
Preventative Effects of Cordyceps cicadae Mycelial Extracts on the Early-Stage Development of Cataracts in UVB-Induced Mice Cataract Model

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
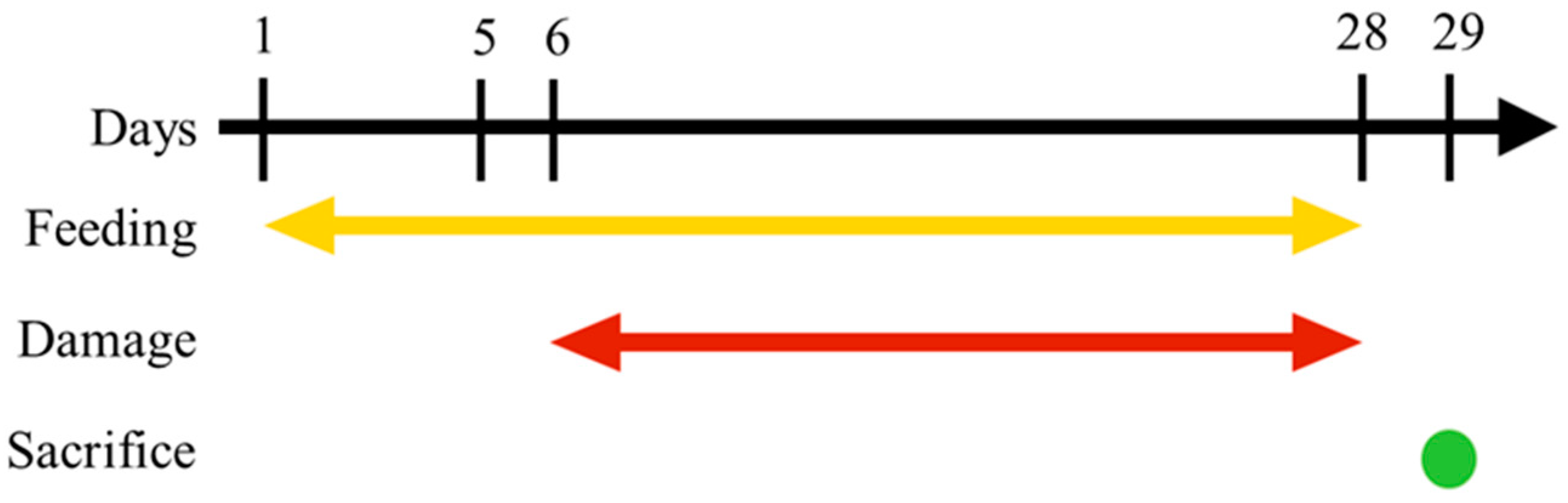
2.2. Animals and Feeding
2.3. UVB Damage Induction
2.4. Lens and Eyeball Imaging Test
2.5. Immunohistochemistry (IHC) Stainning
2.6. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Assay
3. Results
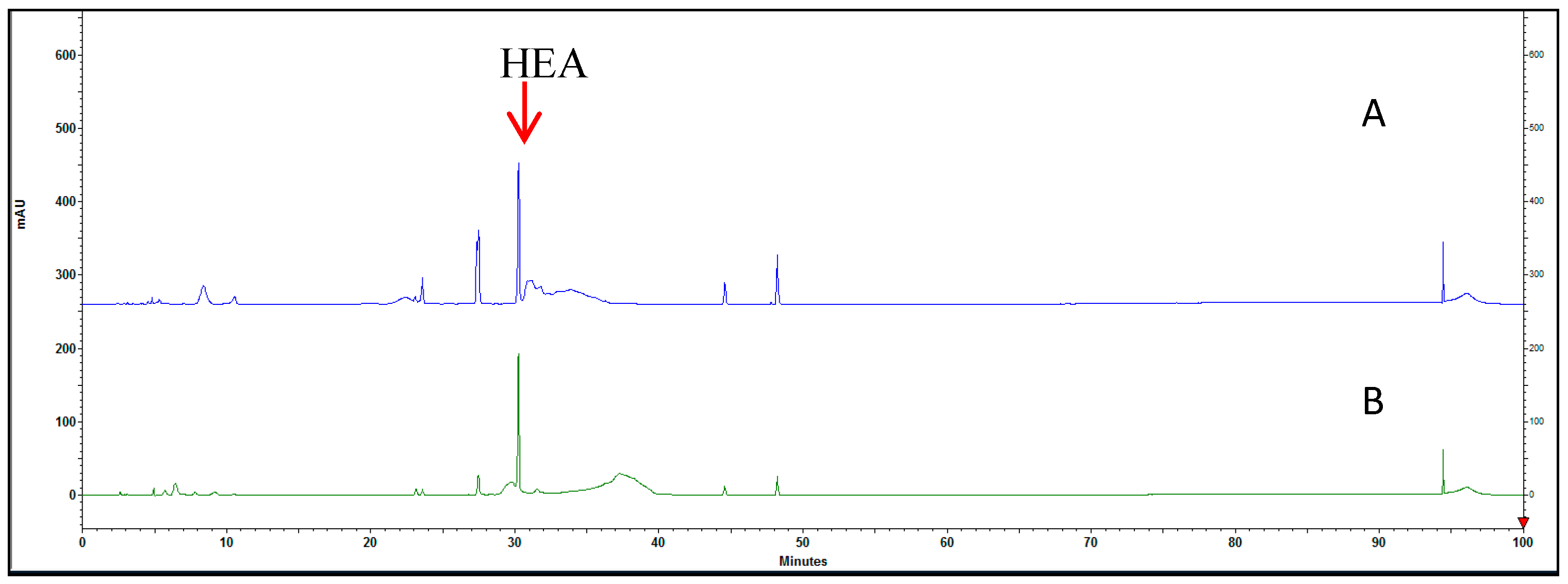
3.1. Proximate Composition Analysis of HEA in C. cicadar Mycelial Water and Ethanol Extracts
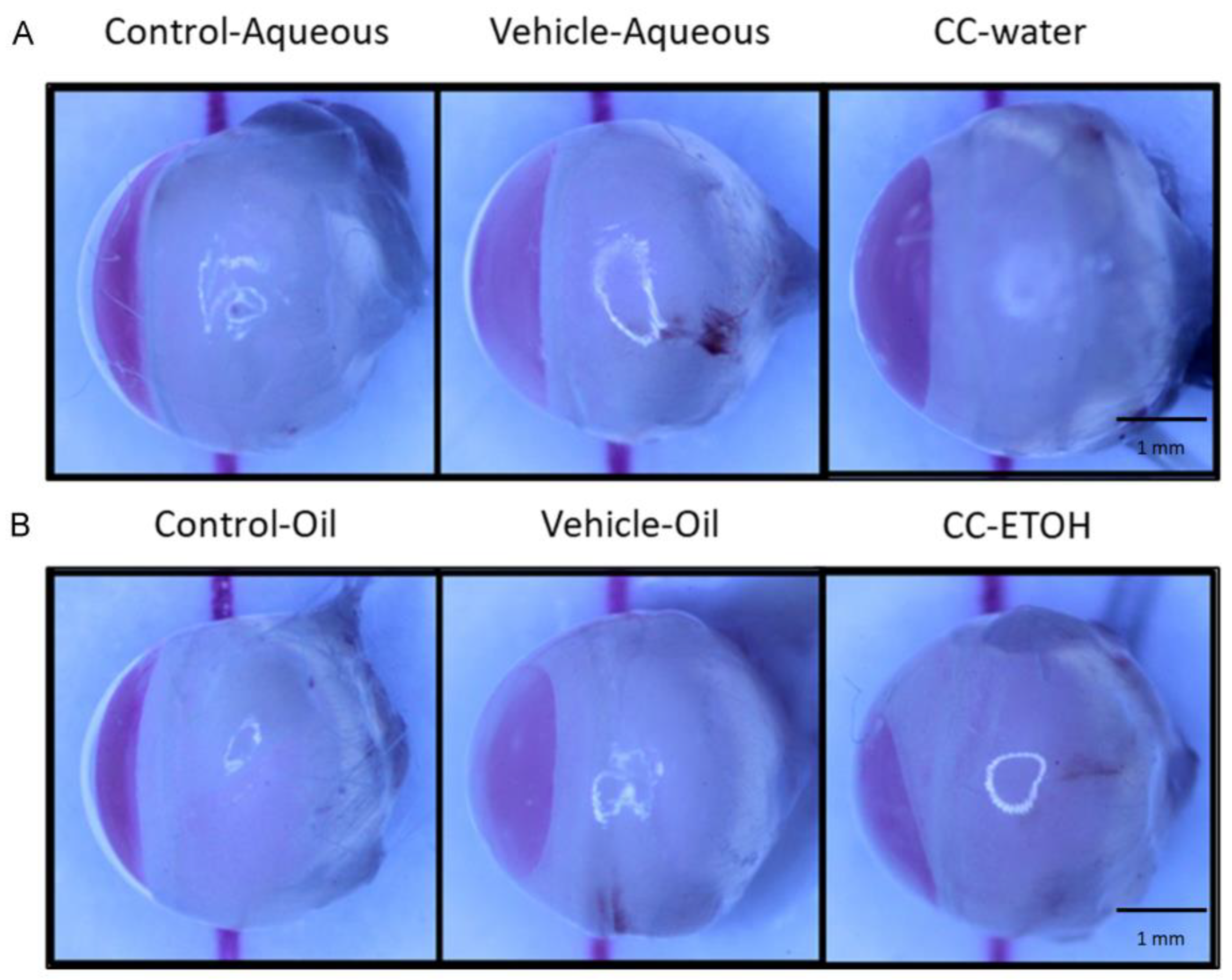
3.2. C. cicadae Ameliorate the Severity of Lens Opacity in Cataract
3.3. C. cicadae Decelerate the Development of Cataract
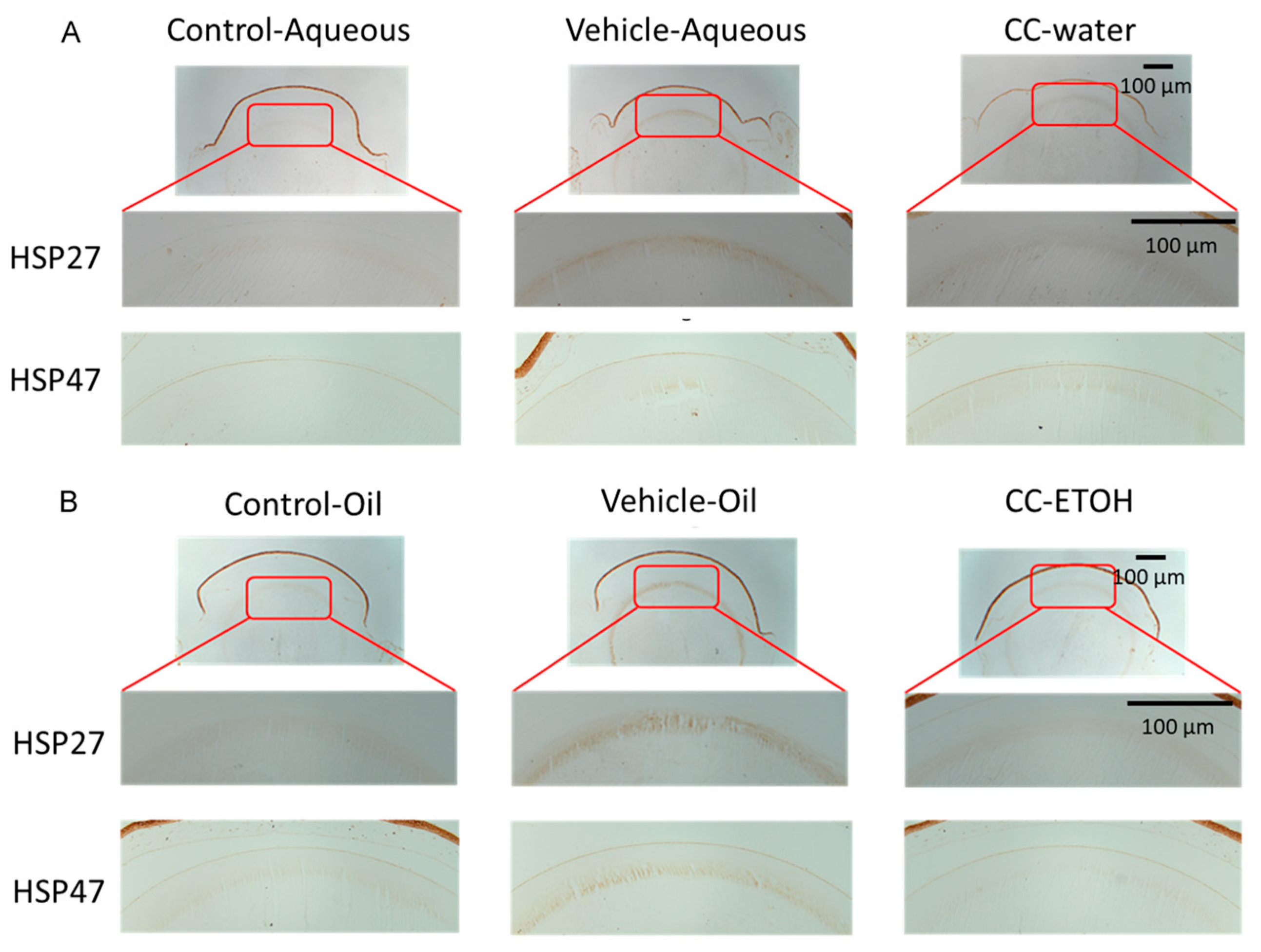
3.4. C. cicadae Mycelium Suppres HSPs in the Cortex of Crystalline Lens
3.5. C. cicadae Mycelium Suppress HSPs in hLECs
3.6. C. cicadae Mycelium Suppres Apoptosis of hLECs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.-C.; Wilkins, M.; Kim, T.; Malyugin, B.; Mehta, J.S. Cataracts. Lancet 2017, 390, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Asbell, P.A.; Dualan, I.; Mindel, J.; Brocks, D.; Ahmad, M.; Epstein, S. Age-Related Cataract. Lancet 2005, 365, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Ang, M.J.; Afshari, N.A. Cataract and Systemic Disease: A Review. Clin. Exp. Ophthalmol. 2021, 49, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Lakhani, N. Cataracts. Prim. Care 2015, 42, 409–423. [Google Scholar] [CrossRef]
- Sheppard, A.L.; Wolffsohn, J.S. Digital Eye Strain: Prevalence, Measurement and Amelioration. BMJ Open Ophthalmol. 2018, 3, e000146. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Tejedor, J.; Marchena, M.; Ramírez, L.; García-Ayuso, D.; Gómez-Vicente, V.; Sánchez-Ramos, C.; de la Villa, P.; Germain, F. Removal of the Blue Component of Light Significantly Decreases Retinal Damage after High Intensity Exposure. PLoS ONE 2018, 13, e0194218. [Google Scholar] [CrossRef]
- Chen, X.; Li, P.; Zhang, G.; Kang, L.; Qin, B.; Mao, X.; Qin, M.; Cao, Y.; Wang, Y.; Guan, H. Comprehensive Profiling of Proteome and Ubiquitome Changes in Human Lens Epithelial Cell Line after Ultraviolet-B Irradiation. ACS Omega 2020, 5, 32171–32182. [Google Scholar] [CrossRef]
- Yamaoka, R.; Kanada, F.; Nagaya, M.; Takashima, M.; Takamura, Y.; Inatani, M.; Oki, M. Analysis of Cataract-Regulated Genes Using Chemical DNA Damage Induction in a Rat Ex Vivo Model. PLoS ONE 2022, 17, e0273456. [Google Scholar] [CrossRef]
- Sorte, K.; Sune, P.; Bhake, A.; Shivkumar, V.; Gangane, N.; Basak, A. Quantitative Assessment of DNA Damage Directly in Lens Epithelial Cells from Senile Cataract Patients. Mol. Vis. 2011, 17, 1–6. [Google Scholar]
- Dubrez, L.; Causse, S.; Bonan, N.B.; Dumétier, B.; Garrido, C. Heat-Shock Proteins: Chaperoning DNA Repair. Oncogene 2020, 39, 516–529. [Google Scholar] [CrossRef]
- Shan, Q.; Ma, F.; Wei, J.; Li, H.; Ma, H.; Sun, P. Physiological Functions of Heat Shock Proteins. Curr. Protein Pept. Sci. 2020, 21, 751–760. [Google Scholar] [CrossRef]
- Urbak, L.; Henrik, V. Heat Shock Proteins in the Human Eye. Int. J. Proteom. 2010, 2010, 479571. [Google Scholar] [CrossRef] [Green Version]
- Tedesco, B.; Cristofani, R.; Ferrari, V.; Cozzi, M.; Rusmini, P.; Casarotto, E.; Chierichetti, M.; Mina, F.; Galbiati, M.; Piccolella, M.; et al. Insights on Human Small Heat Shock Proteins and Their Alterations in Diseases. Front. Mol. Biosci. 2022, 9, 842149. [Google Scholar] [CrossRef]
- Bagchi, M.; Katar, M.; Maisel, H. Heat Shock Proteins of Adult and Embryonic Human Ocular Lenses. J. Cell. Biochem. 2002, 84, 278–284. [Google Scholar] [CrossRef]
- Zhang, Y.; Gong, J.; Zhang, L.; Xue, D.; Liu, H.; Liu, P. Genetic Polymorphisms of Hsp70 in Age-Related Cataract. Cell Stress Chaperones 2013, 18, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.-B.; Yu, H.; Ge, F.; Yang, J.-Y.; Chen, Z.-H.; Wang, Y.-B.; Dai, Y.-D.; Adams, A. Distribution of Nucleosides in Populations of Cordyceps Cicadae. Molecules 2014, 19, 6123–6141. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.-P.; Hwang, T.-L.; Chan, Y.; El-Shazly, M.; Wu, T.-Y.; Lo, I.-W.; Hsu, Y.-M.; Lai, K.-H.; Hou, M.-F.; Yuan, S.-S.; et al. Research and Development of Cordyceps in Taiwan. Food Sci. Hum. Wellness 2016, 5, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Weng, S.C.; Chou, C.J.; Lin, L.C.; Tsai, W.J.; Kuo, Y.C. Immunomodulatory Functions of Extracts from the Chinese Medicinal Fungus Cordyceps Cicadae. J. Ethnopharmacol. 2002, 83, 79–85. [Google Scholar] [CrossRef]
- Zhu, R.; Chen, Y.P.; Deng, Y.Y.; Zheng, R.; Zhong, Y.F.; Wang, L.; Du, L.P. Cordyceps Cicadae Extracts Ameliorate Renal Malfunction in a Remnant Kidney Model. J. Zhejiang Univ. Sci. B 2011, 12, 1024–1033. [Google Scholar] [CrossRef]
- Lu, M.Y.; Chen, C.C.; Lee, L.Y.; Lin, T.W.; Kuo, C.F. N(6)-(2-Hydroxyethyl)Adenosine in the Medicinal Mushroom Cordyceps Cicadae Attenuates Lipopolysaccharide-Stimulated Pro-Inflammatory Responses by Suppressing Tlr4-Mediated Nf-Κb Signaling Pathways. J. Nat. Prod. 2015, 78, 2452–2460. [Google Scholar] [CrossRef]
- Chen, Y.L.; Yeh, S.H.; Lin, T.W.; Chen, C.C.; Chen, C.S.; Kuo, C.F. A 90-Day Subchronic Toxicity Study of Submerged Mycelial Culture of Cordyceps Cicadae (Ascomycetes) in Rats. Int. J. Med. Mushrooms 2015, 17, 771–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.S.; Hsu, J.H.; Lin, D.P.; Chang, H.H.; Chang, W.J.; Chen, Y.L.; Chen, C.C. Safety Assessment of Hea-Enriched Cordyceps Cicadae Mycelium: A Randomized Clinical Trial. J. Am. Coll. Nutr. 2021, 40, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Chyau, C.C.; Wu, H.L.; Peng, C.C.; Huang, S.H.; Chen, C.C.; Chen, C.H.; Peng, R.Y. Potential Protection Effect of Er Homeostasis of N(6)-(2-Hydroxyethyl)Adenosine Isolated from Cordyceps Cicadae in Nonsteroidal Anti-Inflammatory Drug-Stimulated Human Proximal Tubular Cells. Int. J. Mol. Sci. 2021, 22, 1577. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qin, A.; Xiao, F.; Olatunji, O.J.; Zhang, S.; Pan, D.; Han, W.; Wang, D.; Ni, Y. N6-(2-Hydroxyethyl)-Adenosine from Cordyceps Cicadae Protects against Diabetic Kidney Disease Via Alleviation of Oxidative Stress and Inflammation. J. Food Biochem. 2019, 43, e12727. [Google Scholar] [CrossRef]
- Wen, Y.T.; Jhou, B.Y.; Hsu, J.H.; Fu, H.I.; Chen, Y.L.; Shih, Y.C.; Chen, C.C.; Tsai, R.K. Neuroprotective Effects of Cordyceps Cicadae (Ascomycetes) Mycelium Extract in the Rat Model of Optic Nerve Crush. Int. J. Med. Mushrooms 2022, 24, 41–48. [Google Scholar] [CrossRef]
- Li, I.C.; Lin, S.; Tsai, Y.T.; Hsu, J.H.; Chen, Y.L.; Lin, W.H.; Chen, C.C. Cordyceps Cicadae Mycelia and Its Active Compound Hea Exert Beneficial Effects on Blood Glucose in Type 2 Diabetic Db/Db Mice. J. Sci. Food Agric. 2019, 99, 606–612. [Google Scholar] [CrossRef]
- Chang, C.Y.; Yang, P.X.; Yu, T.L.; Lee, C.L. Cordyceps Cicadae Nttu 868 Mycelia Fermented with Deep Ocean Water Minerals Prevents D-Galactose-Induced Memory Deficits by Inhibiting Oxidative Inflammatory Factors and Aging-Related Risk Factors. Nutrients 2023, 15, 1968. [Google Scholar] [CrossRef]
- Lin, T.-Y.; Chang, H.-H.; Tang, Y.-J.; Chen, C.-C.; Lee, L.-Y.; Lin, D.P.-C. Fermented Cordyceps Cicadae Mycelia Extracts Ameliorate Dry Eye Symptoms through Reduction of Cornea Epithelial Cell Apoptosis and Maintenance of Conjunctival Goblet Cells in a Mouse Dry Eye Model. J. Food Nutr. Res. 2017, 5, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.H.; Chang, W.J.; Jhou, B.Y.; Kuo, S.Y.; Hsu, J.H.; Chen, Y.L.; Chen, C.C.; Lin, D.P. Efficacy of Cordyceps Cicadae (Ascomycota) Mycelium Supplementation for Amelioration of Dry Eye Symptoms: A Randomized, Double-Blind Clinical Pilot Study. Int. J. Med. Mushrooms 2022, 24, 57–67. [Google Scholar] [CrossRef]
- Horng, C.T.; Yang, Y.L.; Chen, C.C.; Huang, Y.S.; Chen, C.; Chen, F.A. Intraocular Pressure-Lowering Effect of Cordyceps Cicadae Mycelia Extract in a Glaucoma Rat Model. Int. J. Med. Sci. 2021, 18, 1007–1014. [Google Scholar] [CrossRef]
- Fu, H.-I.; Hsu, J.-H.; Li, T.-J.; Yeh, S.-H.; Chen, C.-C. Safety Assessment of Hea-Enriched Cordyceps Cicadae Mycelia on the Central Nervous System (Cns), Cardiovascular System, and Respiratory System in Icr Male Mice. Food Sci. Nutr. 2021, 9, 4905–4915. [Google Scholar] [CrossRef]
- Deng, J.S.; Jiang, W.P.; Chen, C.C.; Lee, L.Y.; Li, P.Y.; Huang, W.C.; Liao, J.C.; Chen, H.Y.; Huang, S.S.; Huang, G.J. Cordyceps Cicadae Mycelia Ameliorate Cisplatin-Induced Acute Kidney Injury by Suppressing the Tlr4/Nf-Κb/Mapk and Activating the Ho-1/Nrf2 and Sirt-1/Ampk Pathways in Mice. Oxid. Med. Cell. Longev. 2020, 2020, 7912763. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, T.; Li, C.; Xie, L.; Li, N.; Hou, T.; Wang, Y.; Wang, B. Potential Therapeutic Effects of Cordyceps Cicadae and Paecilomyces Cicadae on Adenine-Induced Chronic Renal Failure in Rats and Their Phytochemical Analysis. Drug Des. Devel Ther. 2019, 13, 103–117. [Google Scholar] [CrossRef] [Green Version]
- Arrigo, A.P.; Simon, S. Expression and Functions of Heat Shock Proteins in the Normal and Pathological Mammalian Eye. Curr. Mol. Med. 2010, 10, 776–793. [Google Scholar] [CrossRef]
- Goodman, D.; Ness, S. The Role of Oxidative Stress in the Aging Eye. Life 2023, 13, 837. [Google Scholar] [CrossRef]
- Cicinelli, M.V.; Buchan, J.C.; Nicholson, M.; Varadaraj, V.; Khanna, R.C. Cataracts. Lancet 2023, 401, 377–389. [Google Scholar] [CrossRef]
- Ivanov, I.V.; Mappes, T.; Schaupp, P.; Lappe, C.; Wahl, S. Ultraviolet Radiation Oxidative Stress Affects Eye Health. J. Biophotonics 2018, 11, e201700377. [Google Scholar] [CrossRef] [Green Version]
- Zhuansun, W.; Xu, J.; Liu, H.; Zhao, Y.; Chen, L.; Shan, S.; Song, S.; Zhang, H.; Dong, T.; Zeng, H.; et al. Optimisation of the Production of a Selenium-Enriched Polysaccharide from Cordyceps Cicadae S1 and Its Structure and Antioxidant Activity. Front. Nutr. 2022, 9, 1032289. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, X.; Ge, Q.; Li, J.; Wang, D.; Wei, Y.; Ouyang, Z. Antioxidant and Anti-Aging Activities of Polysaccharides from Cordyceps Cicadae. Int. J. Biol. Macromol. 2020, 157, 394–400. [Google Scholar] [CrossRef]
- Ohe, H.; Harima, S.; Kubo, M. The Classification and Therapeutical Methods of Cataract on Chinese Medicine in the Modern Times. Yakushigaku Zasshi 1995, 30, 125–133. [Google Scholar]
- Yao, Q.; Zhou, Y.; Yang, Y.; Cai, L.; Xu, L.; Han, X.; Guo, Y.; Li, P.A. Activation of Sirtuin1 by Lyceum Barbarum Polysaccharides in Protection against Diabetic Cataract. J. Ethnopharmacol. 2020, 261, 113165. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, F.; Hou, T.; Geng, C.; Xu, T.; Han, B.; Liu, D. Dendrobium Huoshanense C.Z.Tang Et S.J.Cheng: A Review of Its Traditional Uses, Phytochemistry, and Pharmacology. Front. Pharmacol. 2022, 13, 920823. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.J.; Danesh-Meyer, H.V. Caffeine and the Eye. Surv. Ophthalmol. 2019, 64, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Daines, B.S.; Kent, A.R.; McAleer, M.S.; Crosson, C.E. Intraocular Adenosine Levels in Normal and Ocular-Hypertensive Patients. J. Ocul. Pharmacol. Ther. 2003, 19, 113–119. [Google Scholar] [CrossRef]
- Sooraj, K.; Shukla, S.; Kaur, R.; Titiyal, J.S.; Kaur, J. The Protective Role of Hsp27 in Ocular Diseases. Mol. Biol. Rep. 2022, 49, 5107–5115. [Google Scholar] [CrossRef]
- Nagata, K. Hsp47: A Collagen-Specific Molecular Chaperone. Trends Biochem. Sci. 1996, 21, 22–26. [Google Scholar] [CrossRef]
- Bagchi, M.; Katar, M.; Maisel, H. A Heat Shock Transcription Factor Like Protein in the Nuclear Matrix Compartment of the Tissue Cultured Mammalian Lens Epithelial Cell. J. Cell. Biochem. 2001, 80, 382–387. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Z.; Bao, X.; Qin, Y.; Taylor, A.; Shang, F.; Wu, M. Chip Knockdown Reduced Heat Shock Response and Protein Quality Control Capacity in Lens Epithelial Cells. Curr. Mol. Med. 2015, 15, 652–662. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Name | Feed | UVB Damage |
|---|---|---|---|
| Aqueous | Control-Aqueous | 0.9% NaCl solution | No damage |
| Vehicle-Aqueous | 0.9% NaCl solution | Damage, days 6–28 | |
| C. cicadae mycelial water extract | Extract in 0.9% NaCl solution | Damage, days 6–28 | |
| Oil | Control-Oil | SBO | No damage |
| Vehicle-Oil | SBO | Damage, days 6–28 | |
| C. cicadae mycelial ethanol extract | Extract in SBO | Damage, days 6–28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.-H.; Chang, J.-W.; Jhou, B.-Y.; Hsu, J.-H.; Li, T.-J.; Lee, L.-Y.; Chen, Y.-L.; Chang, H.-H.; Chen, C.-C.; Wu, P.-S.; et al. Preventative Effects of Cordyceps cicadae Mycelial Extracts on the Early-Stage Development of Cataracts in UVB-Induced Mice Cataract Model. Nutrients 2023, 15, 3103. https://doi.org/10.3390/nu15143103
Lu T-H, Chang J-W, Jhou B-Y, Hsu J-H, Li T-J, Lee L-Y, Chen Y-L, Chang H-H, Chen C-C, Wu P-S, et al. Preventative Effects of Cordyceps cicadae Mycelial Extracts on the Early-Stage Development of Cataracts in UVB-Induced Mice Cataract Model. Nutrients. 2023; 15(14):3103. https://doi.org/10.3390/nu15143103
Chicago/Turabian StyleLu, Tsung-Han, Jun-Way Chang, Bo-Yi Jhou, Jui-Hsia Hsu, Tsung-Ju Li, Li-Ya Lee, Yen-Lien Chen, Han-Hsin Chang, Chin-Chu Chen, Pey-Shiuan Wu, and et al. 2023. "Preventative Effects of Cordyceps cicadae Mycelial Extracts on the Early-Stage Development of Cataracts in UVB-Induced Mice Cataract Model" Nutrients 15, no. 14: 3103. https://doi.org/10.3390/nu15143103