

Rutin Promotes Proliferation and Orchestrates Epithelial–Mesenchymal Transition and Angiogenesis in MCF-7 and MDA-MB-231 Breast Cancer Cells

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
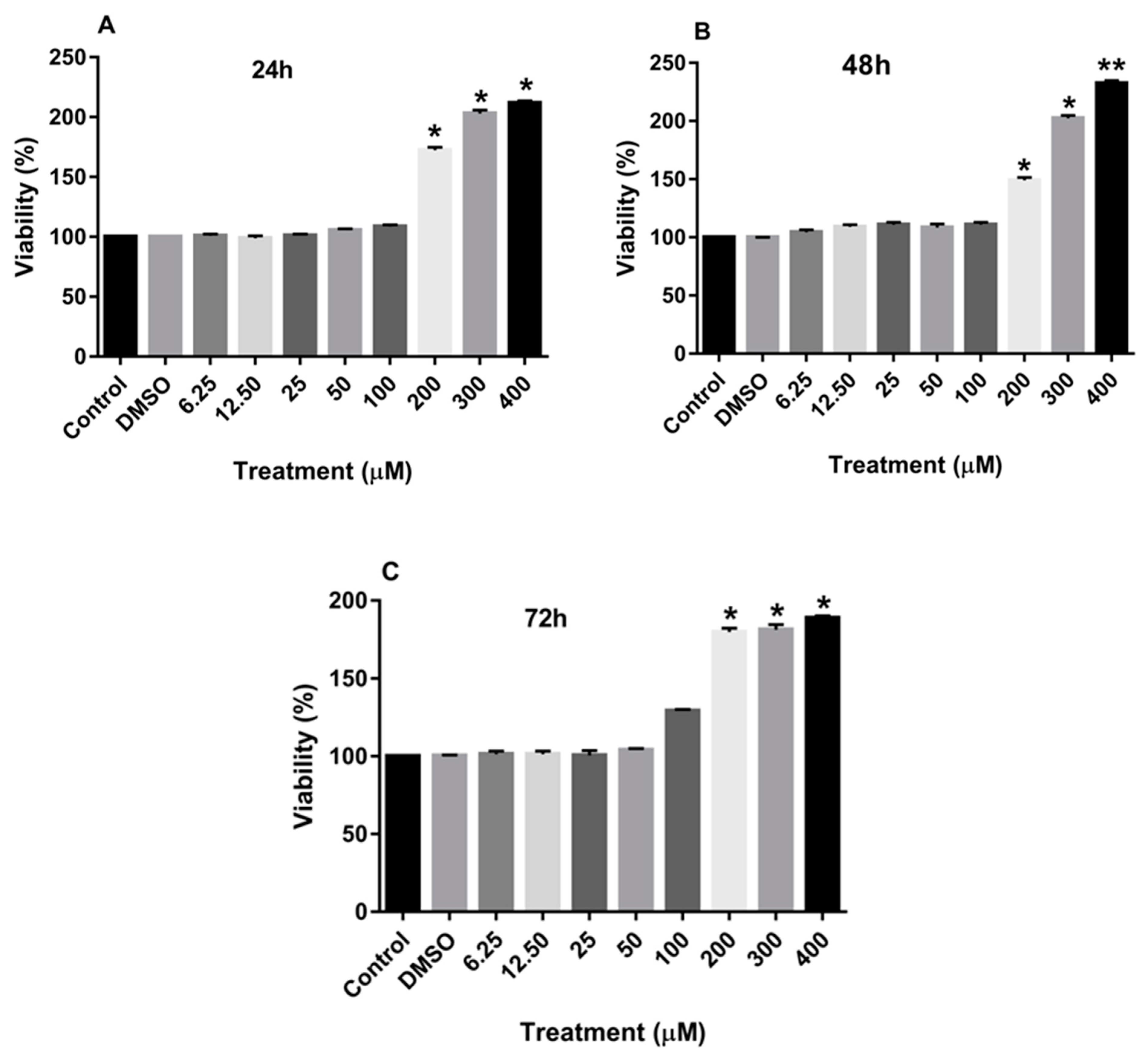
2.2. Cell Viability
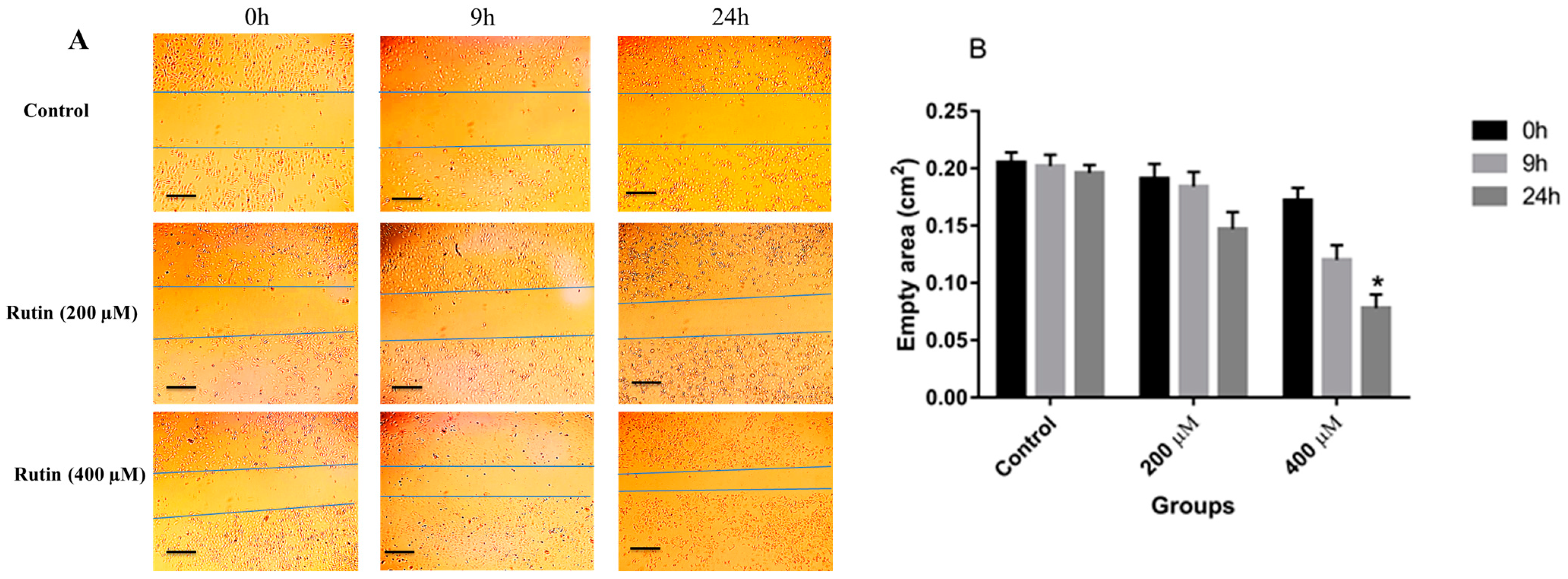
2.3. Wound Healing Assay
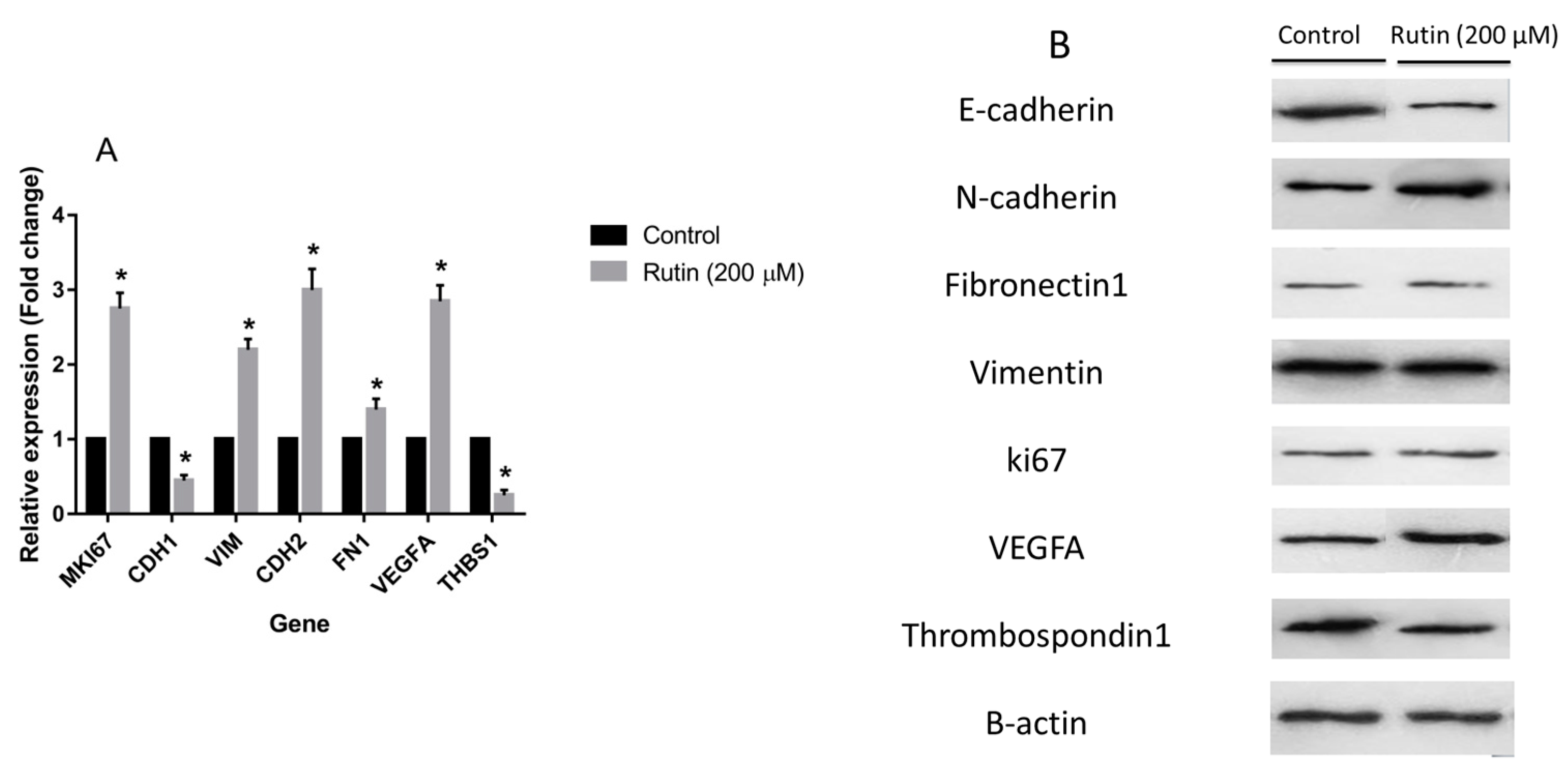
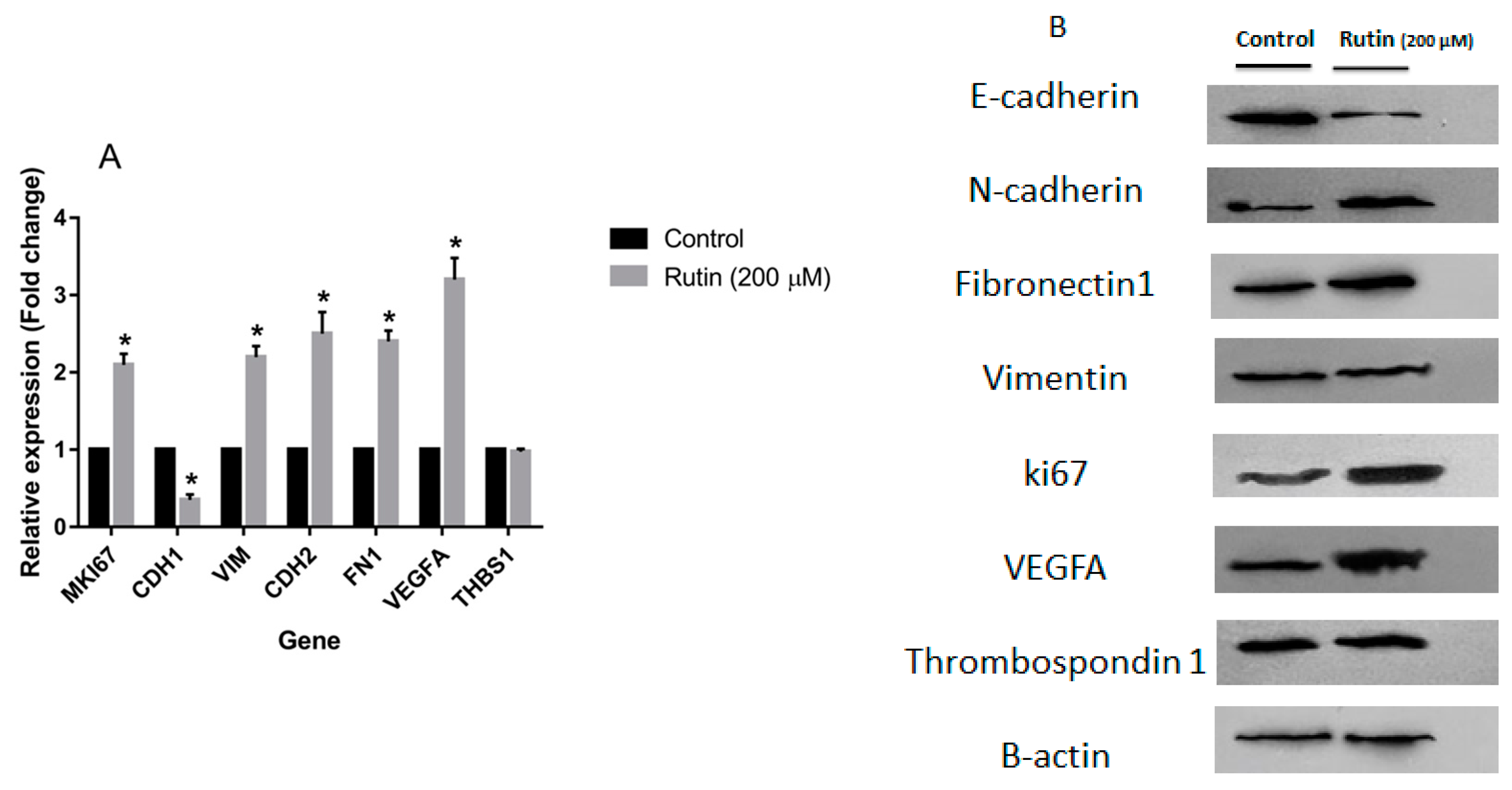
2.4. Real-Time PCR Analysis
2.5. Western Blot Analysis
2.6. Data Analysis
3. Results
3.1. Effect of Rutin on the Proliferation of MDA-MB-231 and MCF-7 Cell Lines
3.2. Effects of Rutin on the Invasion and Migration of MDA-MB-231 and MCF-7 Cell Lines
3.3. Effects of Rutin on Gene and Protein Expression in MDA-MB-231 and MCF-7 Cell Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trayes, K.P.; Cokenakes, S.E.H. Breast Cancer Treatment. Am. Fam. Physician 2021, 104, 171–178. [Google Scholar] [PubMed]
- Azamjah, N.; Soltan-Zadeh, Y.; Zayeri, F. Global Trend of Breast Cancer Mortality Rate: A 25-Year Study. Asian Pac. J. Cancer Prev. 2019, 20, 2015–2020. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, A.M.; Friebel-Klingner, T.; Ehsan, S.; He, W.; Welch, M.; Chen, J.; Kontos, D.; Domchek, S.M.; Conant, E.F.; Semine, A.; et al. Relationship of established risk factors with breast cancer subtypes. Cancer Med. 2021, 10, 6456–6467. [Google Scholar] [CrossRef] [PubMed]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Shahali, A.; Ghanadian, M.; Jafari, S.M.; Aghaei, M. Mitochondrial and caspase pathways are involved in the induction of apoptosis by nardosinen in MCF-7 breast cancer cell line. Res. Pharm. Sci. 2018, 13, 12–21. [Google Scholar]
- Malik, J.A.; Ahmed, S.; Jan, B.; Bender, O.; Al Hagbani, T.; Alqarni, A.; Anwar, S. Drugs repurposed: An advanced step towards the treatment of breast cancer and associated challenges. Biomed. Pharmacother. 2022, 145, 112375. [Google Scholar] [CrossRef]
- Rajabi, S.; Rajani, H.F.; Mohammadkhani, N.; Ramírez-Coronel, A.A.; Maleki, M.; Maresca, M.; Hajimehdipoor, H. Long Non-Coding RNAs as Novel Targets for Phytochemicals to Cease Cancer Metastasis. Molecules 2023, 28, 987. [Google Scholar] [CrossRef]
- Park, M.; Kim, D.; Ko, S.; Kim, A.; Mo, K.; Yoon, H. Breast Cancer Metastasis: Mechanisms and Therapeutic Implications. Int. J. Mol. Sci. 2022, 23, 6806. [Google Scholar] [CrossRef]
- Karamanou, K.; Franchi, M.; Vynios, D.; Brézillon, S. Epithelial-to-mesenchymal transition and invadopodia markers in breast cancer: Lumican a key regulator. Semin. Cancer Biol. 2020, 62, 125–133. [Google Scholar] [CrossRef]
- Shakib, H.; Rajabi, S.; Dehghan, M.H.; Mashayekhi, F.J.; Safari-Alighiarloo, N.; Hedayati, M. Epithelial-to-mesenchymal transition in thyroid cancer: A comprehensive review. Endocrine 2019, 66, 435–455. [Google Scholar] [CrossRef]
- Kim, D.H.; Xing, T.; Yang, Z.; Dudek, R.; Lu, Q.; Chen, Y.H. Epithelial Mesenchymal Transition in Embryonic Development, Tissue Repair and Cancer: A Comprehensive Overview. J. Clin. Med. 2017, 7, 1. [Google Scholar] [CrossRef]
- Malla, R.; Puvalachetty, K.; Vempati, R.K.; Marni, R.; Merchant, N.; Nagaraju, G.P. Cancer Stem Cells and Circulatory Tumor Cells Promote Breast Cancer Metastasis. Clin. Breast Cancer 2022, 22, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, S.; Dehghan, M.H.; Dastmalchi, R.; Jalali Mashayekhi, F.; Salami, S.; Hedayati, M. The roles and role-players in thyroid cancer angiogenesis. Endocr. J. 2019, 66, 277–293. [Google Scholar] [CrossRef]
- Caparica, R.; Júlio, A.; Araújo, M.E.M.; Baby, A.R.; Fonte, P.; Costa, J.G.; Santos de Almeida, T. Anticancer Activity of Rutin and Its Combination with Ionic Liquids on Renal Cells. Biomolecules 2020, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Ben Sghaier, M.; Pagano, A.; Mousslim, M.; Ammari, Y.; Kovacic, H.; Luis, J. Rutin inhibits proliferation, attenuates superoxide production and decreases adhesion and migration of human cancerous cells. Biomed. Pharmacother. 2016, 84, 1972–1978. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Kubina, R.; Cochis, A.; Sorrentino, R.; Varoni, E.M.; Kabała-Dzik, A.; Azzimonti, B.; Dziedzic, A.; Rimondini, L.; Wojtyczka, R.D. Rutin, a Quercetin Glycoside, Restores Chemosensitivity in Human Breast Cancer Cells. Phytother. Res. 2017, 31, 1529–1538. [Google Scholar] [CrossRef]
- Elsayed, H.E.; Ebrahim, H.Y.; Mohyeldin, M.M.; Siddique, A.B.; Kamal, A.M.; Haggag, E.G.; El Sayed, K.A. Rutin as A Novel c-Met Inhibitory Lead for The Control of Triple Negative Breast Malignancies. Nutr. Cancer 2017, 69, 1256–1271. [Google Scholar] [CrossRef]
- Hedayati, M.; Rajabi, S.; Nikzamir, A. Papillary Thyroid Cancer-Promoting Activities of Combined Oral Contraceptive Components. Galen Med. J. 2020, 9, e1648. [Google Scholar] [CrossRef] [PubMed]
- Kielkopf, C.L.; Bauer, W.; Urbatsch, I.L. Bradford Assay for Determining Protein Concentration. Cold Spring Harb. Protoc. 2020, 2020, 102269. [Google Scholar] [CrossRef]
- Feng, S.; Yin, J. Radiomics of dynamic contrast-enhanced magnetic resonance imaging parametric maps and apparent diffusion coefficient maps to predict Ki-67 status in breast cancer. Front. Oncol. 2022, 12, 847880. [Google Scholar] [CrossRef]
- Lai, X.; Li, Q.; Wu, F.; Lin, J.; Chen, J.; Zheng, H.; Guo, L. Epithelial-Mesenchymal Transition and Metabolic Switching in Cancer: Lessons from Somatic Cell Reprogramming. Front. Cell Dev. Biol. 2020, 8, 760. [Google Scholar] [CrossRef] [PubMed]
- Loh, C.Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [PubMed]
- Dudley, A.C.; Griffioen, A.W. Pathological angiogenesis: Mechanisms and therapeutic strategies. Angiogenesis 2023, 1, 1–35. [Google Scholar] [CrossRef]
- Canivenc-Lavier, M.C.; Bennetau-Pelissero, C. Phytoestrogens and Health Effects. Nutrients 2023, 15, 317. [Google Scholar] [CrossRef] [PubMed]
- Van Duursen, M.B.M. Modulation of estrogen synthesis and metabolism by phytoestrogens in vitro and the implications for women’s health. Toxicol. Res. 2017, 6, 772–794. [Google Scholar] [CrossRef]
- Guo, X.-D.; Diao, Q.-Y.; Wang, Y.-Y.; Tu, Y.; Deng, K.-D.; Wang, X.-J.; Fu, T.; Yan, G.-L. The Effect of Administration of Rutin on Plasma Levels of Estrogen, Prolactin, Growth Hormone and Gene Expression of Their Receptors in Mammary Glands in Ovariectomized Rats. J. Integr. Agric. 2012, 11, 1700–1706. [Google Scholar] [CrossRef]
- Harris, R.M.; Wood, D.M.; Bottomley, L.; Blagg, S.; Owen, K.; Hughes, P.J.; Waring, R.H.; Kirk, C.J. Phytoestrogens are potent inhibitors of estrogen sulfation: Implications for breast cancer risk and treatment. J. Clin. Endocrinol. Metab. 2004, 89, 1779–1787. [Google Scholar] [CrossRef]
- Kirk, C.J.; Harris, R.M.; Wood, D.M.; Waring, R.H.; Hughes, P.J. Do dietary phytoestrogens influence susceptibility to hormone-dependent cancer by disrupting the metabolism of endogenous oestrogens? Biochem. Soc. Trans. 2001, 29, 209–216. [Google Scholar] [CrossRef]
- Yuan, B.; Wang, L.; Jin, Y.; Zhen, H.; Xu, P.; Xu, Y.; Li, C.; Xu, H. Role of metabolism in the effects of genistein and its phase II conjugates on the growth of human breast cell lines. AAPS J. 2012, 14, 329–344. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| MKI67 | 5′-GCTACTCCAAAGAAGCCTGTG-3′ | 5′-AAGTTGTTGAGCACTCTGTAGG-3′ |
| CDH1 | 5′-GGGGTCTGTCATGGAAGGTG-3′ | 5′-CGACGTTAGCCTCGTTCTCA-3′ |
| CDH2 | 5′-GCGTCTGTAGAGGCTTCTGG-3′ | 5′-GCCACTTGCCACTTTTCCTG-3′ |
| FN1 | 5′-ACAAGCATGTCTCTCTGCCAA-3′ | 5′-TCAGGAAACTCCCAGGGTGA-3′ |
| VIM | 5′-TCCGCACATTCGAGCAAAGA-3′ | 5′-ATTCAAGTCTCAGCGGGCTC-3′ |
| VEGFA | 5′-GAGCAAGACAAGAAAATCCC-3′ | 5′-CCTCGGCTTGTCACATCTG-3′ |
| THBS1 | 5′-CCCTTGTGCTCAGAGTGGAT-3′ | 5′-GCCAGTAGAGAACAAATAAGCATGG-3′ |
| GAPDH | 5′-ACCCACTCCTCCACCTTTGA-3′ | 5′-CT GTTGCTGTAGCCAAATTCGT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajimehdipoor, H.; Tahmasvand, Z.; Nejad, F.G.; Maresca, M.; Rajabi, S. Rutin Promotes Proliferation and Orchestrates Epithelial–Mesenchymal Transition and Angiogenesis in MCF-7 and MDA-MB-231 Breast Cancer Cells. Nutrients 2023, 15, 2884. https://doi.org/10.3390/nu15132884
Hajimehdipoor H, Tahmasvand Z, Nejad FG, Maresca M, Rajabi S. Rutin Promotes Proliferation and Orchestrates Epithelial–Mesenchymal Transition and Angiogenesis in MCF-7 and MDA-MB-231 Breast Cancer Cells. Nutrients. 2023; 15(13):2884. https://doi.org/10.3390/nu15132884
Chicago/Turabian StyleHajimehdipoor, Homa, Zahra Tahmasvand, Fatemeh Ghorban Nejad, Marc Maresca, and Sadegh Rajabi. 2023. "Rutin Promotes Proliferation and Orchestrates Epithelial–Mesenchymal Transition and Angiogenesis in MCF-7 and MDA-MB-231 Breast Cancer Cells" Nutrients 15, no. 13: 2884. https://doi.org/10.3390/nu15132884
APA StyleHajimehdipoor, H., Tahmasvand, Z., Nejad, F. G., Maresca, M., & Rajabi, S. (2023). Rutin Promotes Proliferation and Orchestrates Epithelial–Mesenchymal Transition and Angiogenesis in MCF-7 and MDA-MB-231 Breast Cancer Cells. Nutrients, 15(13), 2884. https://doi.org/10.3390/nu15132884