Effects of FODMAPs and Gluten on Gut Microbiota and Their Association with the Metabolome in Irritable Bowel Syndrome: A Double-Blind, Randomized, Cross-Over Intervention Study

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Study Design
2.2. Dietary Interventions
2.3. Questionnaires
2.4. Collection of Fecal Samples
2.5. Gut Microbiota Data
2.6. Analysis of Short-Chain Fatty Acids
2.6.1. Sample Preparation of Plasma Samples
2.6.2. Sample Preparation of Fecal Samples
2.6.3. Parameters of LC-MS
2.7. Metabolite Data
2.8. Data Analysis
3. Results
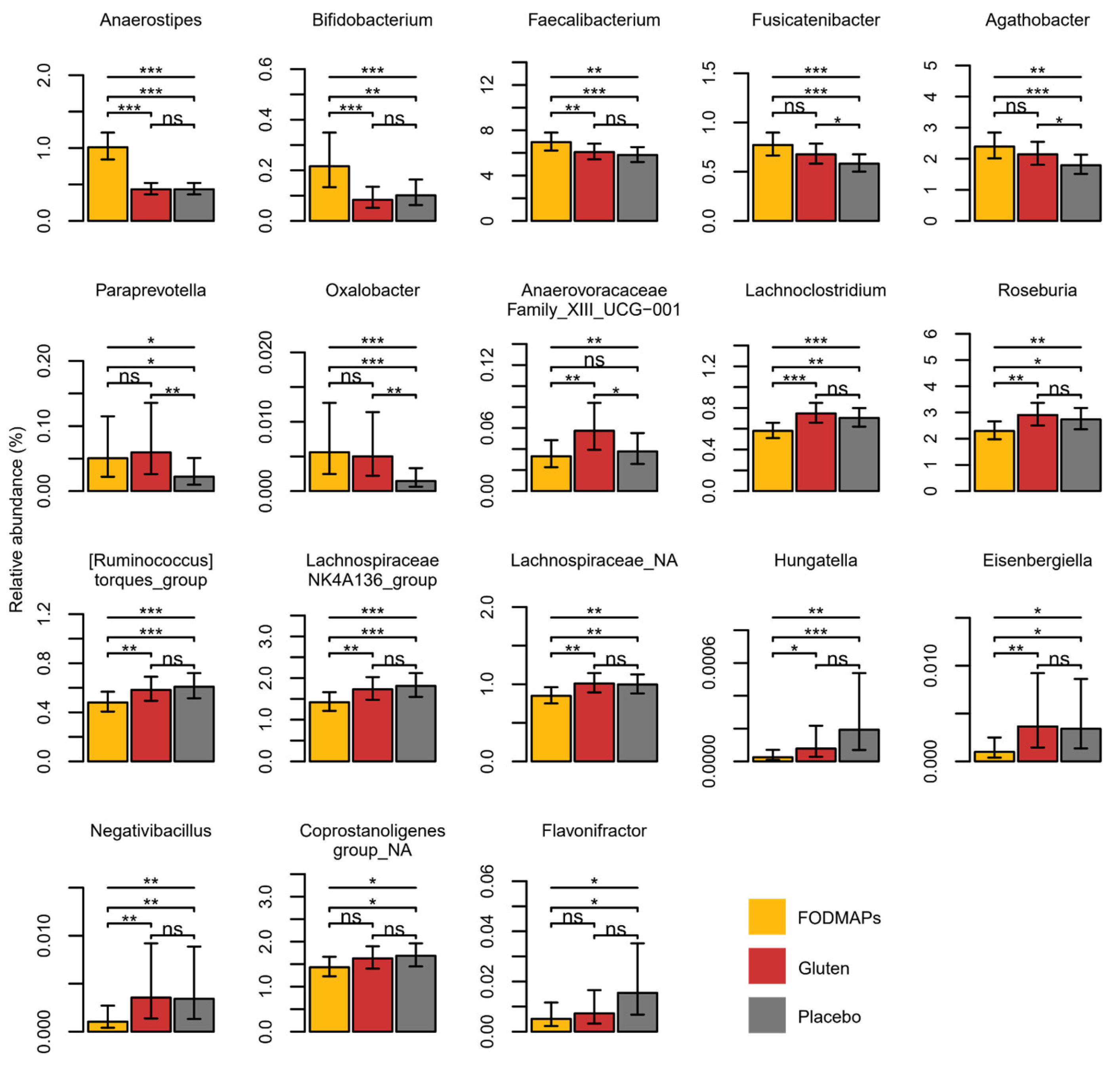
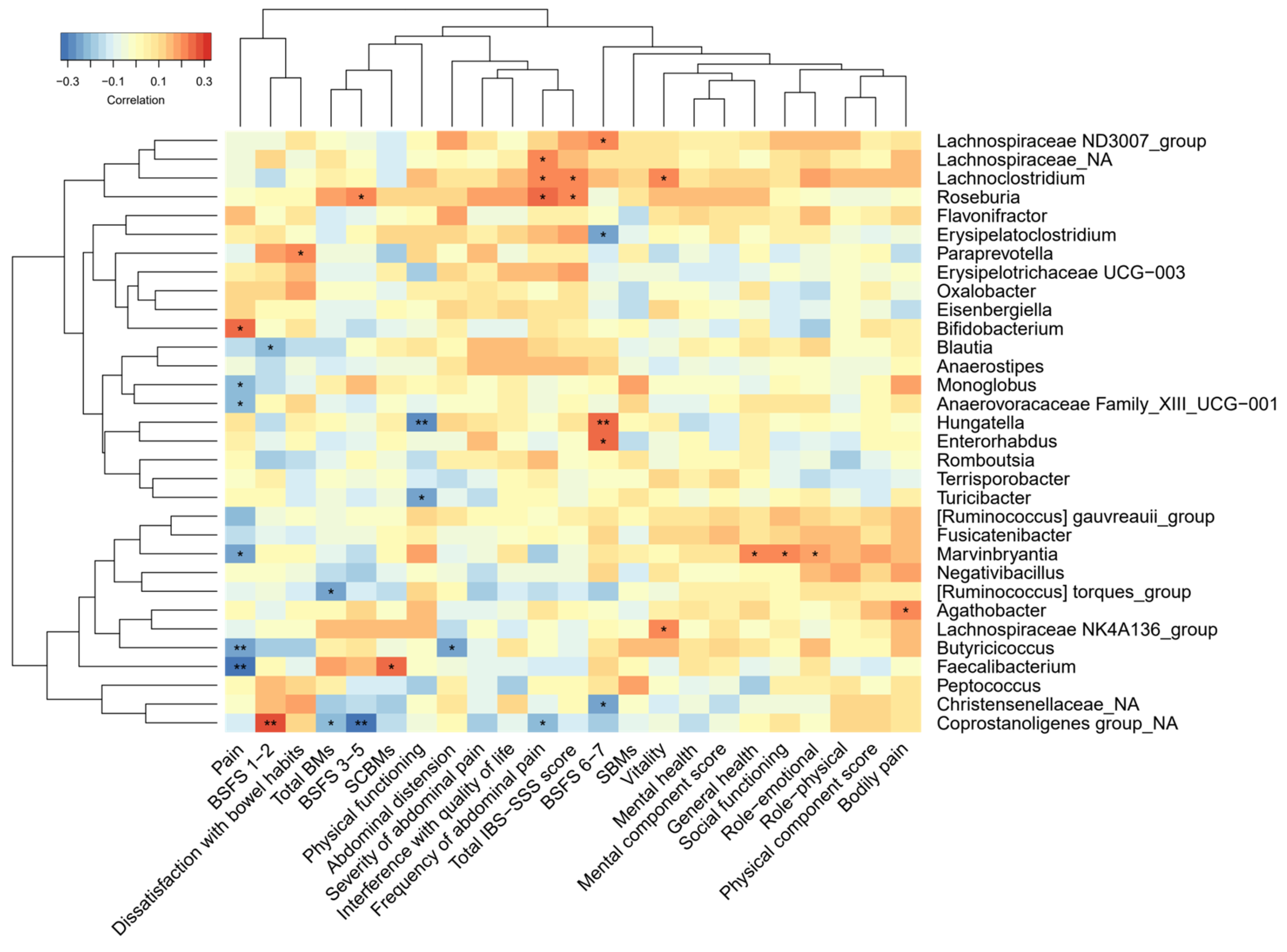
3.1. Gut Microbiota, SCFAs and Correlation with IBS Symptoms
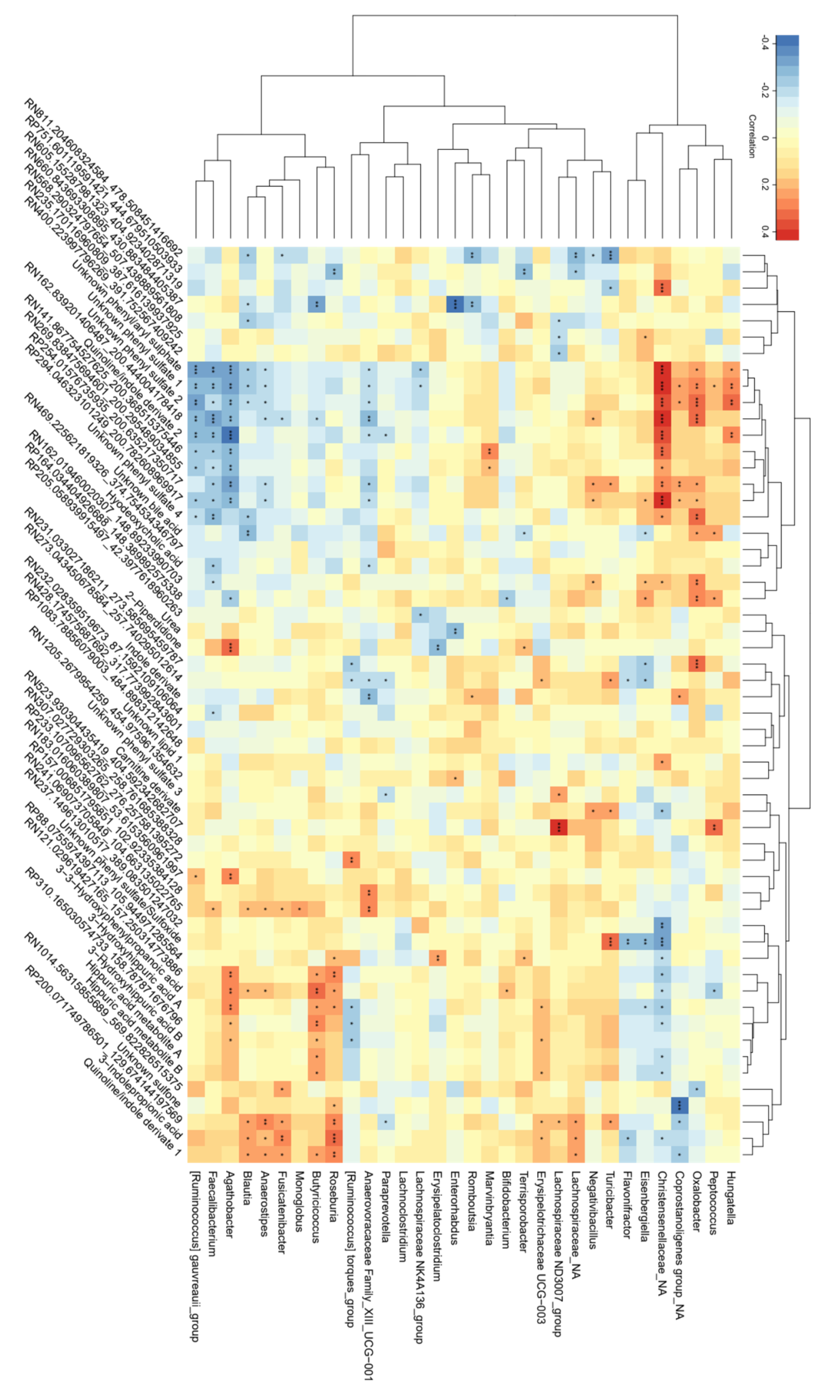
3.2. Correlations between Bacterial Genera and Metabolites Related to FODMAP Exposure
4. Discussion
4.1. Intervention Effects on the Gut Microbiota Composition and SCFAs
4.2. Correlations between Gut Microbiota and IBS Symptoms
4.3. Correlations between Bacterial Genera and Metabolites Related to Intake of FODMAPs
4.4. Gluten
4.5. Limitation and Strength
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drossman, D.A. Functional Gastrointestinal Disorders: History, Pathophysiology, Clinical Features and Rome IV. Gastroenterology 2016, 150, 1262–1279.e2. [Google Scholar] [CrossRef]
- El-Salhy, M.; Mazzawi, T. Fecal Microbiota Transplantation for Managing Irritable Bowel Syndrome. Expert Rev. Gastroenterol. Hepatol. 2018, 12, 439–445. [Google Scholar] [CrossRef]
- El-Salhy, M.; Mazzawi, T.; Gundersen, D.; Hausken, T. Chromogranin A Cell Density in the Rectum of Patients with Irritable Bowel Syndrome. Mol. Med. Rep. 2012, 6, 1223–1225. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Stress and the Gut: Pathophysiology, Clinical Consequences, Diagnostic Approach and Treatment Options. J. Physiol. Pharmacol. 2011, 62, 591–599. [Google Scholar]
- El-Salhy, M.; Seim, I.; Chopin, L.; Gundersen, D.; Hatlebakk, J.G.; Hausken, T. Irritable Bowel Syndrome: The Role of Gut Neuroendocrine Peptides. Front. Biosci.-Elite 2012, 4, 2683–2700. [Google Scholar] [CrossRef] [PubMed]
- Dionne, J.; Ford, A.C.; Yuan, Y.; Chey, W.D.; Lacy, B.E.; Saito, Y.A.; Quigley, E.M.M.; Moayyedi, P. A Systematic Review and Meta-Analysis Evaluating the Efficacy of a Gluten-Free Diet and a Low FODMAPS Diet in Treating Symptoms of Irritable Bowel Syndrome. Am. J. Gastroenterol. 2018, 113, 1290–1300. [Google Scholar] [CrossRef]
- Staudacher, H.M.; Whelan, K. The Low FODMAP Diet: Recent Advances in Understanding Its Mechanisms and Efficacy in IBS. Gut 2017, 66, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, H.W.J.; Naylor, C.P.E.; MacFarlane, G.T. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Gibson, G.R.; Cummings, J.H. Comparison of Fermentation Reactions in Different Regions of the Human Colon. J. Appl. Bacteriol. 1992, 72, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Farias, S. Effect of Inulin on the Human Gut Microbiota: Stimulation of Bifidobacterium Adolescentis and Faecalibacterium Prausnitzii. Br. J. Nutr. 2008, 101, 541–550. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Reed, D.E.; Schneider, T.; Dang, F.; Keshteli, A.H.; de Palma, G.; Madsen, K.; Bercik, P.; Vanner, S. FODMAPs Alter Symptoms and the Metabolome of Patients with IBS: A Randomised Controlled Trial. Gut 2017, 66, 1241–1251. [Google Scholar] [CrossRef]
- Hustoft, T.N.; Hausken, T.; Ystad, S.O.; Valeur, J.; Brokstad, K.; Hatlebakk, J.G.; Lied, G.A. Effects of Varying Dietary Content of Fermentable Short-Chain Carbohydrates on Symptoms, Fecal Microenvironment, and Cytokine Profiles in Patients with Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2017, 29, e12969. [Google Scholar] [CrossRef] [PubMed]
- Sloan, T.J.; Jalanka, J.; Major, G.A.D.; Krishnasamy, S.; Pritchard, S.; Abdelrazig, S.; Korpela, K.; Singh, G.; Mulvenna, C.; Hoad, C.L.; et al. A Low FODMAP Diet Is Associated with Changes in the Microbiota and Reduction in Breath Hydrogen but Not Colonic Volume in Healthy Subjects. PLoS ONE 2018, 13, e0201410. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, H.M.; Lomer, M.C.E.; Anderson, J.L.; Barrett, J.S.; Muir, J.G.; Irving, P.M.; Whelan, K. Fermentable Carbohydrate Restriction Reduces Luminal Bifidobacteria and Gastrointestinal Symptoms in Patients with Irritable Bowel Syndrome. J. Nutr. 2012, 142, 1510–1518. [Google Scholar] [CrossRef]
- So, D.; Loughman, A.; Staudacher, H.M. Effects of a Low FODMAP Diet on the Colonic Microbiome in Irritable Bowel Syndrome: A Systematic Review with Meta-Analysis. Am. J. Clin. Nutr. 2022, 116, 943–952. [Google Scholar] [CrossRef]
- Gibson, P.R.; Halmos, E.P.; Muir, J.G. Review Article: FODMAPS, Prebiotics and Gut Health-the FODMAP Hypothesis Revisited. Aliment. Pharmacol. Ther. 2020, 52, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Halmos, E.P.; Christophersen, C.T.; Bird, A.R.; Shepherd, S.J.; Gibson, P.R.; Muir, J.G. Diets That Differ in Their FODMAP Content Alter the Colonic Luminal Microenvironment. Gut 2015, 64, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Valeur, J.; Røseth, A.G.; Knudsen, T.; Malmstrøm, G.H.; Fiennes, J.T.; Midtvedt, T.; Berstad, A. Fecal Fermentation in Irritable Bowel Syndrome: Influence of Dietary Restriction of Fermentable Oligosaccharides, Disaccharides, Monosaccharides and Polyols. Digestion 2016, 94, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Biesiekierski, J.R.; Newnham, E.D.; Irving, P.M.; Barrett, J.S.; Haines, M.; Doecke, J.D.; Shepherd, S.J.; Muir, J.G.; Gibson, P.R. Gluten Causes Gastrointestinal Symptoms in Subjects Without Celiac Disease: A Double-Blind Randomized Placebo-Controlled Trial. Am. J. Gastroenterol. 2011, 106, 508–514. [Google Scholar] [CrossRef]
- Skodje, G.I.; Sarna, V.K.; Minelle, I.H.; Rolfsen, K.L.; Muir, J.G.; Gibson, P.R.; Veierød, M.B.; Henriksen, C.; Lundin, K.E.A. Fructan, Rather Than Gluten, Induces Symptoms in Patients With Self-Reported Non-Celiac Gluten Sensitivity. Gastroenterology 2018, 154, 529–539.e2. [Google Scholar] [CrossRef]
- Mohseni, F.; Agah, S.; Ebrahimi-Daryani, N.; Taher, M.; Nattagh-Eshtivani, E.; Karimi, S.; Rastgoo, S.; Bourbour, F.; Hekmatdoost, A. The Effect of Low FODMAP Diet with and without Gluten on Irritable Bowel Syndrome: A Double Blind, Placebo Controlled Randomized Clinical Trial. Clin. Nutr. ESPEN 2022, 47, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Nordin, E.; Brunius, C.; Landberg, R.; Hellström, P.M. Fermentable Oligo-, Di-, Monosaccharides, and Polyols (FODMAPs), but Not Gluten, Elicit Modest Symptoms of Irritable Bowel Syndrome: A Double-Blind, Placebo-Controlled, Randomized Three-Way Crossover Trial. Am. J. Clin. Nutr. 2022, 115, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Biesiekierski, J.R.; Iven, J. Non-Coeliac Gluten Sensitivity: Piecing the Puzzle Together. United Eur. Gastroenterol. J. 2015, 3, 160–165. [Google Scholar] [CrossRef]
- Dieterich, W.; Schuppan, D.; Schink, M.; Schwappacher, R.; Wirtz, S.; Agaimy, A.; Neurath, M.F.; Zopf, Y. Influence of Low FODMAP and Gluten-Free Diets on Disease Activity and Intestinal Microbiota in Patients with Non-Celiac Gluten Sensitivity. Clin. Nutr. 2019, 38, 697–707. [Google Scholar] [CrossRef]
- Golfetto, L.; de Senna, F.D.; Hermes, J.; Beserra, B.T.S.; França, F.; França, F.d.S.; Martinello, F. Lower Bifidobacteria Counts in Adult Patients with Celiac Disease on a Gluten-Free Diet. Arq. Gastroenterol. 2014, 51, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.B.S.; Roager, H.M.; Søndertoft, N.B.; Gøbel, R.J.; Kristensen, M.; Vallès-Colomer, M.; Vieira-Silva, S.; Ibrügger, S.; Lind, M.V.; Mærkedahl, R.B.; et al. A Low-Gluten Diet Induces Changes in the Intestinal Microbiome of Healthy Danish Adults. Nat. Commun. 2018, 9, 4630. [Google Scholar] [CrossRef]
- Bonder, M.J.; Tigchelaar, E.F.; Cai, X.; Trynka, G.; Cenit, M.C.; Hrdlickova, B.; Zhong, H.; Vatanen, T.; Gevers, D.; Wijmenga, C.; et al. The Influence of a Short-Term Gluten-Free Diet on the Human Gut Microbiome. Genome Med. 2016, 8, 45. [Google Scholar] [CrossRef]
- Caminero, A.; Nistal, E.; Arias, L.; Vivas, S.; Comino, I.; Real, A.; Sousa, C.; de Morales, J.M.R.; Ferrero, M.A.; Rodríguez-Aparicio, L.B.; et al. A Gluten Metabolism Study in Healthy Individuals Shows the Presence of Faecal Glutenasic Activity. Eur. J. Nutr. 2012, 51, 293–299. [Google Scholar] [CrossRef]
- Subramanian, I.; Verma, S.; Kumar, S.; Jere, A.; Anamika, K. Multi-Omics Data Integration, Interpretation, and Its Application. Bioinform. Biol. Insights 2020, 14, 1177932219899051. [Google Scholar] [CrossRef]
- Lamichhane, S.; Sen, P.; Dickens, A.M.; Orešič, M.; Bertram, H.C. Gut Metabolome Meets Microbiome: A Methodological Perspective to Understand the Relationship between Host and Microbe. Methods 2018, 149, 3–12. [Google Scholar] [CrossRef]
- Mayneris-Perxachs, J.; Fernández-Real, J.M. Exploration of the Microbiota and Metabolites within Body Fluids Could Pinpoint Novel Disease Mechanisms. FEBS J. 2020, 287, 856–865. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Tyson, G.W. Metagenomics. Nature 2008, 455, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut Microbial Metabolites as Multi-Kingdom Intermediates. Nat. Rev. Microbiol. 2020, 19, 77–94. [Google Scholar] [CrossRef]
- Fobofou, S.A.; Savidge, T. Microbial Metabolites: Cause or Consequence in Gastrointestinal Disease? Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 322, G535–G552. [Google Scholar] [CrossRef]
- Zierer, J.; Jackson, M.A.; Kastenmüller, G.; Mangino, M.; Long, T.; Telenti, A.; Mohney, R.P.; Small, K.S.; Bell, J.T.; Steves, C.J.; et al. The Fecal Metabolome as a Functional Readout of the Gut Microbiome Europe PMC Funders Group. Nat. Genet. 2018, 50, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, K.F.; Sayols-Baixeras, S.; Baldanzi, G.; Nowak, C.; Hammar, U.; Nguyen, D.; Varotsis, G.; Brunkwall, L.; Nielsen, N.; Eklund, A.C.; et al. An Online Atlas of Human Plasma Metabolite Signatures of Gut Microbiome Composition. Nat. Commun. 2022, 13, 5370. [Google Scholar] [CrossRef]
- Bosi, A.; Banfi, D.; Bistoletti, M.; Giaroni, C.; Baj, A. Tryptophan Metabolites Along the Microbiota-Gut-Brain Axis: An Interkingdom Communication System Influencing the Gut in Health and Disease. Int. J. Tryptophan Res. 2020, 13, 1178646920928984. [Google Scholar] [CrossRef] [PubMed]
- Nordin, E.; Hellström, P.M.; Vuong, E.; Ribbenstedt, A.; Brunius, C.; Landberg, R. IBS Randomized Crossover Challenge Study: FODMAPs Alter Bile Acids, Tryptophan and Phenolic-Derived Metabolites, While Gluten Modifies Lipid. 2023. Available online: https://journals.physiology.org/doi/abs/10.1152/ajpregu.00016.2023 (accessed on 1 May 2023).
- Tuomainen, M.; Lindström, J.; Lehtonen, M.; Auriola, S.; Pihlajamäki, J.; Peltonen, M.; Tuomilehto, J.; Uusitupa, M.; de Mello, V.D.; Hanhineva, K. Associations of Serum Indolepropionic Acid, a Gut Microbiota Metabolite, with Type 2 Diabetes and Low-Grade Inflammation in High-Risk Individuals. Nutr. Diabetes 2018, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Konopelski, P.; Mogilnicka, I. Biological Effects of Indole-3-Propionic Acid, a Gut Microbiota-Derived Metabolite, and Its Precursor Tryptophan in Mammals’ Health and Disease. Int. J. Mol. Sci. 2022, 23, 1222. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Mu, C.L.; Farzi, A.; Zhu, W.Y. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial Tryptophan Catabolites in Health and Disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Robinson, H.; Cai, T.; Tagle, D.A.; Li, J. Structural Insight into the Inhibition of Human Kynurenine Aminotransferase I/Glutamine Transaminase K. J. Med. Chem. 2009, 52, 2786–2793. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, P.; Cassidy Eugene, M.; Clarke, G.; Scully, P.; Barry, S.; Quigley Eamonn, M.M.; Shanahan, F.; Cryan, J.; Dinan Timothy, G. Tryptophan Catabolism in Females with Irritable Bowel Syndrome: Relationship to Interferon-Gamma, Severity of Symptoms and Psychiatric Co-Morbidity. Neurogastroenterol. Motil. 2008, 20, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Hill, P.; Muir, J.G.; Gibson, P.R. Controversies and Recent Developments of the Low-FODMAP Diet. Gastroenterol. Hepatol. 2017, 13, 36–45. [Google Scholar]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia Spp.: A Marker of Health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef]
- Halmos, E.P.; Power, V.A.; Shepherd, S.J.; Gibson, P.R.; Muir, J.G. A Diet Low in FODMAPs Reduces Symptoms of Irritable Bowel Syndrome. Gastroenterology 2014, 146, 67–75.e5. [Google Scholar] [CrossRef]
- Amcoff, E.; Edberg, A.; Barbieri, H.E.; Lindroos, A.K.; Nalsén, C.; Pearson, M.; Lemming, E.W. Riksmaten-Vuxna 2010-11: Food and Nutrition Intake among Adults in Sweden; Swedish Food Agency: Uppsala, Sweden, 2012. [Google Scholar]
- Francis, C.Y.; Morris, J.; Whorwell, P.J. The Irritable Bowel Severity Scoring System: A Simple Method of Monitoring Irritable Bowel Syndrome and Its Progress. Aliment. Pharmacol. Ther. 1997, 11, 395–402. [Google Scholar] [CrossRef]
- Ware, J.E.; Sherbourne, C.D. The MOS 36-Item Short-Form Health Survey (SF-36). I. Conceptual Framework and Item Selection. Med. Care 1992, 30, 473–483. [Google Scholar] [CrossRef]
- Lewis, S.J.; Heaton, K.W. Stool Form Scale as a Useful Guide to Intestinal Transit Time. Scand. J. Gastroenterol. 1997, 32, 920–924. [Google Scholar] [CrossRef]
- Sloots, C.E.J.; Poen, A.C.; Kerstens, R.; Stevens, M.; De Pauw, M.; Van Oene, J.C.; Meuwissen, S.G.M.; Felt-Bersma, R.J.F. Effects of Prucalopride on Colonic Transit, Anorectal Function and Bowel Habits in Patients with Chronic Constipation. Aliment. Pharmacol. Ther. 2002, 16, 759–767. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Westerhuis, J.A.; Rosén, J.; Landberg, R.; Brunius, C. Variable Selection and Validation in Multivariate Modelling. Bioinformatics 2019, 35, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Filzmoser, P.; Liebmann, B.; Varmuza, K. Repeated Double Cross Validation. J. Chemom. 2009, 23, 160–171. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; van Velzen, E.J.J.; Hoefsloot, H.C.J.; Smilde, A.K. Multivariate Paired Data Analysis: Multilevel PLSDA versus OPLSDA. Metabolomics 2010, 6, 119–128. [Google Scholar] [CrossRef]
- Nordin, E. Effects of FODMAPs and Gluten on Irritable Bowel Syndrome-from Self-Reported Symptoms to Molecular Profiling [Dissertation]; Chalmers University of Technology: Gothenburg, Sweden, 2023. [Google Scholar]
- Schär, M.Y.; Corona, G.; Soycan, G.; Dine, C.; Kristek, A.; Alsharif, S.N.; Behrends, V.; Lovegrove, A.; Shewry, P.R.; Spencer, J.P. Excretion of Avenanthramides, Phenolic Acids and Their Major Metabolites Following Intake of Oat Bran. Mol. Nutr. Food Res. 2018, 62, 1700499. [Google Scholar] [CrossRef]
- Roowi, S.; Mullen, W.; Edwards, C.A.; Crozier, A. Yoghurt Impacts on the Excretion of Phenolic Acids Derived from Colonic Breakdown of Orange Juice Flavanones in Humans. Mol. Nutr. Food Res. 2009, 53 (Suppl. S1), S68–S75. [Google Scholar] [CrossRef]
- Velikonja, A.; Lipoglavšek, L.; Zorec, M.; Orel, R.; Avguštin, G. Alterations in Gut Microbiota Composition and Metabolic Parameters after Dietary Intervention with Barley Beta Glucans in Patients with High Risk for Metabolic Syndrome Development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef]
- Iversen, K.N.; Dicksved, J.; Zoki, C.; Fristedt, R.; Pelve, E.A.; Langton, M.; Landberg, R. The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study). Nutrients 2022, 14, 1669. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic Inulin-Type Fructans Induce Specific Changes in the Human Gut Microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef] [PubMed]
- Beisner, J.; Gonzalez-Granda, A.; Basrai, M.; Damms-Machado, A.; Bischoff, S.C. Fructose-Induced Intestinal Microbiota Shift Following Two Types of Short-Term High-Fructose Dietary Phases. Nutrients 2020, 12, 3444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ouyang, Y.; Li, H.; Shen, L.; Ni, Y.; Fang, Q.; Wu, G.; Qian, L.; Xiao, Y.; Zhang, J.; et al. Metabolic Phenotypes and the Gut Microbiota in Response to Dietary Resistant Starch Type 2 in Normal-Weight Subjects: A Randomized Crossover Trial. Sci. Rep. 2019, 9, 4376. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Sun, C.; Tang, X.; Zhang, X.; Han, D.; Liang, S.; Qu, R.; Hui, X.; Shan, Y.; Hu, L.; et al. Anti-Inflammatory and Intestinal Microbiota Modulation Properties of Jinxiang Garlic (Allium Sativum L.) Polysaccharides toward Dextran Sodium Sulfate-Induced Colitis. J. Agric. Food Chem. 2020, 68, 12295–12309. [Google Scholar] [CrossRef] [PubMed]
- Genoni, A.; Christophersen, C.T.; Lo, J.; Coghlan, M.; Boyce, M.C.; Bird, A.R.; Lyons-Wall, P.; Devine, A. Long-Term Paleolithic Diet Is Associated with Lower Resistant Starch Intake, Different Gut Microbiota Composition and Increased Serum TMAO Concentrations. Eur. J. Nutr. 2020, 59, 1845–1858. [Google Scholar] [CrossRef]
- Djekic, D.; Shi, L.; Brolin, H.; Carlsson, F.; Särnqvist, C.; Savolainen, O.; Cao, Y.; Bäckhed, F.; Tremaroli, V.; Landberg, R.; et al. Effects of a Vegetarian Diet on Cardiometabolic Risk Factors, Gut Microbiota, and Plasma Metabolome in Subjects With Ischemic Heart Disease: A Randomized, Crossover Study. J. Am. Heart Assoc. 2020, 9, e016518. [Google Scholar] [CrossRef]
- Cronin, P.; Joyce, S.A.; O’toole, P.W.; O’connor, E.M. Dietary Fibre Modulates the Gut Microbiota. Nutrients 2021, 13, 1655. [Google Scholar] [CrossRef]
- Eriksen, A.K.; Brunius, C.; Mazidi, M.; Hellström, P.M.; Risérus, U.; Iversen, K.N.; Fristedt, R.; Sun, L.; Huang, Y.; Nørskov, N.P.; et al. Effects of Whole-Grain Wheat, Rye, and Lignan Supplementation on Cardiometabolic Risk Factors in Men with Metabolic Syndrome: A Randomized Crossover Trial. Am. J. Clin. Nutr. 2020, 111, 864–876. [Google Scholar] [CrossRef]
- van Loo, J.; Cummings, J.; Delzenne, N.; Englyst, H.; Franck, A.; Hopkins, M.; Kok, N.; Macfarlane, G.; Newton, D.; Quigley, M.; et al. Functional Food Properties of Non-Digestible Oligosaccharides: A Consensus Report from the ENDO Project (DGXII AIRII-CT94-1095). Br. J. Nutr. 1999, 81, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Hoffman, K.L.; Chen, J.S.; Shivappa, N.; Sood, A.; Browman, G.J.; Dirba, D.D.; Hanash, S.; Wei, P.; Hebert, J.R.; et al. Dietary Inflammatory Potential in Relation to the Gut Microbiome: Results from a Cross-Sectional Study. Br. J. Nutr. 2020, 124, 931. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D. The Gut Microbiota Manages Host Metabolism. Nat. Rev. Endocrinol. 2013, 10, 74–76. [Google Scholar] [CrossRef]
- Azcarate-Peril, M.A.; Ritter, A.J.; Savaiano, D.; Monteagudo-Mera, A.; Anderson, C.; Magness, S.T.; Klaenhammer, T.R. Impact of Short-Chain Galactooligosaccharides on the Gut Microbiome of Lactose-Intolerant Individuals. Proc. Natl. Acad. Sci. USA 2017, 114, E367–E375. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The Human Gut Bacteria Christensenellaceae Are Widespread, Heritable, and Associated with Health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed]
- Latorre-Pérez, A.; Hernández, M.; Iglesias, J.R.; Morán, J.; Pascual, J.; Porcar, M.; Vilanova, C.; Collado, L. The Spanish Gut Microbiome Reveals Links between Microorganisms and Mediterranean Diet. Sci. Rep. 2021, 11, 21602. [Google Scholar] [CrossRef]
- Liu, B.; Ye, D.; Yang, H.; Song, J.; Sun, X.; He, Z.; Mao, Y.; Hao, G. Assessing the Relationship between Gut Microbiota and Irritable Bowel Syndrome: A Two-Sample Mendelian Randomization Analysis. BMC Gastroenterol. 2023, 23, 150. [Google Scholar] [CrossRef] [PubMed]
- Hollister, E.B.; Oezguen, N.; Chumpitazi, B.P.; Luna, R.A.; Weidler, E.M.; Rubio-Gonzales, M.; Dahdouli, M.; Cope, J.L.; Mistretta, T.A.; Raza, S.; et al. Leveraging Human Microbiome Features to Diagnose and Stratify Children with Irritable Bowel Syndrome. J. Mol. Diagn. 2019, 21, 449. [Google Scholar] [CrossRef]
- Pike, B.L.; Paden, K.A.; Alcala, A.N.; Jaep, K.M.; Gormley, R.P.; Maue, A.C.; Christmann, B.S.; Elson, C.O.; Riddle, M.S.; Porter, C.K. Immunological Biomarkers in Postinfectious Irritable Bowel Syndrome. J. Travel. Med. 2015, 22, 242–250. [Google Scholar] [CrossRef]
- Caminero, A.; Nistal, E.; Herrán, A.R.; Pérez-Andrés, J.; Vaquero, L.; Vivas, S.; Ruíz de Morales, J.M.; Casqueiro, J. Gluten Metabolism in Humans. Involvement of the Gut Microbiota. In Wheat and Rice in Disease Prevention and Health; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 157–170. ISBN 9780124017160. [Google Scholar]
- Pittayanon, R.; Lau, J.T.; Yuan, Y.; Leontiadis, G.I.; Tse, F.; Surette, M.; Moayyedi, P. Gut Microbiota in Patients With Irritable Bowel Syndrome—A Systematic Review. Gastroenterology 2019, 157, 97–108. [Google Scholar] [CrossRef]
- Chey, W.; Menees, S. The Gut Microbiome and Irritable Bowel Syndrome [Version 1; Referees: 3 Approved]. F1000Res 2018, 7, 1–10. [Google Scholar]
- Elsenbruch, S.; Enck, P. Placebo Effects and Their Determinants in Gastrointestinal Disorders. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Reinke, S.N.; Ali, A.; Palmer, D.J.; Christophersen, C.T. Fecal Sample Collection Methods and Time of Day Impact Microbiome Composition and Short Chain Fatty Acid Concentrations. Sci. Rep. 2021, 11, 13964. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, M.; Engelsen, S.B.; Dragsted, L.O. LC-MS Metabolomics Top-down Approach Reveals New Exposure and Effect Biomarkers of Apple and Apple-Pectin Intake. Metabolomics 2012, 8, 64–73. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Liskova, A.; Kubatka, P.; Büsselberg, D. Enzymatic Metabolism of Flavonoids by Gut Microbiota and Its Impact on Gastrointestinal Cancer. Cancers 2021, 13, 3934. [Google Scholar] [CrossRef]
- Carregosa, D.; Pinto, C.; Ávila-Gálvez, M.Á.; Bastos, P.; Berry, D.; Santos, C.N. A Look beyond Dietary (Poly)Phenols: The Low Molecular Weight Phenolic Metabolites and Their Concentrations in Human Circulation. Compr. Rev. Food Sci. Food Saf. 2022, 21, 3931–3962. [Google Scholar] [CrossRef]
- Dodd, D.; Spitzer, M.H.; van Treuren, W.; Merrill, B.D.; Hryckowian, A.J.; Higginbottom, S.K.; Le, A.; Cowan, T.M.; Nolan, G.P.; Fischbach, M.A.; et al. A Gut Bacterial Pathway Metabolizes Aromatic Amino Acids into Nine Circulating Metabolites. Nature 2017, 551, 648. [Google Scholar] [CrossRef]
- Bernstein, H.; Bernstein, C.; Payne, C.M.; Dvorakova, K.; Garewal, H. Bile Acids as Carcinogens in Human Gastrointestinal Cancers. Mutat. Res. 2005, 589, 47–65. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; De los Reyes-Gavilán, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and Their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Henström, M.; Diekmann, L.; Bonfiglio, F.; Hadizadeh, F.; Kuech, E.M.; Von Köckritz-Blickwede, M.; Thingholm, L.B.; Zheng, T.; Assadi, G.; DIerks, C.; et al. Functional Variants in the Sucrase-Isomaltase Gene Associate with Increased Risk of Irritable Bowel Syndrome. Gut 2018, 67, 263–270. [Google Scholar] [CrossRef]
- Schmidt, T.S.B.; Raes, J.; Bork, P. The Human Gut Microbiome: From Association to Modulation. Cell 2018, 172, 1198–1215. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut Microbiota in Human Metabolic Health and Disease. Nat. Rev. Microbiol. 2020, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the Normal Gut Microbiota. World J. Gastroenterol. 2015, 21, 8836–8847. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.A.M.; Merker, A.; Nilsson, P.; Sørensen, H.; Åman, P. Chemical composition of the potential new oil crops Barbarea vulgaris, Barbarea verna and Lepidium campestre. J. Sci. Food Agric. 1999, 79, 179–186. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| FODMAPs | Gluten | Placebo | p-Value | |
|---|---|---|---|---|
| Richness | 436.77 (421.6–451.94) | 447.93 (432.76–463.1) | 447.14 (431.97–462.31) | 0.10 |
| Shannon’s diversity index | 4.59 (4.51–4.66) | 4.63 (4.56–4.7) | 4.63 (4.56–4.71) | 0.20 |
| Simpson’s diversity index | 0.97 (0.97–0.98) | 0.97 (0.97–0.98) | 0.97 (0.97–0.98) | 0.76 |
| Inv Simpson’s diversity index | 46.84 (42.78–50.89) | 47.83 (43.78–51.89) | 47.92 (43.87–51.98) | 0.81 |
| SCFA | FODMAPs | Gluten | Placebo | p-Value |
|---|---|---|---|---|
| FECES (μmol/g- dry weight) | ||||
| Acetate | 85.82 (74.3–99.14) | 78.6 (68.16–90.65) | 83.53 (72.09–96.8) | 0.46 |
| Propionate | 28.45 (24.71–32.76) | 25.78 (22.42–29.63) | 27.57 (23.87–31.84) | 0.35 |
| Butyrate | 34.8 (29.08–41.64) | 31.26 (26.17–37.43) | 33.91 (28.24–40.73) | 0.43 |
| Isobutyrate | 2.27 (1.85–2.78) | 1.96 (1.6–2.39) | 2.55 (2.06–3.14) | 0.18 |
| Succinate | 1.3 (1.01–1.67) | 1.03 (0.81–1.33) | 1.06 (0.82–1.37) | 0.23 |
| Valerate | 4.45 (3.94–5.03) | 4.17 (3.7–4.7) | 4.65 (4.1–5.27) | 0.28 |
| Isovalerate | 2.47 b (2.21–2.77) | 2.09 a (1.87–2.34) | 2.52 b (2.25–2.84) | 0.01 |
| Caproate | 0.88 (0.63–1.22) | 0.83 (0.60–1.14) | 0.96 (0.69–1.33) | 0.57 |
| PLASMA (µmol/L) | ||||
| Formate | 195.37 (175.81–217.11) | 195.71 (176.11–217.49) | 196.4 (176.74–218.25) | 0.99 |
| Acetate | 56.39 (49.9–63.72) | 57.77 (51.12–65.29) | 61.02 (53.99–68.96) | 0.47 |
| Propionate | 0.72 (0.58–0.9) | 0.69 (0.56–0.85) | 0.75 (0.61–0.92) | 0.72 |
| Butyrate | 0.36 (0.3–0.43) | 0.34 (0.28–0.41) | 0.32 (0.27–0.39) | 0.53 |
| Isobutyrate | 0.21 a (0.18–0.24) | 0.24 b (0.21–0.27) | 0.25 b (0.22–0.28) | 0.04 |
| Succinate | 3.09 (2.87–3.33) | 3.12 (2.89–3.36) | 3.17 (2.94–3.41) | 0.75 |
| Valerate | 0.05 (0.04–0.06) | 0.05 (0.04–0.06) | 0.06 (0.05–0.07) | 0.42 |
| Isovalerate | 0.19 (0.16–0.23) | 0.19 (0.16–0.23) | 0.22 (0.19–0.27) | 0.12 |
| Caproate | 0.14 (0.12–0.17) | 0.15 (0.12–0.17) | 0.16 (0.13–0.19) | 0.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nordin, E.; Hellström, P.M.; Dicksved, J.; Pelve, E.; Landberg, R.; Brunius, C. Effects of FODMAPs and Gluten on Gut Microbiota and Their Association with the Metabolome in Irritable Bowel Syndrome: A Double-Blind, Randomized, Cross-Over Intervention Study. Nutrients 2023, 15, 3045. https://doi.org/10.3390/nu15133045
Nordin E, Hellström PM, Dicksved J, Pelve E, Landberg R, Brunius C. Effects of FODMAPs and Gluten on Gut Microbiota and Their Association with the Metabolome in Irritable Bowel Syndrome: A Double-Blind, Randomized, Cross-Over Intervention Study. Nutrients. 2023; 15(13):3045. https://doi.org/10.3390/nu15133045
Chicago/Turabian StyleNordin, Elise, Per M. Hellström, Johan Dicksved, Erik Pelve, Rikard Landberg, and Carl Brunius. 2023. "Effects of FODMAPs and Gluten on Gut Microbiota and Their Association with the Metabolome in Irritable Bowel Syndrome: A Double-Blind, Randomized, Cross-Over Intervention Study" Nutrients 15, no. 13: 3045. https://doi.org/10.3390/nu15133045
APA StyleNordin, E., Hellström, P. M., Dicksved, J., Pelve, E., Landberg, R., & Brunius, C. (2023). Effects of FODMAPs and Gluten on Gut Microbiota and Their Association with the Metabolome in Irritable Bowel Syndrome: A Double-Blind, Randomized, Cross-Over Intervention Study. Nutrients, 15(13), 3045. https://doi.org/10.3390/nu15133045