
Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review

 , , , , and
, , , , and 
Abstract
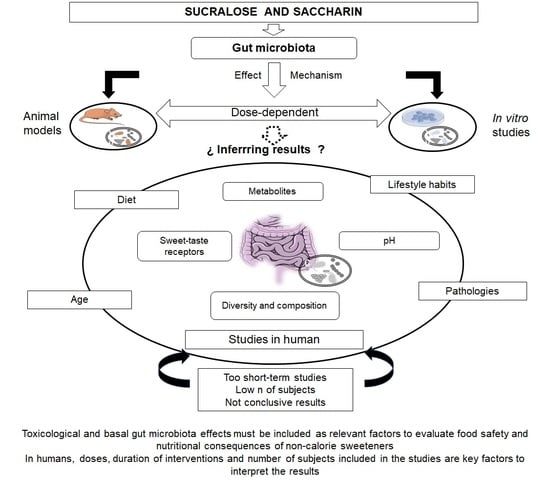
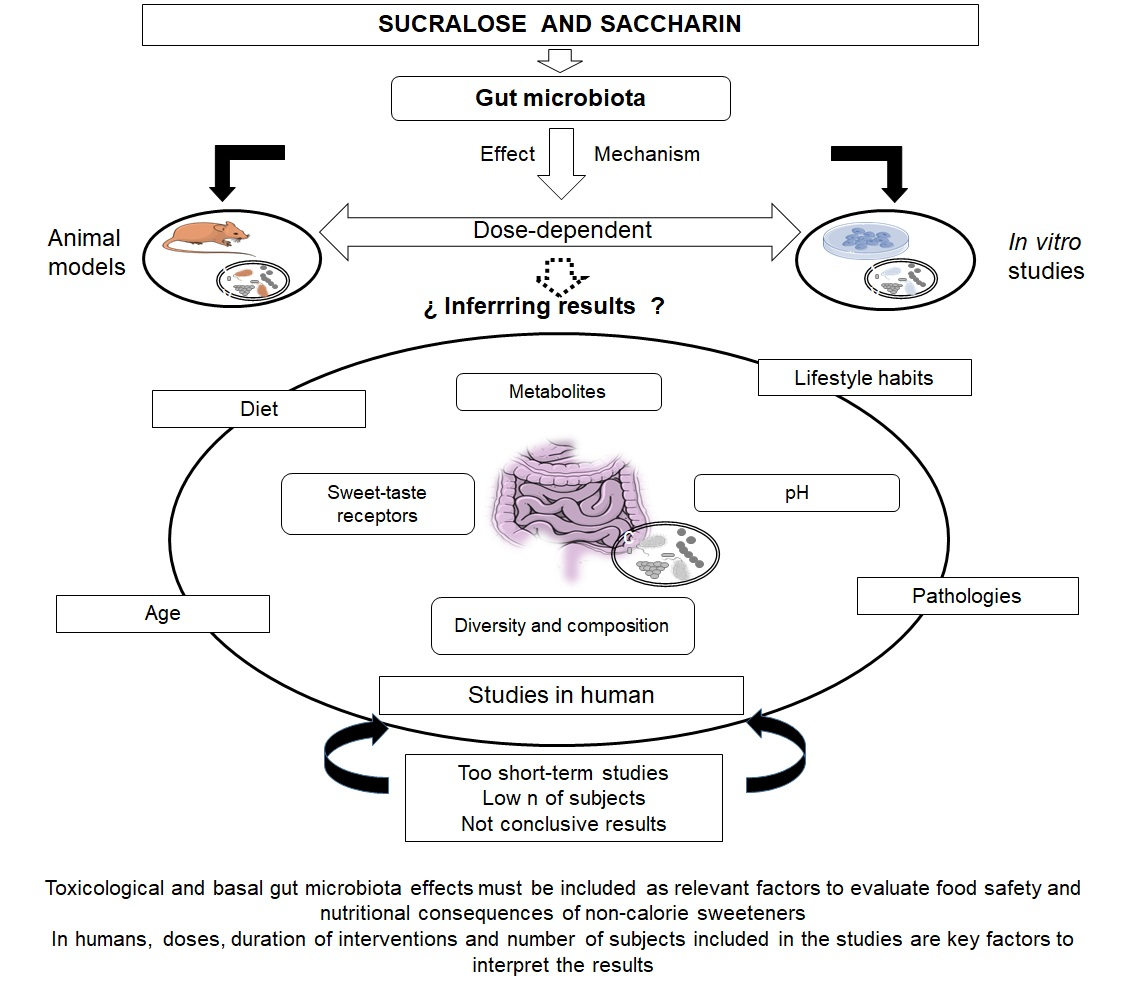
:
1. Introduction
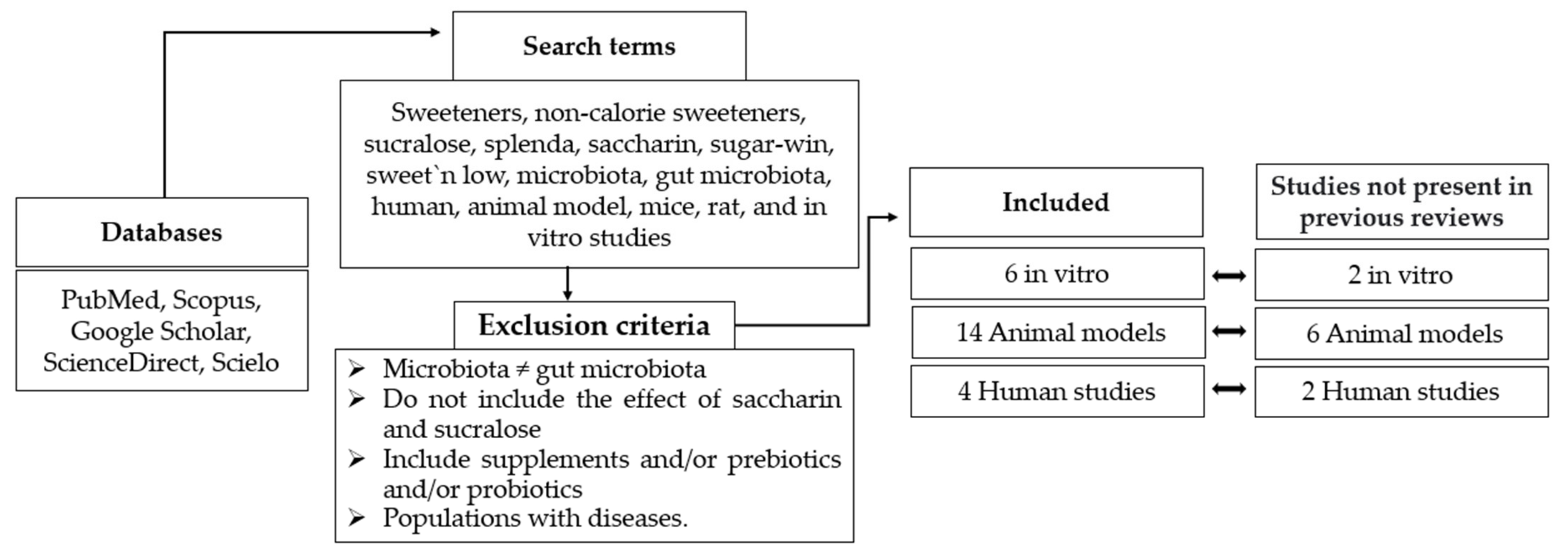
2. Materials and Methods
2.1. Effects of Sweeteners on the Gut Microbiota: In Vitro Trials
2.2. Effects of Sweeteners on the Gut Microbiota in Animal Models
2.3. Effects of Sweeteners on the Gut Microbiota in Human Trials
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Ingesta de Azúcares Para Adultos y Niños; Organización Mundial de la Salud: Geneva, Switzerland, 2015. [Google Scholar]
- Ruiz-Ojeda, F.J.; Plaza-Díaz, J.; Sáez-Lara, M.J.; Gil, A. Effects of Sweeteners on the Gut Microbiota: A Review of Experimental Studies and Clinical Trials. Adv. Nutr. 2019, 10, S31–S48. [Google Scholar] [CrossRef] [Green Version]
- Sylvetsky, A.C.; Jin, Y.; Clark, E.J.; Welsh, J.A.; Rother, K.I.; Talegawkar, S.A. Consumption of Low-Calorie Sweeteners among Children and Adults in the United States. J. Acad. Nutr. Diet 2017, 117, 441–448.e2. [Google Scholar] [CrossRef] [PubMed]
- Dunford, E.K.; Taillie, L.S.; Miles, D.R.; Eyles, H.; Tolentino-Mayo, L.; Ng, S.W. Non-Nutritive Sweeteners in the Packaged Food Supply—An Assessment across 4 Countries. Nutrients 2018, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- Samaniego-Vaesken, M.D.L.; González-Fernández, B.; Partearroyo, T.; Urrialde, R.; Varela-Moreiras, G. Updated Database and Trends of Declared Low- and No-Calorie Sweeteners from Foods and Beverages Marketed in Spain. Front. Nutr. 2021, 8, 670422. [Google Scholar] [CrossRef] [PubMed]
- EUR-Lex-32011R1129-EN-EUR-Lex. Available online: https://eur-lex.europa.eu/legal-content/ES/ALL/?uri=CELEX:32011R1129 (accessed on 13 January 2022).
- Pepino, M.Y. Metabolic effects of non-nutritive sweeteners. Physiol. Behav. 2015, 152, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooradian, A.D. In search for an alternative to sugar to reduce obesity. Int. J. Vitam. Nutr. Res. 2019, 89, 113–117. [Google Scholar] [CrossRef]
- Barber, T.; Valsamakis, G.; Mastorakos, G.; Hanson, P.; Kyrou, I.; Randeva, H.; Weickert, M. Dietary Influences on the Microbiota–Gut–Brain Axis. Int. J. Mol. Sci. 2021, 22, 3502. [Google Scholar] [CrossRef]
- Lobach, A.R.; Roberts, A.; Rowland, I.R. Assessing the in vivo data on low/no-calorie sweeteners and the gut microbiota. Food Chem. Toxicol. 2018, 124, 385–399. [Google Scholar] [CrossRef]
- Daly, K.; Darby, A.C.; Shirazi-Beechey, S.P. Low calorie sweeteners and gut microbiota. Physiol. Behav. 2016, 164, 494–500. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Gasbarrini, A.; Mele, M.C. Food Additives, Gut Microbiota, and Irritable Bowel Syndrome: A Hidden Track. Int. J. Environ. Res. Public Health 2020, 17, 8816. [Google Scholar] [CrossRef]
- Iebba, V.; Totino, V.; Gagliardi, A.; Santangelo, F.; Cacciotti, F.; Trancassini, M.; Mancini, C.; Cicerone, C.; Corazziari, E.; Pantanella, F.; et al. Eubiosis and Dysbiosis: The Two Sides of the Microbiota SuMMAry. New Microbiol. 2016, 39, 1–12. [Google Scholar] [PubMed]
- Redondo-Useros, N.; Nova, E.; González-Zancada, N.; Díaz, L.E.; Gómez-Martínez, S.; Marcos, A. Microbiota and Lifestyle: A Special Focus on Diet. Nutrients 2020, 12, 1776. [Google Scholar] [CrossRef] [PubMed]
- Qin, X. Letter: Non-Nutritive Sweeteners May Have a Bell-Shaped Dose Response for Dysbiosis-Related Adverse Effects That Contributed to Discrepancies between Prospective Cohort and Randomized Controlled Studies. J. Obes. Metab. Syndr. 2020, 29, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef] [PubMed]
- Rosés, C.; Cuevas-Sierra, A.; Quintana, S.; Riezu-Boj, J.; Martínez, J.; Milagro, F.; Barceló, A. Gut Microbiota Bacterial Species Associated with Mediterranean Diet-Related Food Groups in a Northern Spanish Population. Nutrients 2021, 13, 636. [Google Scholar] [CrossRef]
- Boulangé, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.-E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Merra, G.; Noce, A.; Marrone, G.; Cintoni, M.; Tarsitano, M.G.; Capacci, A.; De Lorenzo, A. Influence of Mediterranean Diet on Human Gut Microbiota. Nutrients 2020, 13, 7. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Carocho, M.; Morales, P.; Ferreira, I.C.F.R. Sweeteners as food additives in the XXI century: A review of what is known, and what is to come. Food Chem. Toxicol. 2017, 107, 302–317. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Pastor-Villaescusa, B.; Rueda, A.; Abadia-Molina, F.; Ruiz-Ojeda, F.J. Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies. Nutrients 2020, 12, 1153. [Google Scholar] [CrossRef]
- FDA. Additional Information about High-Intensity Sweeteners Permitted for Use in Food in the United States. Available online: https://www.fda.gov/food/food-additives-petitions/additional-information-about-high-intensity-sweeteners-permitted-use-food-united-states (accessed on 19 November 2021).
- Magnuson, B.A.; Carakostas, M.C.; Moore, N.H.; Poulos, S.; Renwick, A.G. Biological fate of low-calorie sweeteners. Nutr. Rev. 2016, 74, 670–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO/JECFA. Saccharin. Available online: https://apps.who.int/food-additives-contaminants-jecfa-database/Home/Chemical/3164 (accessed on 7 April 2022).
- Administración Nacional de Medicamentos, Alimentos y Tecnología Médica. (A.N.M.A.T.). Código Alimentario Argentino. Alimentos régimen o dietéticos (2021). Available online: http://www.anmat.gov.ar/resultados.asp?cx=018082787451070703178%3Arx-vbt5pdfu&cof=FORID%3A10&ie=UTF-8&q=CAPITULO_XVII.pdf (accessed on 16 March 2022).
- EUFIC. What Is an Acceptable Daily Intake (ADI)? Available online: https://www.eufic.org/en/understanding-science/article/qas-on-acceptable-daily-intakes-adis (accessed on 14 January 2022).
- WHO/JECFA. Sucralose. Available online: https://apps.who.int/food-additives-contaminants-jecfa-database/Home/Chemical/2340 (accessed on 7 April 2022).
- EFSA. Safety of the proposed extension of use of sucralose (E 955) in foods for special medical purposes in young children. EFSA J. 2016, 14, e04361. [Google Scholar] [CrossRef]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; de Bastos, M.L.; Bories, G.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; Kolar, B.; et al. Safety and Efficacy of Sodium Saccharin When Used as a Feed Flavour for Piglets, Pigs for Fattening, Calves for Rearing and Calves for Fattening. EFSA J. 2018, 16, e05208. [Google Scholar] [CrossRef]
- Diabetes Canada. Sweeteners that increase blood sugar levels. 2018. Available online: https://guidelines.diabetes.ca/docs/patient-resources/sugars-and-sweeteners.pdf (accessed on 28 January 2022).
- Abou-Donia, M.B.; El-Masry, E.M.; Abdel-Rahman, A.A.; McLendon, R.E.; Schiffman, S.S. Splenda Alters Gut Microflora and Increases Intestinal P-Glycoprotein and Cytochrome P-450 in Male Rats. J. Toxicol. Environ. Health Part A 2008, 71, 1415–1429. [Google Scholar] [CrossRef]
- Anderson, R.; Kirkland, J. The effect of sodium saccharin in the diet on caecal microflora. Food Cosmet. Toxicol. 1980, 18, 353–355. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Ministerio de Agricultura, Pesca y Alimentación. Informe del Consumo Alimentario en España. 2020. Available online: https://www.mapa.gob.es/es/alimentacion/temas/consumo-tendencias/informe-anual-consumo-2020-v2-nov2021-baja-res_tcm30-562704.pdf (accessed on 15 February 2022).
- Bachmanov, A.A.; Reed, D.R.; Beauchamp, G.K.; Tordoff, M.G. Food Intake, Water Intake, and Drinking Spout Side Preference of 28 Mouse Strains. Behav. Genet. 2002, 32, 435. [Google Scholar] [CrossRef]
- John Hopkin University Species Specific Information: Rat. Available online: https://web.jhu.edu/animalcare/procedures/rat.html#general (accessed on 1 March 2022).
- Harpaz, D.; Yeo, L.P.; Cecchini, F.; Koon, T.H.P.; Kushmaro, A.; Tok, A.I.Y.; Marks, R.S.; Eltzov, E. Measuring Artificial Sweeteners Toxicity Using a Bioluminescent Bacterial Panel. Molecules 2018, 23, 2454. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.-P.; Browman, D.; Herzog, H.; Neely, G.G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef]
- Markus, V.; Share, O.; Shagan, M.; Halpern, B.; Bar, T.; Kramarsky-Winter, E.; Teralı, K.; Özer, N.; Marks, R.S.; Kushmaro, A.; et al. Inhibitory Effects of Artificial Sweeteners on Bacterial Quorum Sensing. Int. J. Mol. Sci. 2021, 22, 9863. [Google Scholar] [CrossRef]
- Gerasimidis, K.; Bryden, K.; Chen, X.; Papachristou, E.; Verney, A.; Roig, M.; Hansen, R.; Nichols, B.; Papadopoulou, R.; Parrett, A. The impact of food additives, artificial sweeteners and domestic hygiene products on the human gut microbiome and its fibre fermentation capacity. Eur. J. Nutr. 2019, 59, 3213–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shil, A.; Chichger, H. Artificial Sweeteners Negatively Regulate Pathogenic Characteristics of Two Model Gut Bacteria, E. coli and E. faecalis. Int. J. Mol. Sci. 2021, 22, 5228. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E.; Pelinescu, D.; Gatea, F.; Sârbu, I. Altered in Vitro Metabolomic Response of the Human Microbiota to Sweeteners. Genes 2019, 10, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omran, A.; Baker, R.; Coughlin, C. Differential Bacteriostatic Effects of Sucralose on Various Species of Environmental Bacteria. Int. Sch. Res. Not. 2013, 2013, 415070. [Google Scholar] [CrossRef]
- Sünderhauf, A.; Pagel, R.; Künstner, A.; Wagner, A.E.; Rupp, J.; Ibrahim, S.M.; Derer, S.; Sina, C. Saccharin Supplementation Inhibits Bacterial Growth and Reduces Experimental Colitis in Mice. Nutrients 2020, 12, 1122. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Smith, K.R.; Crouch, A.L.; Sharma, V.; Yi, F.; Vargova, V.; LaMoia, T.E.; Dupont, L.M.; Serna, V.; Tang, F.; et al. High-dose saccharin supplementation does not induce gut microbiota changes or glucose intolerance in healthy humans and mice. Microbiome 2021, 9, 11. [Google Scholar] [CrossRef]
- Falcon, T.; Foletto, K.C.; Siebert, M.; Pinto, D.E.; Andrades, M.; Bertoluci, M.C. Metabarcoding reveals that a non-nutritive sweetener and sucrose yield similar gut microbiota patterns in Wistar rats. Genet. Mol. Biol. 2020, 43, e20190028. [Google Scholar] [CrossRef]
- Uebanso, T.; Ohnishi, A.; Kitayama, R.; Yoshimoto, A.; Nakahashi, M.; Shimohata, T.; Mawatari, K.; Takahashi, A. Effects of Low-Dose Non-Caloric Sweetener Consumption on Gut Microbiota in Mice. Nutrients 2017, 9, 560. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Tapia, M.; Miller, A.W.; Granados-Portillo, O.; Tovar, A.R.; Torres, N. The development of metabolic endotoxemia is dependent on the type of sweetener and the presence of saturated fat in the diet. Gut Microbes 2020, 12, 1840766. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, J.; Yang, M.; Qian, C.; Liu, Y.; Qi, Y.; Feng, R.; Yang, M.; Liu, W.; Ma, J. Low Doses of Sucralose Alter Fecal Microbiota in High-Fat Diet-Induced Obese Rats. Front. Nutr. 2021, 8, 787055. [Google Scholar] [CrossRef]
- Li, J.; Zhu, S.; Lv, Z.; Dai, H.; Wang, Z.; Wei, Q.; Hamdard, E.; Mustafa, S.; Shi, F.; Fu, Y. Drinking Water with Saccharin Sodium Alters the Microbiota-Gut-Hypothalamus Axis in Guinea Pig. Animals 2021, 11, 1875. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut Microbiome Response to Sucralose and Its Potential Role in Inducing Liver Inflammation in Mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, X.; Tu, P.; Chi, L.; Gao, B.; Ru, H.; Lu, K. Saccharin induced liver inflammation in mice by altering the gut microbiota and its metabolic functions. Food Chem. Toxicol. 2017, 107, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Guo, Z.; Chen, D.; Li, L.; Song, X.; Liu, T.; Jin, G.; Li, Y.; Liu, Y.; Ajiguli, A.; et al. Maternal sucralose intake alters gut microbiota of offspring and exacerbates hepatic steatosis in adulthood. Gut Microbes 2020, 11, 1043–1063. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.B.; Krieger, K.; Khan, F.M.; Huffman, W.; Chang, M.; Naik, A.; Yongle, R.; Hameed, I.; Krieger, K.; Girardi, L.N.; et al. The current state of animal models in research: A review. Int. J. Surg. 2019, 72, 9–13. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Boets, E.; Gomand, S.V.; Deroover, L.; Preston, T.; Vermeulen, K.; De Preter, V.; Hamer, H.M.; Van den Mooter, G.; De Vuyst, L.; Courtin, C.M.; et al. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: A stable isotope study. J. Physiol. 2017, 595, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, W.; Rył, A.; Mizerski, A.; Walczakiewicz, K.; Sipak, O.; Laszczyńska, M. Immunomodulatory potential of gut microbiome-derived short-chain fatty acids (SCFAs). Acta Biochim. Pol. 2019, 66, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Friel, J.; Mackay, D. The Effects of Non-Nutritive Artificial Sweeteners, Aspartame and Sucralose, on the Gut Microbiome in Healthy Adults: Secondary Outcomes of a Randomized Double-Blinded Crossover Clinical Trial. Nutrients 2020, 12, 3408. [Google Scholar] [CrossRef]
- Thomson, P.; Santibañez, R.; Aguirre, C.; Galgani, J.E.; Garrido, D. Short-term impact of sucralose consumption on the metabolic response and gut microbiome of healthy adults. Br. J. Nutr. 2019, 122, 856–862. [Google Scholar] [CrossRef] [Green Version]
- Aldrete-Velasco, J.; López-García, R.; Zúñiga-Guajardo, S.; Riobó-Serván, P.; Serra-Majem, L.; Suverza-Fernández, A.; Esquivel-Flores, M.; MOlina-Segui, F.; Pedroza-Islas, R.; Rascón-Hernández, M.; et al. Análisis de La Evidencia Disponible Para El Consumo de Edulcorantes No Calóricos. Documento de Expertos. Med. Interna México 2017, 33, 61–83. [Google Scholar]
- Schiffman, S.S.; Nagle, H.T. Revisited: Assessing the in vivo data on low/no-calorie sweeteners and the gut microbiota. Food Chem. Toxicol. 2019, 132, 110692. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| JECFA ADI [25,28] | EFSA ADI [29,30] | Health Canada | [31] ANMAT [26] | |
|---|---|---|---|---|
| Saccharin | 5 | 5 | 5 | 2.5 |
| Sucralose | 15 | 15 | 9 | 15 |
| Saccharin | Sucralose | |
|---|---|---|
| ADI mg/kg body wt (JECFA) | 5 mg/kg | 15 mg/kg |
| ADI subject 70 kg | 350 mg | 1050 mg |
| Average consumption of the Spanish population | 300 mg/day | |
| Reference | Sweeteners/Doses/Duration | Methods | Bacteria | Results/Conclusions Saccharine and/or Sucralose |
|---|---|---|---|---|
| Harpaz et al., 2018 [38] | Aspartame, sucralose, saccharine, neotame, advantame, and acesulfame potassium-k (ace-k). ADI (FDA) | Bioluminescent | E. coli strains (TV1061, DPD2544 and DPD2794) | Toxic effects |
| Wang et al., 2018 [39] | Sucralose, saccharin, acesulfame potassium, and rebaudioside Liquid assay: equal molarity of sodium chloride/5 h Agar: 1.25% (w/v) sucralose and 2.5% (w/v) sucralose/24 h | Liquid culture assay. LB agar plate assay | E. coli HB101 and E. coli K-12 | Bacteriostatic effects |
| Markus V, et al., 2021 [40] | Aspartame, sucralose, saccharin Bioluminescence assay, growth assay: 10 µL non-calorie sweeteners or sports supplements. Swarming motility assay: aspartame (1.36 mM), sucralose (25.2 mM), or saccharine (2.72 mM) QS competition assay using Chromobacterium Violaceum CV026/20 h | Biosensor assays, biophysical protein characterization methods, microscale thermophoresis, swarming motility assays, growth assays, and molecular docking | E coli K802NR and P. aeruginosa lasRI P. aeruginosa PAO1 C. violaceum (CV026) | Inhibition of quorum sensing |
| Gerasimidis C et al., 2020 [41] | Aspartame-based sweetener, sucralose, stevia 50% ADI (male, w: 75 kg) | Gas chromatography | Total bacteria (feces from healthy individuals) and 5 bacterial groups (Bacteroides/Prevotella, Bifidobacterium, B. coccoides, C. leptum and E. coli) | Sucralose: shifted microbiome community structure ↔ bacterial populations ↑ Escherichia/Shigella |
| Shil A and Chichger, H, 2021 [42] | Saccharin, sucralose, and aspartameGrowth curve: 0.1 to 1000 µM/4 d Biofilm formation assay: 100 µM/48 h Haemolysis assay, adhesion assay, and invasion assay: 100 M/24 h Cytotoxicity assay: 100 M/48 h | Models of microbiota and the intestinal epithelium (Caco-2 cells) | E. coli NCTC10418 and E. faecalis ATCC19433 S. aureus | Saccharin bacteriostatic effects Saccharin, sucralose: ↑ biofilm formation ↑ ability of bacteria to adhere to, invade, and kill gut epithelial cells (exception saccharin on E. coli) Negative effect on intestinal epithelial cell apoptosis and permeability |
| Vamanu E et al., 2019 [43] | Sodium cyclamate, sucralose, sodium saccharin, steviol, white sugar 40 mg active substance (more than 90% purity) | Static GIS1 simulator (three segments of the human colon) | Total microbial (feces from healthy individuals) | Saccharin: ↓ number of microorganisms; ↓ SCFAs Both: ↓ phylum Firmicutes; ↓ fermentative processes; ↑ colonic pH; ↑ 10% ammonia synthesized; ↓ SCFAs |
| Reference | Sweeteners/Doses/Duration | Animal Model | Results |
|---|---|---|---|
| Anderson & Kirkland, 1980 [33] | Treatment: 7.5% sodium saccharin in the Purina laboratory chow Control: Cellulose 7.5% in the Purina laboratory chow Duration: 10 d | Weaning male Charles River rats (Weight 55 ± 3 g) (n = 7) | ↑ The numbers of aerobic microbes ↓ Anaerobic/aerobic ratio |
| Serrano et al., 2021 [46] | Treatment: saccharin average daily dose equal to 4 times (250 mg/kg) the human ADI (JECFA) Control: water Duration: 10 wk | 8-wk-old mice | ↔Alpha and beta diversity and relative microbial abundances |
| Falcon et al., 2020 [47] | Control: Sucrose-sweetened yogurt (suc): low-fat yogurt supplemented with 20% sucrose, final solution concentration 11.4% sucrose Treatment: NNS-supplemented yogurt: (0.3% sodium saccharin and sodium cyclamate). Final solution concentration 0.17% NNS Duration: 17 wk | Adult male Wistar rats (weight: 210 ± 6 g) SUC (n = 9 per group) NNS (n = 10 per group) | ↔Species richness ↔ Shannon or Simpson diversity indices |
| Abou-Donia et al., 2008 [32] | Treatment: Splenda (Sucralose) oral gavage: 1.1; 3.3; 5.5 and 11 mg/kg/d sucralose concentrations. Control: water Duration: 12 wk | Male Sprague-Dawley rats (weight: 200–240 g) (n = 10 per group) | ↓ Number of total anaerobes and other anaerobic bacteria (Bifidobacteria, Lactobacilli, Bacteroides, and Clostridium). |
| Uebanso et al., 2017 [48] | Treatment: LS (sucralose solution of 1.5 mg/kg bw/d). HS (sucralose solution of 15 mg/kg bw/d), which is equal to the maximum ADI. Control: distilled water Duration: 8 wk | Male and female C57Bl/6 J mice (4 wk old) (n = 8) | LS vs. HS ↔The relative amounts of fecal total bacteria LS vs. HS ↔ Firmicutes and Bacteroidetes phylum bacteria ↓ relative Clostridium cluster XIVa, dose-dependent |
| Sánchez-Tapia et al., 2020 [49] | Treatment: Sucralose: drinking water 1.5% sucralose Control: water Duration: 4 mo | Male Wistar rats (5 wk old) (n = 6 per group) | ↓α-diversity ↑ B. fragilis abundance |
| Wang et al., 2018 [39] | Treatment: Sucralose: drinking water sucralose (2.5%, w/v) Duration: 8 wk | C57BL/6 mice (5 wk old) | ↔ α-diversity, Actinobacteria, and Proteobacteria ↑ Abundance of Firmicutes |
| Zhang et al., 2021 [50] | Treatment: daily gavage of Sucralose ∼ 0.43 mg, sucralose ~0.62 mg. Control: daily gavage of 2 mL normal saline Duration: 4 wk | Obese Sprague Dawley rats (4 wk old) (8 weeks after high fat diet (HFD)) (n = 6 per group) | 0.43 mg sucralose: ↑ relative abundance of Firmicutes and ↓ Bacteroidetes 0.62 mg sucralose: ↓ relative abundance of Firmicutes ↑ Bacteroidetes The ratio of Firmicutes to Bacteroidetes in 0.43 mg sucralose was higher than that in 0.62 mg |
| Li et al., 2021 [51] | Treatment: Saccharin sodium in drinking water: 1.5 mM Control: water Duration: 4 wk | Female Harley-white guinea pigs (Cavia porcellus) (4 wk old) (weight: 240.7 ± 7.7 g) (n= 6 per group) | ↑ Firmicutes and Lactobacillasceae-Lactobacillus abundance ↓ Relative abundance of Erysipelotrichaceae, Eubacteriaceae, and Ileibacterium |
| Bian et al., 2017 [52] | Treatment: Sucralose tap water (0.1 mg/mL). ADI (FDA) Control: tap water Duration: 6 mo | C57BL/6 male mice (~8 wk old) (n = 10 per group) | ↑Numerous bacterial toxin genes (toxic shock syndrome toxin-1 and shiga toxin subunits) 14 genera exhibited different patterns over time in sucralose, different after 3 and/or 6 mo of treatment |
| Bian et al., 2017 [53] | Treatment: Saccharin, drinking water (0.3 mg/mL). ≈ ADI (FDA) Control: tap water Duration: 6 mo | C57BL/6 J male mice (Weight, ~23 g, ~8 wk old) (n = 10 per group) | Alterations of the gut metabolome with 1743 significant changes in molecular features 3 mo: ↑Sporosarcina, Jeotgalicoccus, Akkermansia, Oscillospira, and Corynebacterium ↓Anaerostipes and Ruminococcus 6 mo: ↑Corynebacterium, Roseburia, and Turicibacter ↓Ruminococcus, Adlercreutzia, and Dorea |
| Suez et al., 2014 [34] | Treatment: Commercial NAS in drinking water 10% solution: (5% saccharin, 95% glucose), (5% Sucralose), (4% Aspartame). Pure saccharin (0.1 mg ml−1) in drinking Control: water or water with 10% glucose or 10% sucrose Duration: 11 wk NAS and 5 wk pure saccharin | Lean C57Bl/6 mice (10 wk old) with NAS treatment (n = 5 per group) C57Bl/6 mice fed on HFD with saccharin treatment (10 wk old) (n = 8 per group) | Saccharin: dysbiosis reflected by more than 40 operational taxonomic units (OTUs) abundances changed ↑ Bacteroides genus and Clostridiales order Dysbiosis in mice that consumed pure saccharin and HFD |
| Dai et al., 2020 [54] | MS treatment: gestation and lactation, sucralose 0.1 mg/mL (FDA ADI) Offspring treatment: weaned pups fed a control diet until 8 wk of age and treated with HDF for 4 wk Control: distilled water in MS maternal control and offspring fed with a control diet Duration: maternal treatment, 6 wk | C57BL/6 pregnant mice 3 wk old, weaned pups | MS: at phylum level ↑ the relative abundance of Verrucomicrobia and Proteobacteria and ↓Bacteroidetes At genus level ↑ abundance of Akkermansia, Blautia, Corynebacterium, Robinsoniella, and ↓ Alistipes, Barnesiella, Paraprevotella, Saccharibacteria genera incertae sedis, and Streptococcus MS alters the gut microbiota in the offspring, ↓alpha diversity of 3-wk-old pups |
| Reference | Sweeteners/Doses/Duration | Design | Results/Conclusions Saccharin and Sucralose |
|---|---|---|---|
| Serrano et al., 2021 [46] | Saccharin 400 mg/d/2 wk | Randomized, double-blind, placebo-controlled interventional study | ↔gut microbiota |
| Ahmad et al., 2020 [59] | Sucralose and aspartame 20% ADI sucralose (~0.136 g sucralose)/14 d | Randomized, double-blind crossover (12 wk) and controlled clinical trial. | ↔ gut microbiota ↔ SCFAs |
| Thomson et al., 2019 [60] | Sucralose 780 mg/d/7 d | Randomized, double-blind study | ↔ gut microbiota |
| Suez et al., 2014 [34] | Saccharin FDA maximal ADI/7 d | Intervention study | Response according to basal microbiota |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
del Pozo, S.; Gómez-Martínez, S.; Díaz, L.E.; Nova, E.; Urrialde, R.; Marcos, A. Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review. Nutrients 2022, 14, 1682. https://doi.org/10.3390/nu14081682
del Pozo S, Gómez-Martínez S, Díaz LE, Nova E, Urrialde R, Marcos A. Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review. Nutrients. 2022; 14(8):1682. https://doi.org/10.3390/nu14081682
Chicago/Turabian Styledel Pozo, Susana, Sonia Gómez-Martínez, Ligia E. Díaz, Esther Nova, Rafael Urrialde, and Ascensión Marcos. 2022. "Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review" Nutrients 14, no. 8: 1682. https://doi.org/10.3390/nu14081682
APA Styledel Pozo, S., Gómez-Martínez, S., Díaz, L. E., Nova, E., Urrialde, R., & Marcos, A. (2022). Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review. Nutrients, 14(8), 1682. https://doi.org/10.3390/nu14081682