
The Protective Effect of Heat-Inactivated Companilactobacillus crustorum on Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Preparation of HICC
2.2. Animals and Treatment
2.3. The Measurement of Disease Activity Index (DAI)
2.4. Histological Analysis
2.5. Immunofluorescence Assay
2.6. RT-qPCR Assay
2.7. The Analysis of Gut Microbiota
2.8. The Determination of Biochemical Parameters
2.9. Statistical Analyses
3. Results
3.1. HICC Alleviated the Pathological Symptoms in DSS-Induced UC Mice
3.2. HICC Ameliorated Intestinal Inflammation in Mice
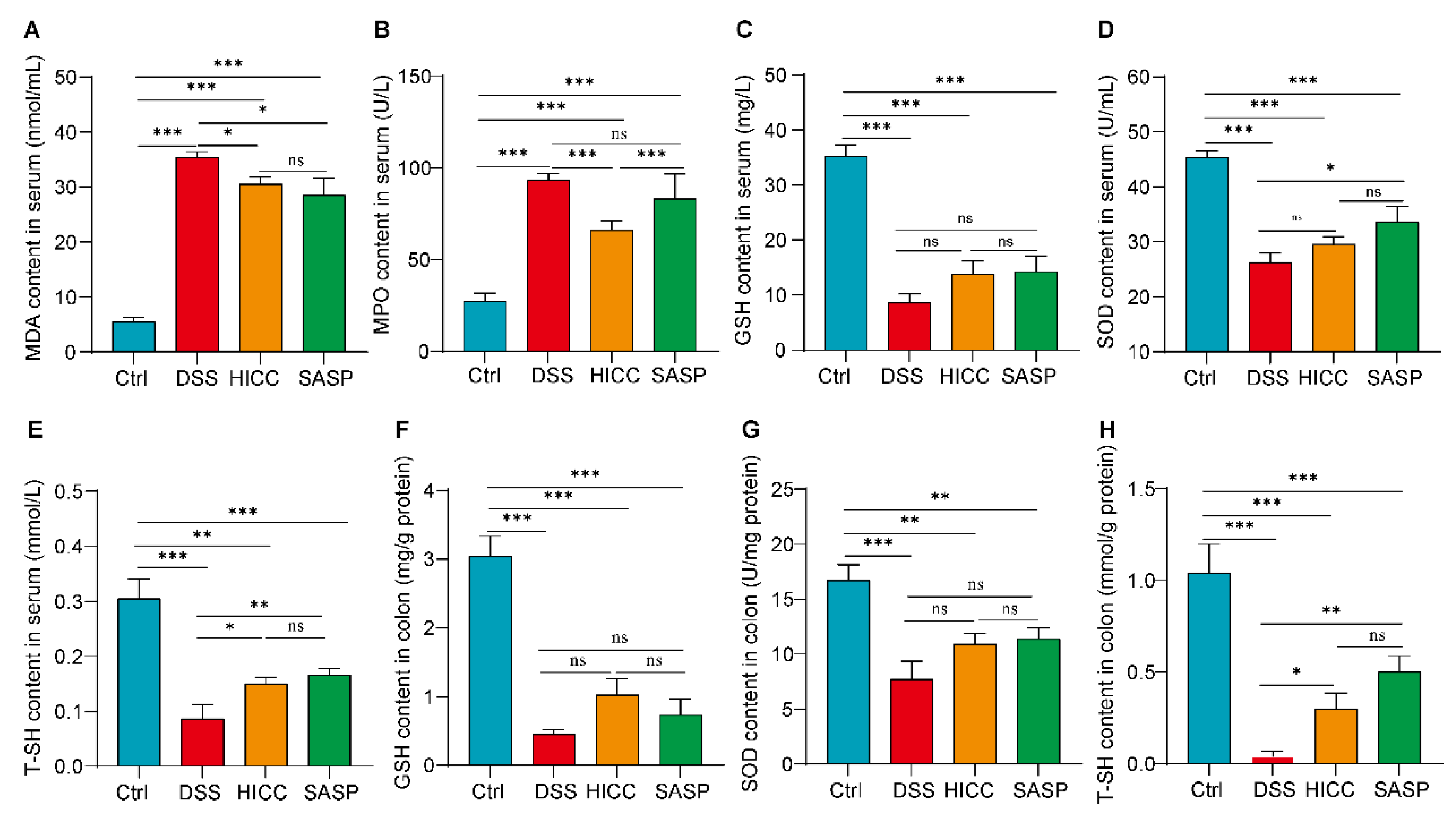
3.3. HICC Attenuated the Oxidative Damage in DSS-Induced UC Mice
3.4. HICC Mitigated Intestinal Barrier Injury in DSS-Induced UC Mice
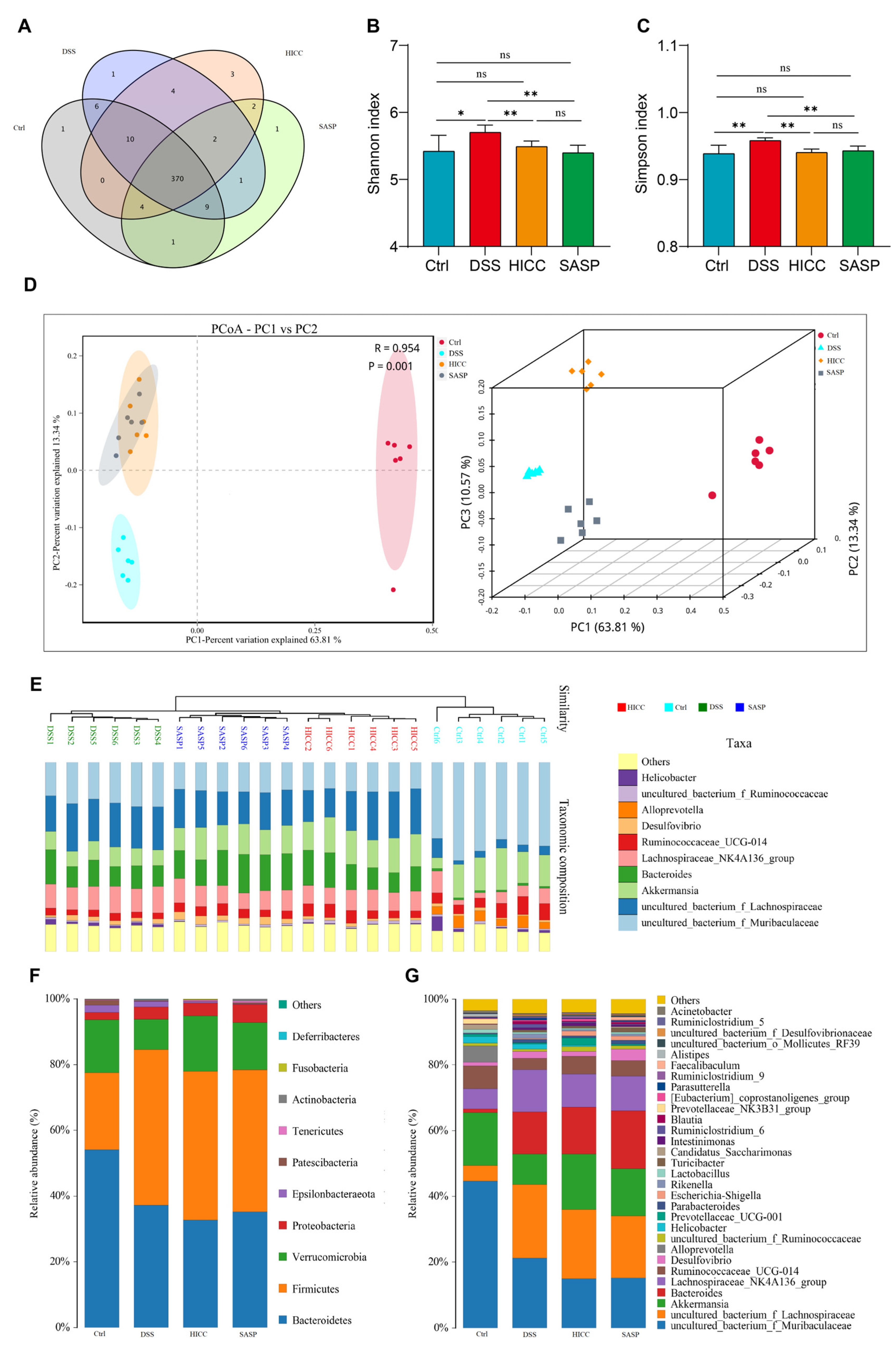
3.5. HICC Modulated Intestinal Microbiota Structure in DSS-Induced UC Mice
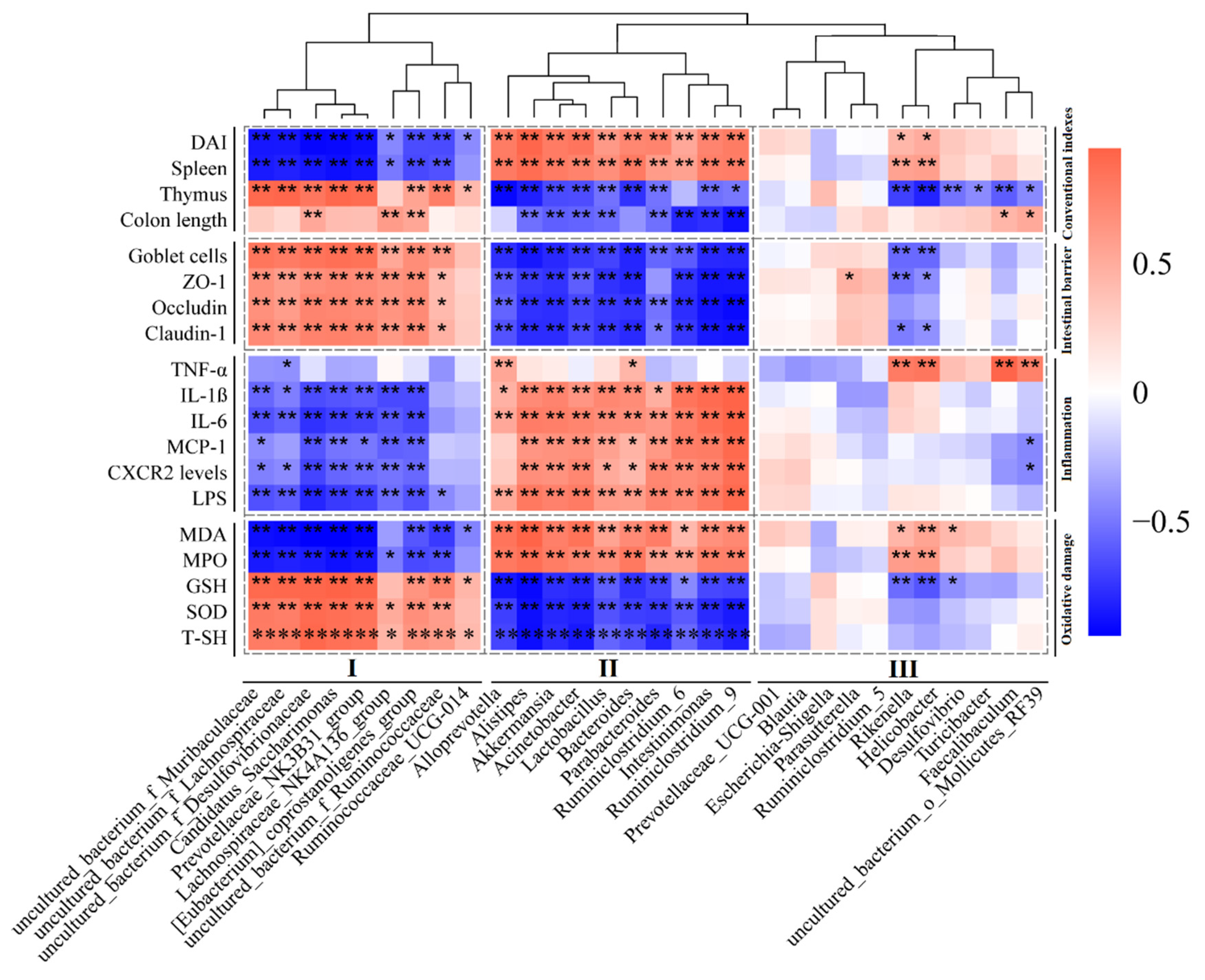
3.6. The Relationship between the Gut Microbiota and UC-Related Pathological Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Du, L.L.; Ha, C. Epidemiology and pathogenesis of ulcerative colitis. Gastroenterol. Clin. N. Am. 2020, 49, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Damas, O.M.; Abreu, M.T. Are patients with ulcerative colitis still at increased risk of colon cancer? Lancet 2020, 395, 92–94. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Shen, Z.F.; Wu, B.S.; Xu, C.B.; He, Z.Q.; Chen, T.; Shang, H.T.; Xie, C.F.; Huang, S.Y.; Chen, Y.G.; et al. Risk of colorectal cancer in ulcerative colitis patients: A systematic review and meta-analysis. Gastroenterol. Res. Pract. 2019, 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonu, I.; Lin, M.V.; Blonski, W.; Lichtenstein, G.R. Clinical pharmacology of 5-ASA compounds in inflammatory bowel disease. Gastroenterol. Clin. N. Am. 2010, 39, 559–599. [Google Scholar] [CrossRef]
- Schauer, C.; Avery, V.; Seleq, S.; Garg, P.; Wang, M.T.M.; Chieng, M.; Rowan, C.; Sekra, A.; Lane, M.; Walmsley, R. A comparison of intravenous methylprednisolone and hydrocortisone for the treatment of acute inflammatory bowel disease. J. Gastroenterol. Hepatol. 2021, 36, 2762–2768. [Google Scholar] [CrossRef]
- Parlak, E.; Dagli, Ü.; Ülker, A.; Alkim, C.; Sahin, B. Comparison of 5-amino salicylic acid plus glucocorticosteroid with metronidazole and ciprofloxacin in patients with active ulcerative colitis. J. Clin. Gastroenterol. 2001, 33, 85–86. [Google Scholar] [CrossRef]
- Porter, R.J.; Kalla, R.; Ho, G.-T. Ulcerative colitis: Recent advances in the understanding of disease pathogenesis. F1000Research 2020, 9, 1–13. [Google Scholar] [CrossRef]
- Hu, Y.; Ye, Z.; Wu, M.; She, Y.; Li, L.; Xu, Y.; Qin, K.; Hu, Z.; Yang, M.; Lu, F. The communication between intestinal microbiota and ulcerative colitis: An exploration of pathogenesis, animal models, and potential therapeutic strategies. Front. Med. 2021, 8, 766126. [Google Scholar] [CrossRef]
- Watson, A.J.M. Claudins and barrier dysfunction in intestinal inflammation: Cause or consequence? Gut 2015, 64, 1501-U1208. [Google Scholar] [CrossRef]
- Miner-Williams, W.M.; Moughan, P.J. Intestinal barrier dysfunction: Implications for chronic inflammatory conditions of the bowel. Nutr. Res. Rev. 2016, 29, 40–59. [Google Scholar] [CrossRef]
- Iheozor-Ejiofor, Z.; Kaur, L.; Gordon, M.; Baines, P.A.; Sinopoulou, V.; Akobeng, A.K. Probiotics for maintenance of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2020, 3, CD007443. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Dhaneshwar, S. Role of prebiotics, probiotics, and synbiotics in management of inflammatory bowel disease: Current perspectives. World J. Gastroenterol. 2023, 29, 2078–2100. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Jin, H.; Liao, N.; Li, J.J.; Jiang, C.M.; Shao, D.Y.; Shi, J.L. Lactobacillus rhamnosus from human breast milk ameliorates ulcerative colitis in mice via gut microbiota modulation. Food Funct. 2021, 12, 5171–5186. [Google Scholar] [CrossRef]
- Dong, F.; Xiao, F.F.; Li, X.L.; Li, Y.R.; Wang, X.F.; Yu, G.J.; Zhang, T.; Wang, Y.Z. Pediococcus pentosaceus CECT 8330 protects DSS-induced colitis and regulates the intestinal microbiota and immune responses in mice. J. Transl. Med. 2022, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.L.; Xu, Y.F.; Kong, X.W.; Wang, J.M.; Zha, X.D.; Wang, Y.Z. Probiotic Bacillus cereus alleviates dextran sulfate sodium-induced colitis in mice through improvement of the intestinal barrier function, anti-inflammation, and gut microbiota modulation. J. Agric. Food Chem. 2021, 69, 14810–14823. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Burgos, M.; Moreno-Fernandez, J.; Alferez, M.J.M.; Diaz-Castro, J.; Lopez-Aliaga, I. New perspectives in fermented dairy products and their health relevance. J. Funct. Foods 2020, 72, 104059. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 671. [Google Scholar] [CrossRef]
- Kye, Y.J.; Lee, S.Y.; Kim, H.R.; Lee, B.H.; Park, M.S.; Ji, G.E.; Sung, M.K. Lactobacillus acidophilus PIN7 paraprobiotic supplementation ameliorates DSS-induced colitis through anti-inflammatory and immune regulatory effects. J. Appl. Microbiol. 2022, 132, 3189–3200. [Google Scholar] [CrossRef]
- Jin, J.H.; Wu, S.Q.; Xie, Y.H.; Liu, H.; Gao, X.Z.; Zhang, H.X. Live and heat-killed cells of Lactobacillus plantarum Zhang-LL ease symptoms of chronic ulcerative colitis induced by dextran sulfate sodium in rats. J. Funct. Foods 2020, 71, 103994. [Google Scholar] [CrossRef]
- Wang, D.; Yin, C.; Ye, H.; Guo, Y. Protective effect of heat-killed Lactobacillus rhamnosus HN001 on dextran sulfate sodium induced colitis in mice. Food Ferment. Ind. 2021, 47, 30–35. [Google Scholar] [CrossRef]
- Wang, T.; Shi, C.H.; Wang, S.X.; Zhang, Y.; Wang, S.; Ismael, M.; Zhang, J.; Wang, X.; Lu, X. Protective effects of Companilactobacillus crustorum MN047 against dextran sulfate sodium-induced ulcerative colitis: A fecal microbiota transplantation study. J. Agric. Food Chem. 2022, 70, 1547–1561. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Sun, H.M.; Chen, J.X.; Luo, L.L.; Gu, Y.X.; Wang, X.; Shan, Y.Y.; Yi, Y.L.; Liu, B.F.; Zhou, Y.; et al. Anti-adhesion effects of Lactobacillus strains on Caco-2 cells against Escherichia coli and their application in ameliorating the symptoms of dextran sulfate sodium-induced colitis in mice. Probiotics Antimicrob. Proteins 2021, 13, 1632–1643. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yan, H.; Lu, Y.Y.; Li, X.; Wang, X.; Shan, Y.Y.; Yi, Y.L.; Liu, B.F.; Zhou, Y.; Lu, X. Anti-obesity effect of Lactobacillus rhamnosus LS-8 and Lactobacillus crustorum MN047 on high-fat and high-fructose diet mice base on inflammatory response alleviation and gut microbiota regulation. Eur. J. Nutr. 2020, 59, 2709–2728. [Google Scholar] [CrossRef]
- Wang, T.; Wang, P.P.; Ge, W.P.; Shi, C.; Xiao, G.N.; Wang, X.; Lu, X. Protective effect of a multi-strain probiotics mixture on azoxymethane/dextran sulfate sodium-induced colon carcinogenesis. Food Biosci. 2021, 44, 101346. [Google Scholar] [CrossRef]
- Zhong, Y.J.; Wang, T.; Luo, R.L.; Liu, J.Y.; Jin, R.Y.; Peng, X.L. Recent advances and potentiality of postbiotics in the food industry: Composition, inactivation methods, current applications in metabolic syndrome, and future trends. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Szajewska, H.; Ruszczynski, M.; Kolacek, S. Meta-analysis shows limited evidence for using Lactobacillus acidophilus LB to treat acute gastroenteritis in children. Acta Paediatr. 2014, 103, 249–255. [Google Scholar] [CrossRef]
- Andresen, V.; Gschossmann, J.; Layer, P. Heat-inactivated Bifidobacterium bifidum MIMBb75 (SYN-HI-001) in the treatment of irritable bowel syndrome: A multicentre, randomised, double-blind, placebo-controlled clinical trial. Lancet Gastroenterol. Hepatol. 2020, 5, 658–666. [Google Scholar] [CrossRef]
- Martyniak, A.; Medynska-Przeczek, A.; Wedrychowicz, A.; Skoczen, S.; Tomasik, P.J. Prebiotics, probiotics, synbiotics, paraprobiotics and postbiotic compounds in IBD. Biomolecules 2021, 11, 1903. [Google Scholar] [CrossRef]
- Feng, C.J.; Zhang, W.Q.; Zhang, T.; He, Q.W.; Kwok, L.Y.; Tan, Y.; Zhang, H.P. Heat-killed Bifidobacterium bifidum B1628 may alleviate dextran sulfate sodium-induced colitis in mice, and the anti-inflammatory effect is associated with gut microbiota modulation. Nutrients 2022, 14, 5233. [Google Scholar] [CrossRef]
- Shida, K.; Kiyoshima-Shibata, J.; Kaji, R.; Nagaoka, M.; Nanno, M. Peptidoglycan from lactobacilli inhibits interleukin-12 production by macrophages induced by Lactobacillus casei through Toll-like receptor 2-dependent and independent mechanisms. Immunology 2009, 128, e858–e869. [Google Scholar] [CrossRef]
- Kim, K.W.; Kang, S.S.; Woo, S.J.; Park, O.J.; Ahn, K.B.; Song, K.D.; Lee, H.K.; Yun, C.H.; Han, S.H. Lipoteichoic acid of probiotic Lactobacillus plantarum attenuates Poly I:C-induced IL-8 production in porcine intestinal epithelial cells. Front. Microbiol. 2017, 8, 1827. [Google Scholar] [CrossRef] [PubMed]
- El-Deeb, N.M.; Yassin, A.M.; Al-Madboly, L.A.; El-Hawiet, A. A novel purified Lactobacillus acidophilus 20079 exopolysaccharide, LA-EPS-20079, molecularly regulates both apoptotic and NF-κB inflammatory pathways in human colon cancer. Microb. Cell Factories 2018, 17, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irrazabal, T.; Thakur, B.K.; Croitoru, K.; Martin, A. Preventing colitis-associated colon cancer with antioxidants: A systematic review. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1177–1197. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.Z.; et al. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.Y.; Liu, X.J.; Hao, J.Y. Gut microbiota in ulcerative colitis: Insights on pathogenesis and treatment. J. Dig. Dis. 2020, 21, 147–159. [Google Scholar] [CrossRef]
- Kushkevych, I.; Dordevic, D.; Vitezova, M. Possible synergy effect of hydrogen sulfide and acetate produced by sulfate-reducing bacteria on inflammatory bowel disease development. J. Adv. Res. 2021, 27, 71–78. [Google Scholar] [CrossRef]
- Jiang, L.; Yuan, C.C.; Ye, W.X.; Huang, Q.X.; Chen, Z.; Wu, W.Z.; Qian, L.C. Akkermansia and its metabolites play key roles in the treatment of campylobacteriosis in mice. Front. Immunol. 2023, 13, 1061627. [Google Scholar] [CrossRef]
- Hadinia, N.; Dovom, M.R.E.; Yavarmanesh, M. The effect of fermentation conditions (temperature, salt concentration, and pH) with lactobacillus strains for producing short chain fatty acids. Lwt-Food Sci. Technol. 2022, 165, 113709. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, N.; Corr, S.C. Gut microbial metabolite-mediated regulation of the intestinal barrier in the pathogenesis of inflammatory bowel disease. Nutrients 2021, 13, 4259. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Y.; Wang, T.; Wang, X.; Lü, X. The Protective Effect of Heat-Inactivated Companilactobacillus crustorum on Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Nutrients 2023, 15, 2746. https://doi.org/10.3390/nu15122746
Zhong Y, Wang T, Wang X, Lü X. The Protective Effect of Heat-Inactivated Companilactobacillus crustorum on Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Nutrients. 2023; 15(12):2746. https://doi.org/10.3390/nu15122746
Chicago/Turabian StyleZhong, Yujie, Tao Wang, Xin Wang, and Xin Lü. 2023. "The Protective Effect of Heat-Inactivated Companilactobacillus crustorum on Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice" Nutrients 15, no. 12: 2746. https://doi.org/10.3390/nu15122746