, 7, 111. [Google Scholar] [CrossRef]
- Dufour, V.; Stahl, M.; Baysse, C. The Antibacterial Properties of Isothiocyanates. Microbiology 2015, 161, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 Defines a Physiologically Important Stress Response Mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 Regulatory Network Provides an Interface between Redox and Intermediary Metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Yamamoto, M. The KEAP1NRF2 System in Cancer. Front. Oncol. 2017, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 Signaling Pathway: Pivotal Roles in Inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting Molecular Cross-Talk between Nrf2 and NF-ΚB Response Pathways. Biochem. Soc. Trans. 2015, 43, 621. [Google Scholar] [CrossRef] [Green Version]
- Nagata, N.; Xu, L.; Kohno, S.; Ushida, Y.; Aoki, Y.; Umeda, R.; Fuke, N.; Zhuge, F.; Ni, Y.; Nagashimada, M.; et al. Glucoraphanin Ameliorates Obesity and Insulin Resistance through Adipose Tissue Browning and Reduction of Metabolic Endotoxemia in Mice. Diabetes 2017, 66, 1222–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleszczyński, K.; Ernst, I.M.A.; Wagner, A.E.; Kruse, N.; Zillikens, D.; Rimbach, G.; Fischer, T.W. Sulforaphane and Phenylethyl Isothiocyanate Protect Human Skin against UVR-Induced Oxidative Stress and Apoptosis: Role of Nrf2-Dependent Gene Expression and Antioxidant Enzymes. Pharmacol. Res. 2013, 78, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Cykowiak, M.; Krajka-Kuźniak, V.; Kleszcz, R.; Kucińska, M.; Szaefer, H.; Piotrowska-Kempisty, H.; Plewiński, A.; Murias, M.; Baer-Dubowska, W. Comparison of the Impact of Xanthohumol and Phenethyl Isothiocyanate and Their Combination on Nrf2 and NF-ΚB Pathways in HepG2 Cells In Vitro and Tumor Burden In Vivo. Nutrients 2021, 13, 3000. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.; Blum, N.M.; Mueller, A.S. Examination of the Anti-Inflammatory, Antioxidant, and Xenobiotic-Inducing Potential of Broccoli Extract and Various Essential Oils during a Mild DSS-Induced Colitis in Rats. Corp. ISRN Gastroenterol. 2013, 2013, 710856. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Wright, S.E.; Kim, S.H.; Srivastava, S.K. Phenethyl Isothiocyanate: A Comprehensive Review of Anti-Cancer Mechanisms. Biochim. Biophys. Acta 2014, 1846, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.K.; Xiao, D.; Lew, K.L.; Hershberger, P.; Kokkinakis, D.M.; Johnson, C.S.; Trump, D.L.; Singh, S.V. Allyl Isothiocyanate, a Constituent of Cruciferous Vegetables, Inhibits Growth of PC-3 Human Prostate Cancer Xenografts in Vivo. Carcinogenesis 2003, 24, 1665–1670. [Google Scholar] [CrossRef]
- Rose, P.; Faulkner, K.; Williamson, G.; Mithen, R. 7-Methylsulfinylheptyl and 8-Methylsulfinyloctyl Isothiocyanates from Watercress Are Potent Inducers of Phase II Enzymes. Carcinogenesis 2000, 21, 1983–1988. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Nagata, N.; Ota, T. Glucoraphanin: A Broccoli Sprout Extract That Ameliorates Obesity-Induced Inflammation and Insulin Resistance. Adipocyte 2018, 7, 218–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelsson, A.S.; Tubbs, E.; Mecham, B.; Chacko, S.; Nenonen, H.A.; Tang, Y.; Fahey, J.W.; Derry, J.M.J.; Wollheim, C.B.; Wierup, N.; et al. Sulforaphane Reduces Hepatic Glucose Production and Improves Glucose Control in Patients with Type 2 Diabetes. Sci. Transl. Med. 2017, 9, eaah4477. [Google Scholar] [CrossRef] [Green Version]
- Abukhabta, S.; Khalil Ghawi, S.; Karatzas, K.A.; Charalampopoulos, D.; McDougall, G.; Allwood, J.W.; Verrall, S.; Lavery, S.; Latimer, C.; Pourshahidi, L.K.; et al. Sulforaphane-Enriched Extracts from Glucoraphanin-Rich Broccoli Exert Antimicrobial Activity against Gut Pathogens in Vitro and Innovative Cooking Methods Increase in Vivo Intestinal Delivery of Sulforaphane. Eur. J. Nutr. 2021, 60, 1263–1276. [Google Scholar] [CrossRef]
- Shahinozzaman, M.; Raychaudhuri, S.; Fan, S.; Obanda, D.N. Kale Attenuates Inflammation and Modulates Gut Microbial Composition and Function in C57BL/6J Mice with Diet-Induced Obesity. Microorganisms 2021, 9, 238. [Google Scholar] [CrossRef]
- Li, X.; Tian, S.; Wang, Y.; Liu, J.; Wang, J.; Lu, Y. Broccoli Microgreens Juice Reduces Body Weight by Enhancing Insulin Sensitivity and Modulating Gut Microbiota in High-Fat Diet-Induced C57BL/6J Obese Mice. Eur. J. Nutr. 2021, 60, 3829–3839. [Google Scholar] [CrossRef]
- Kellingray, L.; Tapp, H.S.; Saha, S.; Doleman, J.F.; Narbad, A.; Mithen, R.F. Consumption of a Diet Rich in Brassica Vegetables Is Associated with a Reduced Abundance of Sulphate-reducing Bacteria: A Randomised Crossover Study. Mol. Nutr. Food Res. 2017, 61, 1600992. [Google Scholar] [CrossRef] [Green Version]
- Hoon, M.A.; Adler, E.; Lindemeier, J.; Battey, J.F.; Ryba, N.J.P.; Zuker, C.S. Putative Mammalian Taste Receptors: A Class of Taste-Specific GPCRs with Distinct Topographic Selectivity. Cell 1999, 96, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Lossow, K.; Hübner, S.; Roudnitzky, N.; Slack, J.P.; Pollastro, F.; Behrens, M.; Meyerhof, W. Comprehensive Analysis of Mouse Bitter Taste Receptors Reveals Different Molecular Receptive Ranges for Orthologous Receptors in Mice and Humans. J. Biol. Chem. 2016, 291, 15358–15377. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Zhang, C.H.; Lifshitz, L.M.; ZhuGe, R. Extraoral Bitter Taste Receptors in Health and Disease. J. Gen. Physiol. 2017, 149, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Tuzim, K.; Korolczuk, A. An Update on Extra-Oral Bitter Taste Receptors. J. Trans. Med. 2021, 19, 440. [Google Scholar] [CrossRef]
- Avau, B.; Depoortere, I. The Bitter Truth about Bitter Taste Receptors: Beyond Sensing Bitter in the Oral Cavity. Acta Physiol. 2016, 216, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Avau, B.; Rotondo, A.; Thijs, T.; Andrews, C.N.; Janssen, P.; Tack, J.; Depoortere, I. Targeting Extra-Oral Bitter Taste Receptors Modulates Gastrointestinal Motility with Effects on Satiation. Sci. Rep. 2015, 5, 15985. [Google Scholar] [CrossRef]
- Yue, X.; Liang, J.; Gu, F.; Du, D.; Chen, F. Berberine Activates Bitter Taste Responses of Enteroendocrine STC-1 Cells. Mol. Cell. Biochem. 2018, 447, 21–32. [Google Scholar] [CrossRef]
- Chen, M.C.; Wu, S.V.; Reeve, J.R.; Rozengurt, E. Bitter Stimuli Induce Ca2+ Signaling and CCK Release in Enteroendocrine STC-1 Cells: Role of L-Type Voltage-Sensitive Ca2+ Channels. Am. J. Physiol. Cell Physiol. 2006, 291, 726–739. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Wang, X.; Young, R.L.; Horowitz, M.; Rayner, C.K.; Wu, T. Role of Intestinal Bitter Sensing in Enteroendocrine Hormone Secretion and Metabolic Control. Front. Endocrinol. 2018, 9, 576. [Google Scholar] [CrossRef]
- Deshpande, D.A.; Wang, W.C.H.; Mcllmoyle, E.L.; Robinett, K.S.; Schillinger, R.M.; An, S.S.; Sham, J.S.K.; Liggett, S.B. Bitter Taste Receptors on Airway Smooth Muscle Bronchodilate by a Localized Calcium Flux and Reverse Obstruction. Nat. Med. 2010, 16, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.C.; Rosado, T.; Costa, A.R.; Santos, J.; Gallardo, E.; Quintela, T.; Ishikawa, H.; Schwerk, C.; Schroten, H.; Gonçalves, I.; et al. The Bitter Taste Receptor TAS2R14 Regulates Resveratrol Transport across the Human Blood-Cerebrospinal Fluid Barrier. Biochem. Pharmacol. 2020, 177, 113953. [Google Scholar] [CrossRef]
- Kok, B.P.; Galmozzi, A.; Littlejohn, N.K.; Albert, V.; Godio, C.; Kim, W.; Kim, S.M.; Bland, J.S.; Grayson, N.; Fang, M.; et al. Intestinal Bitter Taste Receptor Activation Alters Hormone Secretion and Imparts Metabolic Benefits. Mol. Metab. 2018, 16, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H. Variation in the TAS2R38 Bitterness Receptor Gene Was Associated with Food Consumption and Obesity Risk in Koreans. Nutrients 2019, 11, 1973. [Google Scholar] [CrossRef] [Green Version]
- Dotson, C.D.; Zhang, L.; Xu, H.; Shin, Y.K.; Vigues, S.; Ott, S.H.; Elson, A.E.T.; Choi, H.J.; Shaw, H.; Egan, J.M.; et al. Bitter Taste Receptors Influence Glucose Homeostasis. PLoS ONE 2008, 3, e3974. [Google Scholar] [CrossRef] [Green Version]
- Caremoli, F.; Huynh, J.; Lagishetty, V.; Jacobs, J.; Braun, J.; Sternini, C. Abstract-Bitter Taste Receptors, T2R138 and T2R16, Are Induced in the Large Intestine of Male and Female Mice on a High Fat Diet in a Microbiota-Dependent Manner. Gastroenterology 2017, 152, S156. [Google Scholar] [CrossRef]
- Caremoli, F.; Huynh, J.; de Giorgio, R.; Sternini, C. Fecal Transplantation from Obese, High-Fat Fed Mice Affects Fat and Lean Body Mass and Gut Bitter Taste Receptor Subtypes, T2R138 and T2R116, Expression. Gastroenterology 2018, 154, S568. [Google Scholar] [CrossRef]
- Bufe, B.; Breslin, P.A.S.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.K.; Drayna, D.; Meyerhof, W. The Molecular Basis of Individual Differences in Phenylthiocarbamide and Propylthiouracil Bitterness Perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.T.T.; Stetter, R.; Herz, C.; Spöttel, J.; Krell, M.; Hanschen, F.S.; Schreiner, M.; Rohn, S.; Behrens, M.; Lamy, E. Allyl Isothiocyanate: A TAS2R38 Receptor-Dependent Immune Modulator at the Interface Between Personalized Medicine and Nutrition. Front. Immunol. 2021, 12, 1255. [Google Scholar] [CrossRef] [PubMed]
- Baik, H.Y.; Juvik, J.A.; Jeffery, E.H.; Wallig, M.A.; Kushad, M.; Klein, B.P. Relating Glucosinolate Content and Flavor of Broccoli Cultivars. J. Food Sci. 2003, 68, 1043–1050. [Google Scholar] [CrossRef]
- Tran, H.T.T.; Herz, C.; Ruf, P.; Stetter, R.; Lamy, E. Human T2R38 Bitter Taste Receptor Expression in Resting and Activated Lymphocytes. Front. Immunol. 2018, 9, 2949. [Google Scholar] [CrossRef]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The Molecular Receptive Ranges of Human TAS2R Bitter Taste Receptors. Chem. Senses 2009, 35, 157–170. [Google Scholar] [CrossRef]
- Hao, S.; Sternini, C.; Raybould, H.E. Role of CCK1 and Y2 Receptors in Activation of Hindbrain Neurons Induced by Intragastric Administration of Bitter Taste Receptor Ligands. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R33–R38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, S.; Laermans, J.; Verhulst, P.J.; Thijs, T.; Tack, J.; Depoortere, I. Bitter Taste Receptors and α-Gustducin Regulate the Secretion of Ghrelin with Functional Effects on Food Intake and Gastric Emptying. Proc. Natl. Acad. Sci. USA 2011, 108, 2094–2099. [Google Scholar] [CrossRef] [Green Version]
- Thorup, A.C.; Kristensen, H.L.; Kidmose, U.; Lambert, M.N.T.; Christensen, L.P.; Fretté, X.; Clausen, M.R.; Hansen, S.M.; Jeppesen, P.B. Strong and Bitter Vegetables from Traditional Cultivars and Cropping Methods Improve the Health Status of Type 2 Diabetics: A Randomized Control Trial. Nutrients 2021, 13, 1831. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.; Yehuda, B.S.; Moninger, T.O.; Kline, J.N.; Welsh, M.J. Motile Cilia of Human Airway Epithelia Are Chemosensory. Science 2009, 325, 1131. [Google Scholar] [CrossRef] [Green Version]
- Adappa, N.D.; Zhang, Z.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Lysenko, A.; Reed, D.R.; Scott, T.; Zhao, N.W.; Owens, D.; et al. The Bitter Taste Receptor T2R38 Is an Independent Risk Factor for Chronic Rhinosinusitis Requiring Sinus Surgery. Int. Forum Allergy Rhinol. 2014, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbe, F.; Sidot, E.; Smyth, D.J.; Ohmoto, M.; Matsumoto, I.; Dardalhon, V.; Cesses, P.; Garnier, L.; Pouzolles, M.; Brulin, B.; et al. Intestinal Epithelial Tuft Cells Initiate Type 2 Mucosal Immunity to Helminth Parasites. Nature 2016, 529, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.C.; Costa, A.R.; Gonçalves, I.; Quintela, T.; Preissner, R.; Santos, C.R.A. The Druggability of Bitter Taste Receptors for the Treatment of Neurodegenerative Disorders. Biochem. Pharmacol. 2022, 197, 114915. [Google Scholar] [CrossRef]
- Klomparens, E.A.; Ding, Y. The Neuroprotective Mechanisms and Effects of Sulforaphane. Brain Circ. 2019, 5, 74. [Google Scholar] [CrossRef]
- Xiang, J.; Alesi, G.N.; Zhou, N.; Keep, R.F. Protective Effects of Isothiocyanates on Blood-CSF Barrier Disruption Induced by Oxidative Stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R1–R7. [Google Scholar] [CrossRef]
- Subedi, L.; Venkatesan, R.; Kim, S.Y. Neuroprotective and Anti-Inflammatory Activities of Allyl Isothiocyanate through Attenuation of JNK/NF-ΚB/TNF-α Signaling. Int. J. Mol. Sci. 2017, 18, 1423. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Vrontakis, M.; Parkinson, F.; Chelikani, P. Functional Bitter Taste Receptors Are Expressed in Brain Cells. Biochem. Biophys. Res. Commun. 2011, 406, 146–151. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, T.; Mao, L.; Zhang, F. Sulforaphane Protects against Brain Diseases: Roles of Cytoprotective Enzymes. Austin J. Cerebrovasc. Dis. Stroke 2017, 4, 1054. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, A.; Jeffery, E.H.; Miller, M.J. Is Bitterness Only a Taste? The Expanding Area of Health Benefits of Brassica Vegetables and Potential for Bitter Taste Receptors to Support Health Benefits. Nutrients 2022, 14, 1434. https://doi.org/10.3390/nu14071434
Zhao A, Jeffery EH, Miller MJ. Is Bitterness Only a Taste? The Expanding Area of Health Benefits of Brassica Vegetables and Potential for Bitter Taste Receptors to Support Health Benefits. Nutrients. 2022; 14(7):1434. https://doi.org/10.3390/nu14071434
Chicago/Turabian StyleZhao, Anqi, Elizabeth H. Jeffery, and Michael J. Miller. 2022. "Is Bitterness Only a Taste? The Expanding Area of Health Benefits of Brassica Vegetables and Potential for Bitter Taste Receptors to Support Health Benefits" Nutrients 14, no. 7: 1434. https://doi.org/10.3390/nu14071434