
Transcriptomic Analysis of Human Skeletal Muscle in Response to Aerobic Exercise and Protein Intake

1
Key Laboratory of Molecular Epidemiology of Hunan Province, School of Medicine, Hunan Normal University, Changsha 410013, China
2
Hunan Provincial Institute of Emergency Medicine, Hunan Provincial People’s Hospital, Changsha 410009, China
3
Department of Medicine & Therapeutics, Prince of Wales Hospital, The Chinese University of Hong Kong, Hong Kong SAR, China
*
Author to whom correspondence should be addressed.
Nutrients 2023, 15(15), 3485; https://doi.org/10.3390/nu15153485
Submission received: 16 June 2023
/
Revised: 29 July 2023
/
Accepted: 3 August 2023
/
Published: 7 August 2023
(This article belongs to the Special Issue Nutrition Role in Bone and Muscle Health)
Abstract
:This study aimed to provide a more comprehensive molecular insight into the effects of aerobic exercise (AE), protein intake (PI), and AE combined with PI on human skeletal muscle by comparing their transcriptomic profiles. Fourteen published datasets obtained from the Gene Expression Omnibus (GEO) database were used. The hub genes were identified in response to acute AE (ACTB, IL6), training AE (UBB, COL1A1), PI (EZH2), acute AE combined with PI (DDIT3), and training AE combined with PI (MYC). Both FOS and MYC were upregulated in response to acute AE, and they were, respectively, downregulated by higher PI and a combination of AE and PI. COL1A1 was upregulated by training AE but was downregulated by higher PI. Results from the gene set enrichment analysis (p < 0.05 and FDR < 25%) showed that AE and PI delivered their impacts on human skeletal muscle in analogous pathways, including aerobic respiration, mitochondrial complexes, extracellular matrix (ECM) remodeling, metabolic process, and immune/inflammatory responses, whereas, PI may attenuate the response of immune/inflammation and ECM remodeling which would be promoted by AE, irrespective of its types. Compared to PI alone, acute AE combined with PI would further promote protein turnover and synthesis, but suppress skeletal muscle contraction and movement.
1. Introduction
Muscle metabolism plays an essential role in the genesis of many pathologic conditions and chronic diseases [1]. Sarcopenia is an aging-related loss of muscle mass and/or muscle function [2], which involves adverse muscle changes across the lifetime and the increased risk of falls, fractures, frailty, and mortality [3]. The prevalence of sarcopenia and muscle atrophy is up to 29% in community-dwelling populations and 33% in long-term care populations [4], which results in a large healthcare burden [5]. The development of optimal interventions to maintain muscle and improve mobility is extremely important.
Exercise could help preclude the pathophysiological processes of muscle mass and function, and prevent or delay the development of sarcopenia [6,7,8]. Considerable evidence demonstrates that resistance exercise (RE) is the most effective in enhancing muscular strength and function performance [9,10], and RE is now recommended as the first-line non-pharmacological intervention for counteracting the deleterious consequences of sarcopenia [11]. However, besides the loss of muscle mass, the decrease in muscle mitochondria with age-related muscle atrophy [12,13] contributes significantly to an imbalance of energy homeostasis, and the development of sarcopenia [14,15,16]. Chronic conditions, such as cardiovascular diseases, may accelerate the muscle dysfunction and tissue atrophy through imparting pathological hormonal change, malnutrition, and physical inactivity [17,18], and obesity may contribute to mitochondrial dysfunction through the intramuscular accumulation of impaired β-oxidation capacity [19]. Differing from the exercise modality of RE, aerobic exercise (AE) is more effective in improving aerobic capacity, maintaining whole-body energy homeostasis [9,10,20], managing the metabolism of visceral adipose tissue in obesity [20,21], and improving cardiovascular fitness through the increase in exercise capacity and peak oxygen uptake [22,23], which would benefit muscle health in a divergent pathway. Moreover, previous studies found that AE imbued to mid- and long-term immune modulations [24] and induced strong cellular immune responses and more alternations in immune cell populations [25]. Compared to RE alone, AE appears to be more effective for maintaining and improving maximum aerobic power. Moreover, mixed training or RE in combination with AE was suggested to deliver more positive effects on muscle mass, strength, and function in older people [26,27].
Nutritional interventions have also been recommended as a non-negligible option to preserve muscle mass and function [28,29,30,31]. Double-blind RCT suggested that higher protein intake (PI) brought more benefits than lower PI in preventing sarcopenia in older people [32]. The synergistic effect of exercise and PI has been identified [33]. Besides the positive effect of the combination of RE and PI on skeletal muscle [31,34,35,36], the potential benefit brought by the combination of AE and protein feeding on skeletal muscle myofibrillar plasticity [37,38], greater gains in VO2max, and stimulation of lean mass accretion have also been reported [9,39,40,41]. However, controversies still exist over the capability of the combination of AE and dietary protein on improving skeletal muscle oxidative capacity and endurance performance [42,43,44,45].
More evidence is needed to identify whether AE or and its combination with PI could also be effective interventions for improving human skeletal muscle health. Understanding the key molecular signature characteristics underlying responses to these interventions may bring some new hints. Transcriptomic analysis can provide a unique microscopic insight to access the directions of adaptive changes and predict the underlying mechanisms. Previous studies have used transcriptomic analysis to summarize the overlap and the difference of human skeletal muscle in response to RE, AE, acute exercise, training exercise, and inactivity/disuse [39,40,41]. Nuclear Receptor Subfamily 4 Group A Member 3 (NR4A3) and SMAD Family Member 3 (SMAD3) were identified as two of the most exercise- and inactivity- and acute- and training-responsive genes, respectively [39,41]. The transcriptional signatures of disuse and RE training (RET) suggested that RET was not completely effective in countering disuse atrophy due to the fact that the mechanisms of disuse were not covered by the converse of the RET [40].
Thus, the present study aimed to highlight the divergent transcriptomic responses of human skeletal muscle to AE, PI and their combinations, and to provide more molecular evidence of these interventions for maintaining muscle health and alleviating sarcopenia.
2. Materials and Methods
2.1. Public Datasets
The Gene Expression Omnibus (GEO) database [46] was searched (up to 1 December 2022) to obtain datasets of the gene expression profiling on human skeletal muscle in response to AE, PI, and AE combined with PI. Inclusion criteria were as follows: (1) datasets using microarray were chosen; (2) studies conducted in healthy adults (i.e., free of injury and disease); (3) human muscle biopsy samples were used; and (4) studies with 5 subjects or more were included. If there was more than one record of time points for muscle biopsy, we selected the one with the minimum difference across the studies to improve comparability. The studies, which had results reported separately for gender or reported separately for different age groups, were treated as independent studies.
2.2. Data Pre-Processing
To minimize the heterogeneity across the multiple microarray platforms, quantile normalization and log 2 transformations were used for each microarray expression matrix data. If more than one probe was used for the gene detection, the probe with the maximal average expression would be taken. Whereas, if a probe was used for more than one gene, the probe would be discarded due to its low specificity. If multiple categories were used to assess the degree of PI, the category with the highest degree would be chosen to compare with the one with the lowest degree.
2.3. Identification of Differentially Expressed Genes (DEGs)
Differential expression analysis (DEA) was performed using the Benjamini–Hochberg (BH) correction for paired samples using the limma package in R (version 4.2.0) [47]. The genes with p value less than 0.05 in DEA were identified as the DEGs. These DEGs with a threshold of log2 Fold Change (log2FC) above 0 were considered to be upregulated, while those with log2FC less than 0 were downregulated.
2.4. Functional Analysis and Network Visualization of co-DEGs
Those DEGs which were identified in two or more datasets were labeled as co-differentially expressed genes (co-DEGs). Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis were performed to identify the biological functions and potential pathways of the co-DEGs. The bioconductor package “org.Hs.eg.db” and the “cluster profiler” package [48] were used for GO and KEGG pathway enrichment analysis.
Subsequently, all the co-DEGs were used to construct a gene expression network to find the interactions among these genes using Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) Database (https://cn.string-db.org/, accessed on 26 December 2022, version: 11.5). The gene with the top degree value calculated by CytoHubba plugin [49] in Cytoscape software (version 3.9.1) was considered the hub gene (i.e., the highly connected genes within a module and those significantly associated with biological function).
2.5. Gene Set Enrichment Analysis (GSEA)
GSEA among different interventions was further performed to explore genome-wide expression profiles within each dataset, without setting up an arbitrary threshold. This would provide an alternative way to screen out obvious differential biological functions induced by varied interventions within each dataset. GSEA was performed using the “cluster profiler” package. The number of permutations was set up at 10,000. Gene set was considered to be significantly enriched when the absolute value of normalized enrichment score (NES) was more than 1 with a p-value less than 0.05 and a false discovery rate (FDR) less than 25%.
3. Results
3.1. Overview of Datasets Collection
There were fourteen datasets included in the present study (Supplementary Figure S1). Those were studies of acute AE (GSE43856 [50], GSE59088 [51], GSE126296 [52], GSE59363 [53]), training AE (GSE111551, GSE72462 [54], GSE1786 [55], GSE43760 [56]), PI (GSE8441 [57], GSE9419 [58], GSE73525 [59]), acute AE combined with PI (GSE27285 [60], GSE44818 [61]), and training AE combined with PI (GSE147494 [62]). Acute AE refers to a bout of aerobic exercise, and training AE refers to regular aerobic exercise with frequency and intensity lasting for at least 2 weeks. Both GSE8441 and GSE72462 can be considered to be two independent studies by gender, GSE9419 can be considered two independent studies by age group (the younger one (<50 years) and the older one (≥50 years)). The details of included datasets were shown in Table 1.
3.2. DEGs Identification, Hub Gene Screening, and Gene Expression Network Construction
The details of the number of DEGs from each dataset and co-DEGs among different populations across interventions were summarized (Figure 1 and Figure S2). Compared to PI or AE combined with PI, more DEGs were identified in the studies delivering AE alone (Figure 1A). No co-DEG were reported for training AE, PI, and training AE combined with PI in the younger male (YM) due to only one dataset for each of them being included. Herein, the DEGs were considered the co-DEGs in these studies (Figure 1B).
The top five up- and down-regulated DEGs within each dataset and co-DEGs within each intervention among different populations in response to varied interventions were summarized (Supplementary Table S1). Nuclear Receptor Subfamily 4 Group A Member 3 (NR4A3), Pyruvate Dehydrogenase Kinase 4 (PDK4), PPARG Coactivator 1 Alpha (PPARGC1A) were consistently upregulated in response to acute AE (Supplementary Table S1). These genes were reported to contribute to the regulation of glucose metabolism, cell proliferation, differentiation and transformation, and the transcription by RNA polymerase II. MicroRNA 206 (MIR206), and Circadian-Associated Repressor of Transcription (CIART) were the most downregulated co-DEG in response to acute AE in the YM and the OM, respectively (Supplementary Table S1).
Stearoyl-CoA Desaturase (SCD), Rho GTPase Activating Protein 12 (ARHGAP12), and Asporin (ASPN) were the most upregulated genes in response to training AE in the YM, the OM, and the older female (OF), respectively (Supplementary Table S1). Ankyrin Repeat Domain 2 (ANKRD2), G Protein-Coupled Receptor 37 (GPR37), and Establishment of Sister Chromatid Cohesion N-Acetyltransferase 1 (ESCO1) were the most downregulated co-DEGs in response to training AE in the the YM, the OM, and the OF, respectively (Supplementary Table S1).
Chromosome 10 Open Reading Frame 126 (C10orf126), Secretoglobin Family 2A Member 2 (SCGB2A2), and EPH Receptor A2 (EPHA2) were the most upregulated co-DEGs in response to PI in the YM, the OM, and the OF, respectively (Supplementary Table S1). Splicing Factor Proline/Glutamine-Rich (LOC654780), Adhesion G Protein-Coupled Receptor G2 (ADGRG2), and Mannose Binding Lectin 2 (MBL2) were the most downregulated genes in response to PI in the three groups, respectively (Supplementary Table S1). Noteworthy, FOS was significantly downregulated in response to PI but was upregulated in response to acute AE.
Cell production-, proliferation-, and differentiation-related genes, including Interferon Related Developmental Regulator 1 (IFRD1) and DNA Damage Inducible Transcript 3 (DDIT3), were consistently upregulated in response to acute AE combined with PI. Protein Phosphatase 1 Regulatory Subunit 15A (PPP1R15A) was the most downregulated co-DEG in response to acute AE combined with PI. What is noteworthy is that some immune/inflammation-related genes such as C-C Motif Chemokine Ligand 2 (CCL2) and C-X-C Motif Chemokine Receptor 7 (CXCR7) were downregulated in response to acute AE combined with PI (Table S1). Genes involved in ECM remodeling, including Collagen Type I Alpha 1 Chain (COL1A1) and Collagen Type III Alpha 1 Chain (COL3A1), were downregulated both in response to PI and acute AE combined with PI (Supplementary Table S1). Similar to PI alone, FOS was downregulated in response to acute AE combined with PI. Most of the top five responded genes to PI and acute AE combined with PI were strongly associated with the immune/inflammation process and ECM remodeling.
The top upregulated and downregulated genes in response to training AE combined with PI were CCR4-NOT Transcription Complex Subunit 7 (CNOT7) and Solute Carrier Family 13 Member 2 (SLC13A2), respectively. They were reported to be related to the negative regulation of cell proliferation and the formation of kidney stones, respectively (Supplementary Table S1).
A minimum interaction score of 0.4 was set up for the gene expression network construction in the STRING database using the identified co-DEGs. The degree value was calculated and the top five genes were summarized for each network (Supplementary Tables S1 and S4). Actin Beta (ACTB) and Interleukin 6 (IL6) were the hub genes in response to acute AE in the YM and the OM, respectively. Ubiquitin B (UBB) and COL1A1 were the hub genes in response to training AE in the YM and the OF, respectively. The enhancer Of Zeste 2 Polycomb Repressive Complex 2 Subunit (EZH2), DDIT3, and MYC Proto-Oncogene (MYC) were the hub genes in response to PI, acute AE combined with PI, and training AE combined with PI in the YM, respectively (Supplementary Tables S1 and S2).
FOS, MYC, and COL1A1 identified above were upregulated by acute AE and training AE, but were downregulated by PI and acute AE combined with PI (Supplementary Table S1). Subsequently, the expression levels of these genes in the intervention and control groups within each dataset were compared across the different interventions. Compared to each own control group, the expression levels of FOS, MYC, and COL1A1 were higher in acute AE and training AE but lower in the PI-related studies, although statistical significance was not shown in all comparisons (Supplementary Figures S3–S5).
3.3. Functional Enrichment Analysis for co-DEGs
GO terms and KEGG pathways enrichment analysis were conducted to explore the biological functions of the co-DEGs. For acute AE studies, co-DEGs were mainly enriched in the terms related to starvation, peptide hormones, protein turnover and synthesis, cellular response to external stimuli, and the circadian regulation of gene expression (Figure 2A and Figure S6, Supplementary Table S3). The AGE-RAGE signaling pathway in diabetic complications and circadian rhythm was the most significant KEGG pathway in response to acute AE in the YM and the OM, respectively (Figure 3A and Figure S7, Supplementary Table S4). For training AE, the most significant GO terms in the YM, OM, and OF were fatty acid transmembrane transport, fatty acid beta-oxidation using acyl-CoA dehydrogenase, and positive regulation of smooth muscle cell proliferation, respectively (Figure 2B and Figure S6, Supplementary Table S3). No significant KEGG pathway was enriched in response to training AE (Figure 3B and Figure S7, Supplementary Table S4).
For PI in the OM, the most significant GO term was the positive regulation of vasoconstriction (Supplementary Figure S6 and Table S3). No GO term was significantly enriched in the co-DEGs of PI after BH correction in the YM and OF (Figure 2C and Figure S6, Supplementary Table S3). And, no KEGG pathway was significantly enriched by co-DEGs in response to PI (Figure 3C and Figure S7, Supplementary Table S4). The co-DEGs of acute AE combined with PI were mainly associated with the function or pathway of protein synthesis and turnover, such as endoplasmic reticulum unfolded protein response and cellular response to unfolded protein. This was similar to acute AE but not to PI alone in the YM (Figure 2D and Figure 3D, Supplementary Table S3 and S4). Few GO terms and KEGG pathways were significantly enriched by the co-DEGs from the response to training AE combined with PI (Figure 2E and Figure 3E, Supplementary Table S3 and S4).
3.4. GSEA
Consistent results were shown across the studies of acute AE. The gene sets which were significantly upregulated in response to acute AE included the cellular response to external stimuli, regulation of fatty acid oxidation, energy metabolic process, ECM remodeling, vasculature development, skeletal muscle development, and immune/inflammation process (Figure 4 and Figure S8, Supplementary Table S5).
Compared to acute AE, more profound transcriptomic changes in the ECM remodeling and immune/inflammation process were observed in response to training AE in the older people. Gene sets related to extracellular stimuli, protein turnover and synthesis, and angiogenesis regulation were not significantly enriched in response to training AE (Supplementary Table S5). Compared to the older people, mitochondrion complex, aerobic respiration, electron transport complex, and ATP synthesis were downregulated in the YM (which were upregulated in the older people) (Supplementary Figure S9 and Table S5). Gene sets related to ECM organization, basement membrane, cornified envelope, and peptide cross-linking were upregulated in response to training AE in the OM, while nuclear glucocorticoid receptor binding, nuclear receptor activity, and ligand-activated transcription factor activity were downregulated (Supplementary Figure S8 and Table S5). In the OF, the inflammation/immunity-related gene sets were profoundly upregulated in response to training AE, while gene sets related to nuclear glucocorticoid receptor binding, keratin filament, and intermediate filament were downregulated (Supplementary Figure S10 and Table S5).
Gene sets in relation to aerobic respiration, mitochondrial complex, and electron transport complex were upregulated in response to PI in the older people (Supplementary Figure S8 and S10, Table S5). Notably, gene sets in relation to the immune/inflammation process, collagen fibril organization, and ECM remodeling were significantly downregulated in response to PI (Supplementary Figure S8 and Table S5), which were upregulated in response to AE.
Contrary to AE alone, gene sets associated with the immune/inflammation process, ECM remodeling, cellular response, muscle organ development, renal system development, and regulation of vasculature development were downregulated in response to acute AE combined with PI, which were opposite to their responses to AE alone (Supplementary Table S5). No gene set was significantly enriched in response to training AE combined with PI (p > 0.05, FDR > 25%, Supplementary Table S5).
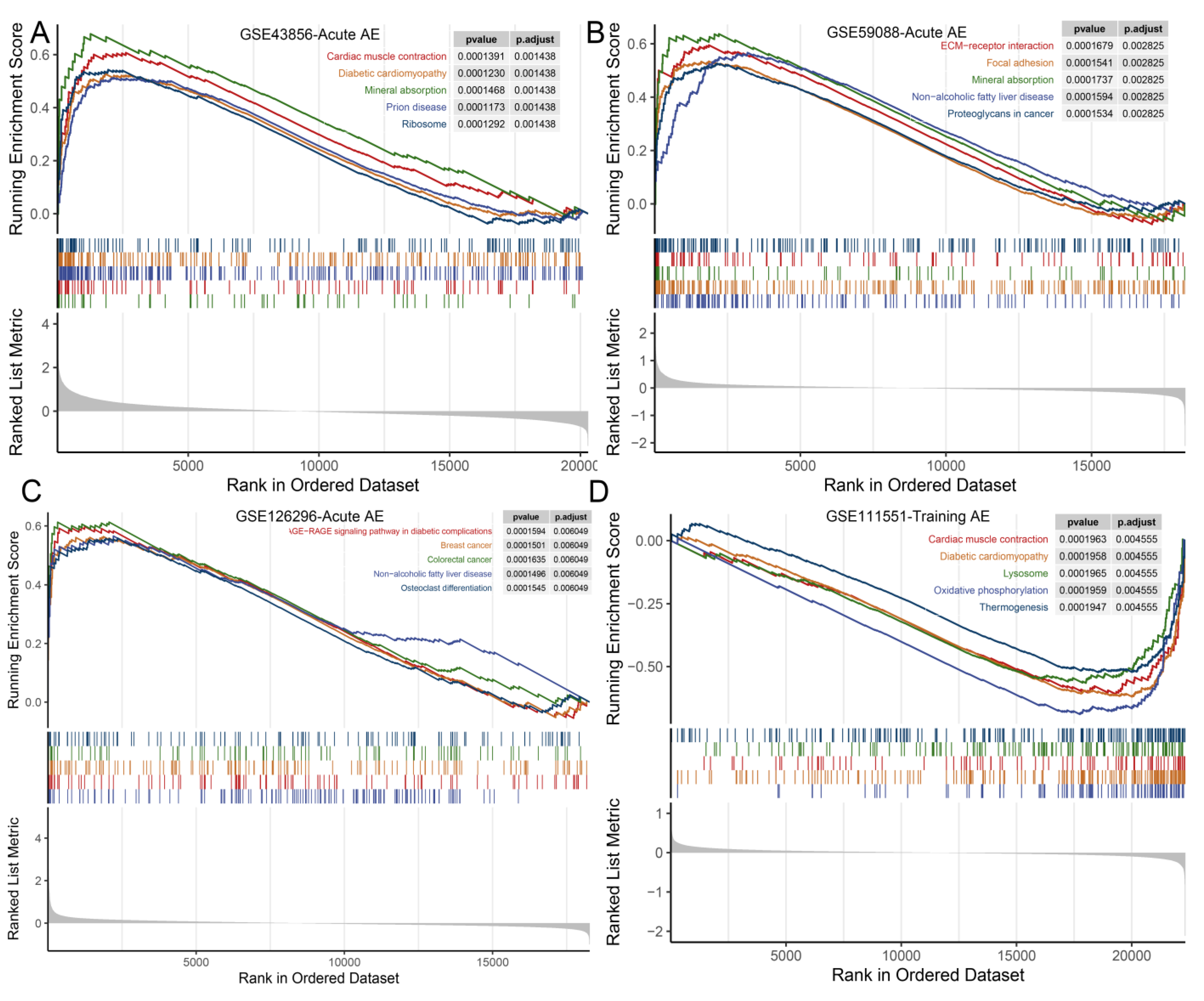
KEGG gene sets’ enrichment analysis revealed that the oxidative phosphorylation pathway was consistently upregulated both in response to acute AE and PI (Supplementary Table S6). Pathways of mineral absorption, non-alcoholic fatty liver disease, ECM receptor interaction, immune-related signaling pathways, and cardiac muscle contraction were upregulated in response to acute AE (Figure 5 and Figure S11, Supplementary Table S6). The Fanconi-anemia-pathway-, spliceosome-, and Herpes-simplex-virus-1-infection-related gene sets were downregulated in response to acute AE in the YM, and the Wnt signaling pathway and long-term potentiation were significantly downregulated in the OM (Supplementary Table S6). Oxidative phosphorylation, diabetic cardiomyopathy, and cardiac muscle contraction gene sets were downregulated in response to training AE in the YM (Figure 5D and Supplementary Table S6). ECM-receptor interaction and malaria pathways were upregulated in response to training AE in the older people (Supplementary Figures S12 and S13, Table S6). The neuroactive ligand–receptor interaction pathway was upregulated, while no KEGG pathway-related gene set was significantly enriched in response to PI in the YM (Supplementary Figure S11).
Gene sets concerning the olfactory transduction pathway were upregulated both in response to PI and acute AE combined with PI (Supplementary Table S6). Gene sets in the pathway of antigen processing and presentation, staphylococcus aureus infection, and phagosome were downregulated in response to PI in the OM (Supplementary Table S6). Herpes simplex virus 1 infection, systemic lupus erythematosus, and phagosome were downregulated in response to PI in the OF (Supplementary Figure S13). Protein-digestion-and-absorption-, systemic-lupus-erythematosus-, and TNF-signaling-pathway-related gene sets were downregulated in response to acute AE combined with PI (Supplementary Figure S11). Contrary to the response to acute AE, gene sets in AGE-RAGE signaling pathway (diabetic complications and cardiac muscle contraction) and ECM-receptor interaction pathway were downregulated to acute AE combined with PI (Supplementary Table S6). Additionally, no gene set in KEGG pathway was significantly enriched in response to training AE combined with PI (p > 0.05, FDR > 25%, Supplementary Figure S11 and Table S6).
4. Discussion
The transcriptional profiles of human skeletal muscle in response to AE, PI, and AE combined with PI were analyzed in the present study. ACTB, IL6, UBB, COL1A1, EZH2, DDIT3, MYC was the hub gene in response to acute AE in the YM and the OM, training AE in the YM and the OF, PI in the YM, acute AE combined with PI in the YM, and training AE combined with PI in the YM, respectively. FOS was upregulated in response to acute AE but was downregulated in response to PI and acute AE combined with PI. MYC was upregulated in response to acute AE while it was downregulated in response to AE combined with PI. COL1A1 was upregulated in response to training AE while it was downregulated in response to PI.
AE and PI may deliver impacts on human skeletal muscle in several analogous pathways, which included aerobic respiration, mitochondrial complexes, ECM remodeling, energy metabolic process, and immune/inflammatory responses. However, compared to PI, AE have an opposite impact on ECM remodeling and immune/inflammation process. Compared to PI, acute AE combined with PI may have additional impact on upregulating protein turnover and synthesis and downregulating musculoskeletal movement and muscle contraction. Few gene set was enriched in response to training AE combined with PI, which suggested that the limited observed biological impact on skeletal muscle induced by additional PI based on training AE.
4.1. Transcriptomic Changes in Skeletal Muscle in Response to Acute AE, PI, and Acute AE Combined with PI
The shared biological pathways and the overlap genes identified to acute AE and PI implied the molecular responses of muscle to them were not separate. The upregulated gene sets with both acute AE and PI alone were related to ATP synthesis and energy metabolic. AE could improve body metabolism by optimizing global energy expenditure and activating multiple neuroendocrine pathways [63], while more energy is needed to maintain skeletal muscle contraction due to rapid increase in mitochondrial oxidative enzymes during AE [64]. Meanwhile, high-protein diets could induce higher energy expenditure to regulate body metabolic status [65]. In the present study, mitochondrial-complex-related gene sets were consistently upregulated in acute AE, PI, and acute AE combined with PI, which suggested that both acute AE and PI may promote the aerobic respiration function. Moreover, AE could remodel the quantity and quality (function per unit) of skeletal muscle mitochondria to promote substrate oxidation and increase mitochondrial oxidative enzymes [64], while dietary protein-derived amino acid could incorporate into mitochondrial protein to promote mitochondrial content [45]. These may partly explain the feeble impact on mitochondrial content induced by acute AE, PI, and acute AE combined with PI.
The downregulation of immune/inflammation molecular process was identified both in response to PI and acute AE combined with PI in the present study, which was consistent with the reports in the previous RCTs [57,66]. Although inflammation induced by AE plays an important role in the increase in skeletal muscle mass and fiber components by skeletal muscle releasing cytokines, prostaglandins, and chemokines [67,68], PI may weaken the inflammation response. For example, the anti-oxidative effect of dietary whey protein may counteract chronic inflammation through regulation of the nuclear transcription factor kB signaling pathway and cytokine production [69]. ECM remodeling is also influenced by mesenchymal cells, and is closely related to the immune/inflammation process [70]. Reasonably, the response of ECM remodeling-related genes (sets) across acute AE, PI, and acute AE combined with PI was similar to the response of immune/inflammation in the present study.
The transcriptomic profile in response to acute AE combined with PI was comparable to PI alone, which may be due to the fact that the impact induced by acute AE had been offset in the combined intervention studies, where acute AE was always assigned to the control group. Meanwhile, the downregulation of skeletal muscle contraction, musculoskeletal movement, and muscle system development were found in response to acute AE combined with PI, which was contrary to the responses to AE alone. Thus, the additional impact may be induced by the combination of PI and AE. Postexercise PI, which may provide the most efficient gains in muscle plasticity and support myofibrillar protein synthesis as key components in defining contractile performance in response to AE [71]. Protein synthesis and turnover were upregulated both in response to acute AE and the combination of acute AE and PI but not in response to PI alone, which indicated that acute AE had a strong effect on protein synthesis and turnover and this effect may be enhanced when PI was added. This is reasonable in that muscle contraction during exercise, whether RE or EE, has a profound effect on muscle protein turnover [72]. These findings were also consistent with the reports from previous RCTs [73,74], which indicated that PI alone could not modulate myofibrillar protein synthesis (MyoPS) rates that were caused by disuse, while pre-sleep protein ingestion could stimulate myofibrillar and mitochondrial protein synthesis during an overnight recovery from AE. Protein ingestion may facilitate skeletal muscle conditioning following endurance training and may help athletes to improve endurance training efficiency [74]. Moreover, the type, dose, and time of PI may also contribute to the difference [75]. Protein supplementation studies suggested that a relatively higher protein intake was required to maximally stimulate skeletal muscle protein synthesis in the older people [76,77]. Greater PI intake during recovery from AE was important in the regulation of whole-body and muscle protein synthesis rates [45]. In addition, compared to plant protein, animal protein was more likely to be a stimulator of muscle protein synthesis [78].
ACTB are highly conserved proteins that are involved in cell motility, structure, integrity, and intercellular signaling, which helps organize and maintain the cellular morphology by virtue of facilitating the processes of migration, division, growth, signaling and shaping the cytoskeleton [79]. Meanwhile, previous studies found that ACTB might participate in the mechanism of vascular remodeling and thrombogenesis [80]. As the hub gene in response to acute AE in the OM, IL6 encodes a cytokine that functions in inflammation, maturation of B cells, tissue regeneration, and metabolism, which was consistent with the upregulation of the inflammation/immunity process in response to acute AE. FOS plays an important role in the gene interaction network of muscle in response to acute AE both in the YM and the OM, and it was the gene which was most upregulated in response to acute AE but was downregulated in response to PI and acute AE combined with PI. As a regulator of cell proliferation, differentiation, and transformation, FOS was involved in multiple inflammation pathways [81], which was also suggested by the results of GSEA in the present study. As the hub gene identified in response to acute AE combined with PI in the YM, DDIT3 is implicated in adipogenesis and erythropoiesis, is activated by endoplasmic reticulum stress, promotes apoptosis, and plays a crucial role in endoplasmic reticulum stress-induced apoptosis and autophagy, host innate immunity repression, and viral infection facilitation [82].
4.2. Transcriptomic Changes in Skeletal Muscle in Response to Training AE, PI, and Training AE Combined with PI
Different from the response to acute AE, a cluster of ATP-metabolic-processes-, oxidative-phosphorylation-, aerobic-respiratory-, fatty-acid-oxidation-and-transport-, and mitochondrion-related complexes were downregulated in response to training AE in the YM but not in the elderly. The training volume may critically affect the changes in mitochondrial content, whereas exercise intensity may play a more important role in the changes of mitochondrial function [83]. Training AE contributes to profound adaptions of the cardiorespiratory system that enhance the delivery of oxygen from the atmosphere to the mitochondria and enable a tighter regulation of muscle metabolism [84]. Hence, compared to acute AE, a period of regular endurance training may have already reserved a wider adaptation in the function of metabolism and mitochondria. Notably, both acute AE and training AE were related to promoted skeletal muscle development, inflammation/immunity responses, and ECM remodeling.
Inconsistent findings were reported for the response of protein turnover and protein synthesis rate to AE combined with PI [9,37,45,85]. The types of protein synthesis influenced by varied modes of exercise and period of interventions may help explain the inconsistences [37]. Acute AE, but not training AE or its combination with PI, may have a more positive effect on regulating protein folding/refolding and contribute more in protein quality control, protein turnover, and protein synthesis [86]. Additionally, compared to training AE combined with PI, acute AE combined with PI may have a more obvious impact on reducing immune/inflammation response, ECM remodeling, the positive regulation of receptor signaling pathway via JAK-STAT and STAT, cellular response, and renal system vasculature development and protein turnover.
UBB encodes ubiquitin, which is involved in targeting cellular proteins for degradation by the 26S proteosome and the maintenance of chromatin structure, the regulation of gene expression, and the stress response. Previous studies suggested AE training prevents oxidative stress and ubiquitin-proteasome system overactivity and reverse skeletal muscle atrophy [87]. Exercises could counter these fibrotic changes by stimulating the activity of genes associated with ECM remodeling like COL1A1, which was related to ECM collagen synthesis [88]. As the hub gene of muscles, in response to training AE combined with PI, MYC was significantly upregulated in response to acute AE in the younger male but downregulated in AE combined with PI, which has a widespread impact on the transcriptome including regulation of cell growth arrest and adhesion, metabolism, ribosome biogenesis, protein synthesis, and mitochondrial function [89]. Moreover, MYC was suggested to be associated with insulin sensitivity [90], which might play a vital role in the muscle metabolism response to exercise and PI.
4.3. Strengths and Limitations
The strength of the present study was that this is the first study to compare the transcriptomic profiles of human skeletal muscle in response to AE, PI, and AE combined with PI. The findings provided the molecular evidence to support the positive role of PI in addition to acute AE, but limited molecular evidence was found for training AE combined with PI. This would help provide molecular hints for developing optimal measures to promote muscle health. The present study had some limitations. Firstly, there were heterogeneities among the five groups of intervention, such as the type and duration of AE, the dose and content of PI, which may had a profound impact on the transcriptomic changes and bias the comparisons. Secondly, demographic characteristics were not adjusted for in the comparison across varied interventions. Moreover, no blank control group was set in the studies of the intervention of AE combined with PI, which limited the power to assess the interaction effect of AE and PI.
5. Conclusions
The present study suggested that human skeletal muscle may respond to AE and PI in analogous molecular pathways but the responses may differ in direction. PI and additional PI based on acute AE may attenuate the immune/inflammation response and ECM remodeling which would be promoted by AE, irrespective of the type of AE, age and gender. Compared to PI alone, acute AE combined with PI would further promote protein turnover and synthesis, but may suppress skeletal muscle contraction and movement. The molecular impact on human skeletal muscle delivered by training AE combined with PI was not significant. Further studies exploring on the optimal dose of PI in combination with AE for synergic effects are needed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/nu15153485/s1. Table S1: The summarization of top 5 up- and down-regulated DEGs, co-DEGs ranking by the absolute value of log2FC and gene expression network of skeletal muscle in response to different interventions within each dataset; Table S2: The hub gene screening from co-DEGs calculated by Cytohubba plugin in Cytoscape software (ranked by Degree value); Table S3: The GO enrichment analysis of co-DEGs from varied interventions in different populations; Table S4: The KEGG enrichment analysis of co-DEGs from varied interventions in different populations; Table S5: The gene sets related to GO terms from GSEA in each dataset by different interventions; Table S6: The gene sets related to KEGG pathways from GSEA in each dataset by different interventions. Figure S1: Flowchart of the screening of GEO datasets; Figure S2: Volcano plots across interventions for each dataset. The significantly upregulated and downregulated DEGs at the first 10 ranks were noted; Figure S3: The expression levels of FOS in each dataset by different interventions; Figure S4: The expression levels of MYC in each dataset by different interventions; Figure S5: The expression levels of COL1A1 in each dataset by different interventions; Figure S6: The GO enrichment analysis of co-DEGs from varied interventions in the older people; Figure S7: The KEGG enrichment analysis of co-DEGs from varied interventions in the older people; Figure S8: The gene sets related to GO terms from GSEA in each dataset by different interventions in the older male; Figure S9: The gene sets related to GO terms from GSEA in each dataset by varied interventions in the younger male; Figure S10: The gene sets related to GO terms from GSEA in each dataset by different interventions in the older female; Figure S11: Top 5 KEGG-pathway-related gene sets from GSEA in each dataset by different interventions in the younger male; Figure S12: Top 5 KEGG-pathway-related gene sets from GSEA-in each dataset by different interventions in the older male; Figure S13: The top 5 KEGG-pathway-related gene sets from GSEA in each dataset by different interventions in the older female.
Author Contributions
Conceptualization, Y.S.; methodology, Y.S. and X.Z.; software, X.Z.; validation, Y.S. and Z.X.; formal analysis, X.Z.; resources, Y.S. and X.Z.; data curation, X.Z. and L.L.; writing—original draft preparation, X.Z.; writing—review and editing, Y.S., L.Z., and T.K.; visualization, X.Z.; supervision, Y.S. and T.K.; project administration, Y.S.; funding acquisition, Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China [No. 82003541], the Science-Technology Foundation for Young Scientists of Hunan Province of China [No. 2022RC1207], the Research Foundation of Education Bureau of Hunan Province [No. 20B367], and the Natural Science Foundation of Hunan Province of China [No. 2021JJ40371].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The summary-level datasets used and/or analyzed in the current study are available from the corresponding authors on reasonable request.
Acknowledgments
Thanks to all the original studies used for the analysis, which are publicly available on the GEO repository. Thanks to all the investigators and participants who contributed to those studies.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wolfe, R.R. The underappreciated role of muscle in health and disease. Am. J. Clin. Nutr. 2006, 84, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.; Morley, J.E.; Schols, A.; Ferrucci, L.; Cruz-Jentoft, A.J.; Dent, E.; Baracos, V.E.; Crawford, J.A.; Doehner, W.; Heymsfield, S.B.; et al. Sarcopenia: A Time for Action. An SCWD Position Paper. J. Cachexia Sarcopenia Muscle 2019, 10, 956–961. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Landi, F.; Schneider, S.M.; Zuniga, C.; Arai, H.; Boirie, Y.; Chen, L.K.; Fielding, R.A.; Martin, F.C.; Michel, J.P.; et al. Prevalence of and interventions for sarcopenia in ageing adults: A systematic review. Report of the International Sarcopenia Initiative (EWGSOP and IWGS). Age Ageing 2014, 43, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef]
- Vikberg, S.; Sorlen, N.; Branden, L.; Johansson, J.; Nordstrom, A.; Hult, A.; Nordstrom, P. Effects of Resistance Training on Functional Strength and Muscle Mass in 70-Year-Old Individuals with Pre-sarcopenia: A Randomized Controlled Trial. J. Am. Med. Dir. Assoc. 2019, 20, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Kemmler, W.; Kohl, M.; Frohlich, M.; Jakob, F.; Engelke, K.; von Stengel, S.; Schoene, D. Effects of High-Intensity Resistance Training on Osteopenia and Sarcopenia Parameters in Older Men with Osteosarcopenia-One-Year Results of the Randomized Controlled Franconian Osteopenia and Sarcopenia Trial (FrOST). J. Bone Miner. Res. 2020, 35, 1634–1644. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, Y.; Yamada, Y.; Maeda, A.; Izumo, T.; Rogi, T.; Shibata, H.; Fukuda, M.; Arimitsu, T.; Miyamoto, N.; Hashimoto, T. Effects of resistance training intensity on muscle quantity/quality in middle-aged and older people: A randomized controlled trial. J. Cachexia Sarcopenia Muscle 2022, 13, 894–908. [Google Scholar] [CrossRef]
- Hartono, F.A.; Martin-Arrowsmith, P.W.; Peeters, W.M.; Churchward-Venne, T.A. The Effects of Dietary Protein Supplementation on Acute Changes in Muscle Protein Synthesis and Longer-Term Changes in Muscle Mass, Strength, and Aerobic Capacity in Response to Concurrent Resistance and Endurance Exercise in Healthy Adults: A Systematic Review. Sports Med. 2022, 52, 1295–1328. [Google Scholar]
- Fyfe, J.J.; Bishop, D.J.; Stepto, N.K. Interference between concurrent resistance and endurance exercise: Molecular bases and the role of individual training variables. Sports Med. 2014, 44, 743–762. [Google Scholar] [CrossRef]
- Hurst, C.; Robinson, S.M.; Witham, M.D.; Dodds, R.M.; Granic, A.; Buckland, C.; De Biase, S.; Finnegan, S.; Rochester, L.; Skelton, D.A.; et al. Resistance exercise as a treatment for sarcopenia: Prescription and delivery. Age Ageing 2022, 51, afac003. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Befroy, D.; Dufour, S.; Dziura, J.; Ariyan, C.; Rothman, D.L.; DiPietro, L.; Cline, G.W.; Shulman, G.I. Mitochondrial dysfunction in the elderly: Possible role in insulin resistance. Science 2003, 300, 1140–1142. [Google Scholar] [CrossRef] [Green Version]
- Short, K.R.; Bigelow, M.L.; Kahl, J.; Singh, R.; Coenen-Schimke, J.; Raghavakaimal, S.; Nair, K.S. Decline in skeletal muscle mitochondrial function with aging in humans. Proc. Natl. Acad. Sci. USA 2005, 102, 5618–5623. [Google Scholar] [CrossRef] [PubMed]
- Mitsuishi, M.; Miyashita, K.; Muraki, A.; Tamaki, M.; Tanaka, K.; Itoh, H. Dietary protein decreases exercise endurance through rapamycin-sensitive suppression of muscle mitochondria. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E776–E784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leduc-Gaudet, J.P.; Hussain, S.N.A.; Barreiro, E.; Gouspillou, G. Mitochondrial Dynamics and Mitophagy in Skeletal Muscle Health and Aging. Int. J. Mol. Sci. 2021, 22, 8179. [Google Scholar] [CrossRef]
- Petersen, K.F.; Dufour, S.; Befroy, D.; Garcia, R.; Shulman, G.I. Impaired mitochondrial activity in the insulin-resistant offspring of patients with type 2 diabetes. N. Engl. J. Med. 2004, 350, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Curcio, F.; Testa, G.; Liguori, I.; Papillo, M.; Flocco, V.; Panicara, V.; Galizia, G.; Della-Morte, D.; Gargiulo, G.; Cacciatore, F.; et al. Sarcopenia and Heart Failure. Nutrients 2020, 12, 211. [Google Scholar] [CrossRef] [Green Version]
- Lavine, K.J.; Sierra, O.L. Skeletal muscle inflammation and atrophy in heart failure. Heart Fail. Rev. 2017, 22, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Kalinkovich, A.; Livshits, G. Sarcopenic obesity or obese sarcopenia: A cross talk between age-associated adipose tissue and skeletal muscle inflammation as a main mechanism of the pathogenesis. Ageing Res. Rev. 2017, 35, 200–221. [Google Scholar] [CrossRef]
- Petridou, A.; Siopi, A.; Mougios, V. Exercise in the management of obesity. Metabolism 2019, 92, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Ismail, I.; Keating, S.E.; Baker, M.K.; Johnson, N.A. A systematic review and meta-analysis of the effect of aerobic vs. resistance exercise training on visceral fat. Obes. Rev. 2012, 13, 68–91. [Google Scholar] [CrossRef] [PubMed]
- Beckers, P.J.; Denollet, J.; Possemiers, N.M.; Wuyts, F.L.; Vrints, C.J.; Conraads, V.M. Combined endurance-resistance training vs. endurance training in patients with chronic heart failure: A prospective randomized study. Eur. Heart J. 2008, 29, 1858–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Kitzman, D.W.; Brubaker, P.; Haykowsky, M.J.; Morgan, T.; Becton, J.T.; Berry, J.D. Response to Endurance Exercise Training in Older Adults with Heart Failure with Preserved or Reduced Ejection Fraction. J. Am. Geriatr. Soc. 2017, 65, 1698–1704. [Google Scholar] [CrossRef]
- Joisten, N.; Kummerhoff, F.; Koliamitra, C.; Schenk, A.; Walzik, D.; Hardt, L.; Knoop, A.; Thevis, M.; Kiesl, D.; Metcalfe, A.J.; et al. Exercise and the Kynurenine pathway: Current state of knowledge and results from a randomized cross-over study comparing acute effects of endurance and resistance training. Exerc. Immunol. Rev. 2020, 26, 24–42. [Google Scholar] [PubMed]
- Schlagheck, M.L.; Walzik, D.; Joisten, N.; Koliamitra, C.; Hardt, L.; Metcalfe, A.J.; Wahl, P.; Bloch, W.; Schenk, A.; Zimmer, P. Cellular immune response to acute exercise: Comparison of endurance and resistance exercise. Eur. J. Haematol. 2020, 105, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Mao, L.; Feng, Y.; Ainsworth, B.E.; Liu, Y.; Chen, N. Effects of different exercise training modes on muscle strength and physical performance in older people with sarcopenia: A systematic review and meta-analysis. BMC Geriatr. 2021, 21, 708. [Google Scholar] [CrossRef]
- Strasser, B.; Keinrad, M.; Haber, P.; Schobersberger, W. Efficacy of systematic endurance and resistance training on muscle strength and endurance performance in elderly adults—A randomized controlled trial. Wien. Klin. Wochenschr. 2009, 121, 757–764. [Google Scholar] [CrossRef]
- Rondanelli, M.; Cereda, E.; Klersy, C.; Faliva, M.A.; Peroni, G.; Nichetti, M.; Gasparri, C.; Iannello, G.; Spadaccini, D.; Infantino, V.; et al. Improving rehabilitation in sarcopenia: A randomized-controlled trial utilizing a muscle-targeted food for special medical purposes. J. Cachexia Sarcopenia Muscle 2020, 11, 1535–1547. [Google Scholar] [CrossRef]
- Park, S.J.; Park, J.; Won, C.W.; Lee, H.J. The Inverse Association of Sarcopenia and Protein-Source Food and Vegetable Intakes in the Korean Elderly: The Korean Frailty and Aging Cohort Study. Nutrients 2022, 14, 1375. [Google Scholar] [CrossRef]
- Verlaan, S.; Maier, A.B.; Bauer, J.M.; Bautmans, I.; Brandt, K.; Donini, L.M.; Maggio, M.; McMurdo, M.E.T.; Mets, T.; Seal, C.; et al. Sufficient levels of 25-hydroxyvitamin D and protein intake required to increase muscle mass in sarcopenic older adults—The PROVIDE study. Clin. Nutr. 2018, 37, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Nunes, E.A.; Colenso-Semple, L.; McKellar, S.R.; Yau, T.; Ali, M.U.; Fitzpatrick-Lewis, D.; Sherifali, D.; Gaudichon, C.; Tome, D.; Atherton, P.J.; et al. Systematic review and meta-analysis of protein intake to support muscle mass and function in healthy adults. J Cachexia Sarcopenia Muscle 2022, 13, 795–810. [Google Scholar] [CrossRef]
- Park, Y.; Choi, J.E.; Hwang, H.S. Protein supplementation improves muscle mass and physical performance in undernourished prefrail and frail elderly subjects: A randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2018, 108, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.Y.; Huang, K.S.; Chen, K.M.; Chou, C.P.; Tu, Y.K. Exercise, Nutrition, and Combined Exercise and Nutrition in Older Adults with Sarcopenia: A Systematic Review and Network Meta-analysis. Maturitas 2021, 145, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Deldicque, L. Protein Intake and Exercise-Induced Skeletal Muscle Hypertrophy: An Update. Nutrients 2020, 12, 2023. [Google Scholar] [CrossRef]
- Longland, T.M.; Oikawa, S.Y.; Mitchell, C.J.; Devries, M.C.; Phillips, S.M. Higher compared with lower dietary protein during an energy deficit combined with intense exercise promotes greater lean mass gain and fat mass loss: A randomized trial. Am. J. Clin. Nutr. 2016, 103, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Kemmler, W.; Kohl, M.; Jakob, F.; Engelke, K.; von Stengel, S. Effects of High Intensity Dynamic Resistance Exercise and Whey Protein Supplements on Osteosarcopenia in Older Men with Low Bone and Muscle Mass. Final Results of the Randomized Controlled FrOST Study. Nutrients 2020, 12, 2341. [Google Scholar] [CrossRef] [PubMed]
- Breen, L.; Philp, A.; Witard, O.C.; Jackman, S.R.; Selby, A.; Smith, K.; Baar, K.; Tipton, K.D. The influence of carbohydrate-protein co-ingestion following endurance exercise on myofibrillar and mitochondrial protein synthesis. J. Physiol. 2011, 589, 4011–4025. [Google Scholar] [CrossRef]
- Hawley, J.A.; Burke, L.M.; Phillips, S.M.; Spriet, L.L. Nutritional modulation of training-induced skeletal muscle adaptations. J Appl. Physiol. 2011, 110, 834–845. [Google Scholar] [CrossRef] [Green Version]
- Pillon, N.J.; Gabriel, B.M.; Dollet, L.; Smith, J.A.B.; Sardon Puig, L.; Botella, J.; Bishop, D.J.; Krook, A.; Zierath, J.R. Transcriptomic profiling of skeletal muscle adaptations to exercise and inactivity. Nat. Commun. 2020, 11, 470. [Google Scholar] [CrossRef] [Green Version]
- Deane, C.S.; Willis, C.R.G.; Phillips, B.E.; Atherton, P.J.; Harries, L.W.; Ames, R.M.; Szewczyk, N.J.; Etheridge, T. Transcriptomic meta-analysis of disuse muscle atrophy vs. resistance exercise-induced hypertrophy in young and older humans. J. Cachexia Sarcopenia Muscle 2021, 12, 629–645. [Google Scholar] [CrossRef]
- Amar, D.; Lindholm, M.E.; Norrbom, J.; Wheeler, M.T.; Rivas, M.A.; Ashley, E.A. Time trajectories in the transcriptomic response to exercise-a meta-analysis. Nat. Commun. 2021, 12, 3471. [Google Scholar] [CrossRef] [PubMed]
- Knuiman, P.; van Loon, L.J.C.; Wouters, J.; Hopman, M.; Mensink, M. Protein supplementation elicits greater gains in maximal oxygen uptake capacity and stimulates lean mass accretion during prolonged endurance training: A double-blind randomized controlled trial. Am. J. Clin. Nutr. 2019, 110, 508–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonvik, K.L.; Paulussen, K.J.M.; Danen, S.L.; Ceelen, I.J.M.; Horstman, A.M.; Wardenaar, F.C.; VAN Loon, L.J.C.; VAN Dijk, J.-W. Protein Supplementation Does Not Augment Adaptations to Endurance Exercise Training. Med. Sci. Sports Exerc. 2019, 51, 2041–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, E.; Kato, H.; Volterman, K.A.; Suzuki, K.; Moore, D.R. The Effect of Dietary Protein on Protein Metabolism and Performance in Endurance-trained Males. Med. Sci. Sports Exerc. 2019, 51, 352–360. [Google Scholar] [CrossRef]
- Churchward-Venne, T.A.; Pinckaers, P.J.M.; Smeets, J.S.J.; Betz, M.W.; Senden, J.M.; Goessens, J.P.B.; Gijsen, A.P.; Rollo, I.; Verdijk, L.B.; van Loon, L.J.C. Dose-response effects of dietary protein on muscle protein synthesis during recovery from endurance exercise in young men: A double-blind randomized trial. Am. J. Clin. Nutr. 2020, 112, 303–317. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. S4), S11. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, O.; Sabapathy, S.; Lazarus, R.; Jowett, J.B.; Desbrow, B.; Peake, J.M.; Cameron-Smith, D.; Haseler, L.J.; Wagner, K.H.; Bulmer, A.C. Transcriptome analysis of neutrophils after endurance exercise reveals novel signaling mechanisms in the immune response to physiological stress. J. Appl. Physiol. 2013, 114, 1677–1688. [Google Scholar] [CrossRef] [Green Version]
- Vissing, K.; Schjerling, P. Simplified data access on human skeletal muscle transcriptome responses to differentiated exercise. Sci. Data 2014, 1, 140041. [Google Scholar] [CrossRef] [Green Version]
- Rundqvist, H.C.; Montelius, A.; Osterlund, T.; Norman, B.; Esbjornsson, M.; Jansson, E. Acute sprint exercise transcriptome in human skeletal muscle. PLoS ONE 2019, 14, e0223024. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.S.; Zhao, X.; Irmler, M.; Liu, X.; Hoene, M.; Scheler, M.; Li, Y.; Beckers, J.; Hrabe de Angelis, M.; Haring, H.U.; et al. Type 2 diabetes alters metabolic and transcriptional signatures of glucose and amino acid metabolism during exercise and recovery. Diabetologia 2015, 58, 1845–1854. [Google Scholar] [CrossRef] [Green Version]
- Böhm, A.; Hoffmann, C.; Irmler, M.; Schneeweiss, P.; Schnauder, G.; Sailer, C.; Schmid, V.; Hudemann, J.; Machann, J.; Schick, F.; et al. TGF-b Contributes to Impaired Exercise Response by Suppression of Mitochondrial Key Regulators in Skeletal Muscle. Diabetes 2016, 65, 2846–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radom-Aizik, S.; Hayek, S.; Shahar, I.; Rechavi, G.; Kaminski, N.; Ben-Dov, I. Effects of aerobic training on gene expression in skeletal muscle of elderly men. Med. Sci. Sports Exerc. 2005, 37, 1680–1696. [Google Scholar] [CrossRef]
- Poelkens, F.; Lammers, G.; Pardoel, E.M.; Tack, C.J.; Hopman, M.T. Upregulation of skeletal muscle inflammatory genes links inflammation with insulin resistance in women with the metabolic syndrome. Exp. Physiol. 2013, 98, 1485–1494. [Google Scholar] [CrossRef]
- Thalacker-Mercer, A.E.; Fleet, J.C.; Craig, B.A.; Carnell, N.S.; Campbell, W.W. Inadequate protein intake affects skeletal muscle transcript profiles in older humans. Am. J. Clin. Nutr. 2007, 85, 1344–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thalacker-Mercer, A.E.; Fleet, J.C.; Craig, B.A.; Campbell, W.W. The skeletal muscle transcript profile reflects accommodative responses to inadequate protein intake in younger and older males. J. Nutr. Biochem. 2010, 21, 1076–1082. [Google Scholar] [CrossRef]
- Smith, G.I.; Yoshino, J.; Kelly, S.C.; Reeds, D.N.; Okunade, A.; Patterson, B.W.; Klein, S.; Mittendorfer, B. High-Protein Intake during Weight Loss Therapy Eliminates the Weight-Loss-Induced Improvement in Insulin Action in Obese Postmenopausal Women. Cell Rep. 2016, 17, 849–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowlands, D.S.; Thomson, J.S.; Timmons, B.W.; Raymond, F.; Fuerholz, A.; Mansourian, R.; Zwahlen, M.C.; Metairon, S.; Glover, E.; Stellingwerff, T.; et al. Transcriptome and translational signaling following endurance exercise in trained skeletal muscle: Impact of dietary protein. Physiol. Genom. 2011, 43, 1004–1020. [Google Scholar] [CrossRef] [Green Version]
- Rowlands, D.S.; Nelson, A.R.; Raymond, F.; Metairon, S.; Mansourian, R.; Clarke, J.; Stellingwerff, T.; Phillips, S.M. Protein-leucine ingestion activates a regenerative inflammo-myogenic transcriptome in skeletal muscle following intense endurance exercise. Physiol. Genom. 2016, 48, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knuiman, P.; Hangelbroek, R.; Boekschoten, M.; Hopman, M.; Mensink, M. Impact of protein supplementation during endurance training on changes in skeletal muscle transcriptome. BMC Genom. 2020, 21, 397. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, A.M.; Elizondo-Montemayor, L. The Role of Exercise in the Interplay between Myokines, Hepatokines, Osteokines, Adipokines, and Modulation of Inflammation for Energy Substrate Redistribution and Fat Mass Loss: A Review. Nutrients 2020, 12, 1899. [Google Scholar] [CrossRef]
- Murphy, R.M.; Watt, M.J.; Febbraio, M.A. Metabolic communication during exercise. Nat. Metab. 2020, 2, 805–816. [Google Scholar] [CrossRef]
- Smeets, A.J.; Soenen, S.; Luscombe-Marsh, N.D.; Ueland, O.; Westerterp-Plantenga, M.S. Energy expenditure, satiety, and plasma ghrelin, glucagon-like peptide 1, and peptide tyrosine-tyrosine concentrations following a single high-protein lunch. J. Nutr. 2008, 138, 698–702. [Google Scholar] [CrossRef] [Green Version]
- Gaffney, K.; Lucero, A.; Macartney-Coxson, D.; Clapham, J.; Whitfield, P.; Palmer, B.R.; Wakefield, S.; Faulkner, J.; Stoner, L.; Rowlands, D.S. Effects of whey protein on skeletal muscle microvascular and mitochondrial plasticity following 10 weeks of exercise training in men with type 2 diabetes. Appl. Physiol. Nutr. Metab. 2021, 46, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Neubauer, O.; Della Gatta, P.A.; Nosaka, K. Muscle damage and inflammation during recovery from exercise. J. Appl. Physiol. 2017, 122, 559–570. [Google Scholar] [CrossRef]
- Louis, E.; Raue, U.; Yang, Y.; Jemiolo, B.; Trappe, S. Time course of proteolytic, cytokine, and myostatin gene expression after acute exercise in human skeletal muscle. J. Appl. Physiol. 2007, 103, 1744–1751. [Google Scholar] [CrossRef] [Green Version]
- Draganidis, D.; Karagounis, L.G.; Athanailidis, I.; Chatzinikolaou, A.; Jamurtas, A.Z.; Fatouros, I.G. Inflammaging and Skeletal Muscle: Can Protein Intake Make a Difference? J. Nutr. 2016, 146, 1940–1952. [Google Scholar] [CrossRef] [Green Version]
- Koliaraki, V.; Prados, A.; Armaka, M.; Kollias, G. The mesenchymal context in inflammation, immunity and cancer. Nat. Immunol. 2020, 21, 974–982. [Google Scholar] [CrossRef]
- Rowlands, D.S.; Nelson, A.R.; Phillips, S.M.; Faulkner, J.A.; Clarke, J.; Burd, N.A.; Moore, D.; Stellingwerff, T. Protein-leucine fed dose effects on muscle protein synthesis after endurance exercise. Med. Sci. Sports Exerc. 2015, 47, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Burd, N.A.; Tang, J.E.; Moore, D.R.; Phillips, S.M. Exercise training and protein metabolism: Influences of contraction, protein intake, and sex-based differences. J. Appl. Physiol. 2009, 106, 1692–1701. [Google Scholar] [CrossRef]
- Kilroe, S.P.; Fulford, J.; Jackman, S.; Holwerda, A.; Gijsen, A.; van Loon, L.; Wall, B.T. Dietary protein intake does not modulate daily myofibrillar protein synthesis rates or loss of muscle mass and function during short-term immobilization in young men: A randomized controlled trial. Am. J. Clin. Nutr. 2021, 113, 548–561. [Google Scholar] [CrossRef] [PubMed]
- Trommelen, J.; van Lieshout, G.A.A.; Pabla, P.; Nyakayiru, J.; Hendriks, F.K.; Senden, J.M.; Goessens, J.P.B.; van Kranenburg, J.M.X.; Gijsen, A.P.; Verdijk, L.B.; et al. Pre-sleep Protein Ingestion Increases Mitochondrial Protein Synthesis Rates during Overnight Recovery from Endurance Exercise: A Randomized Controlled Trial. Sports Med. 2023, 53, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, J.; Tomita, T.; Arimitsu, T.; Fujita, S. Evenly Distributed Protein Intake over 3 Meals Augments Resistance Exercise-Induced Muscle Hypertrophy in Healthy Young Men. J. Nutr. 2020, 150, 1845–1851. [Google Scholar] [CrossRef] [Green Version]
- Dirks, M.L.; Wall, B.T.; Nilwik, R.; Weerts, D.H.; Verdijk, L.B.; van Loon, L.J. Skeletal muscle disuse atrophy is not attenuated by dietary protein supplementation in healthy older men. J. Nutr. 2014, 144, 1196–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzke, B.; Neubauer, O.; Cameron-Smith, D.; Wagner, K.H. Dietary Protein, Muscle and Physical Function in the Very Old. Nutrients 2018, 10, 935. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.T.; Pan, B.J.; Toh, D.W.K.; Sutanto, C.N.; Kim, J.E. Animal Protein versus Plant Protein in Supporting Lean Mass and Muscle Strength: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2021, 13, 661. [Google Scholar] [CrossRef]
- Chen, G.; Zou, Y.; Zhang, X.; Xu, L.; Hu, Q.; Li, T.; Yao, C.; Yu, S.; Wang, X.; Wang, C. Beta-Actin protein expression differs in the submandibular glands of male and female mice. Cell Biol. Int. 2016, 40, 779–786. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, Y.; Chen, X.; Lu, X.; Chen, Y.; Zhao, X.; Zhu, L.; Fang, Z.; Zhao, H.; Yao, Y.; et al. The ACTB Variants and Alcohol Drinking Confer Joint Effect to Ischemic Stroke in Chinese Han Population. J. Atheroscler. Thromb. 2020, 27, 226–244. [Google Scholar] [CrossRef]
- Ha, A.T.; Cho, J.Y.; Kim, D. MLK3 Regulates Inflammatory Response via Activation of AP-1 Pathway in HEK293 and RAW264.7 Cells. Int. J. Mol. Sci. 2022, 23, 10874. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hou, P.; Pan, W.; He, W.; He, D.C.; Wang, H.; He, H. DDIT3 Targets Innate Immunity via the DDIT3-OTUD1-MAVS Pathway to Promote Bovine Viral Diarrhea Virus Replication. J. Virol. 2021, 95, e02351-20. [Google Scholar] [CrossRef] [PubMed]
- Granata, C.; Jamnick, N.A.; Bishop, D.J. Training-Induced Changes in Mitochondrial Content and Respiratory Function in Human Skeletal Muscle. Sports Med. 2018, 48, 1809–1828. [Google Scholar] [CrossRef]
- Jones, A.M.; Carter, H. The effect of endurance training on parameters of aerobic fitness. Sports Med. 2000, 29, 373–386. [Google Scholar] [CrossRef]
- Rodriguez, N.R.; Vislocky, L.M.; Gaine, P.C. Dietary protein, endurance exercise, and human skeletal-muscle protein turnover. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Bozaykut, P.; Ozer, N.K.; Karademir, B. Regulation of protein turnover by heat shock proteins. Free Radic. Biol. Med. 2014, 77, 195–209. [Google Scholar] [CrossRef]
- Cunha, T.F.; Bacurau, A.V.; Moreira, J.B.; Paixao, N.A.; Campos, J.C.; Ferreira, J.C.; Leal, M.L.; Negrao, C.E.; Moriscot, A.S.; Wisloff, U.; et al. Exercise training prevents oxidative stress and ubiquitin-proteasome system overactivity and reverse skeletal muscle atrophy in heart failure. PLoS ONE 2012, 7, e41701. [Google Scholar] [CrossRef]
- Huertas, J.R.; Rodríguez Lara, A.; González Acevedo, O.; Mesa-García, M.D. Milk and dairy products as vehicle for calcium and vitamin D: Role of calcium enriched milks. Nutr. Hosp. 2019, 36, 962–973. [Google Scholar]
- Dang, C.V.; O’Donnell, K.A.; Zeller, K.I.; Nguyen, T.; Osthus, R.C.; Li, F. The c-Myc target gene network. Semin. Cancer Biol. 2006, 16, 253–264. [Google Scholar] [CrossRef]
- McLean, C.S.; Mielke, C.; Cordova, J.M.; Langlais, P.R.; Bowen, B.; Miranda, D.; Coletta, D.K.; Mandarino, L.J. Gene and MicroRNA Expression Responses to Exercise; Relationship with Insulin Sensitivity. PLoS ONE 2015, 10, e0127089. [Google Scholar] [CrossRef]
Figure 1.
(A,B) The number of identified DEGs in each dataset and co-DEGs from each intervention among different populations. Note: AE, aerobic exercise; PI, protein intake; Y, the younger one; O, the older one; M, male; F, female; YM, the younger male; OM, the older male; OF, the older female.
Figure 1.
(A,B) The number of identified DEGs in each dataset and co-DEGs from each intervention among different populations. Note: AE, aerobic exercise; PI, protein intake; Y, the younger one; O, the older one; M, male; F, female; YM, the younger male; OM, the older male; OF, the older female.
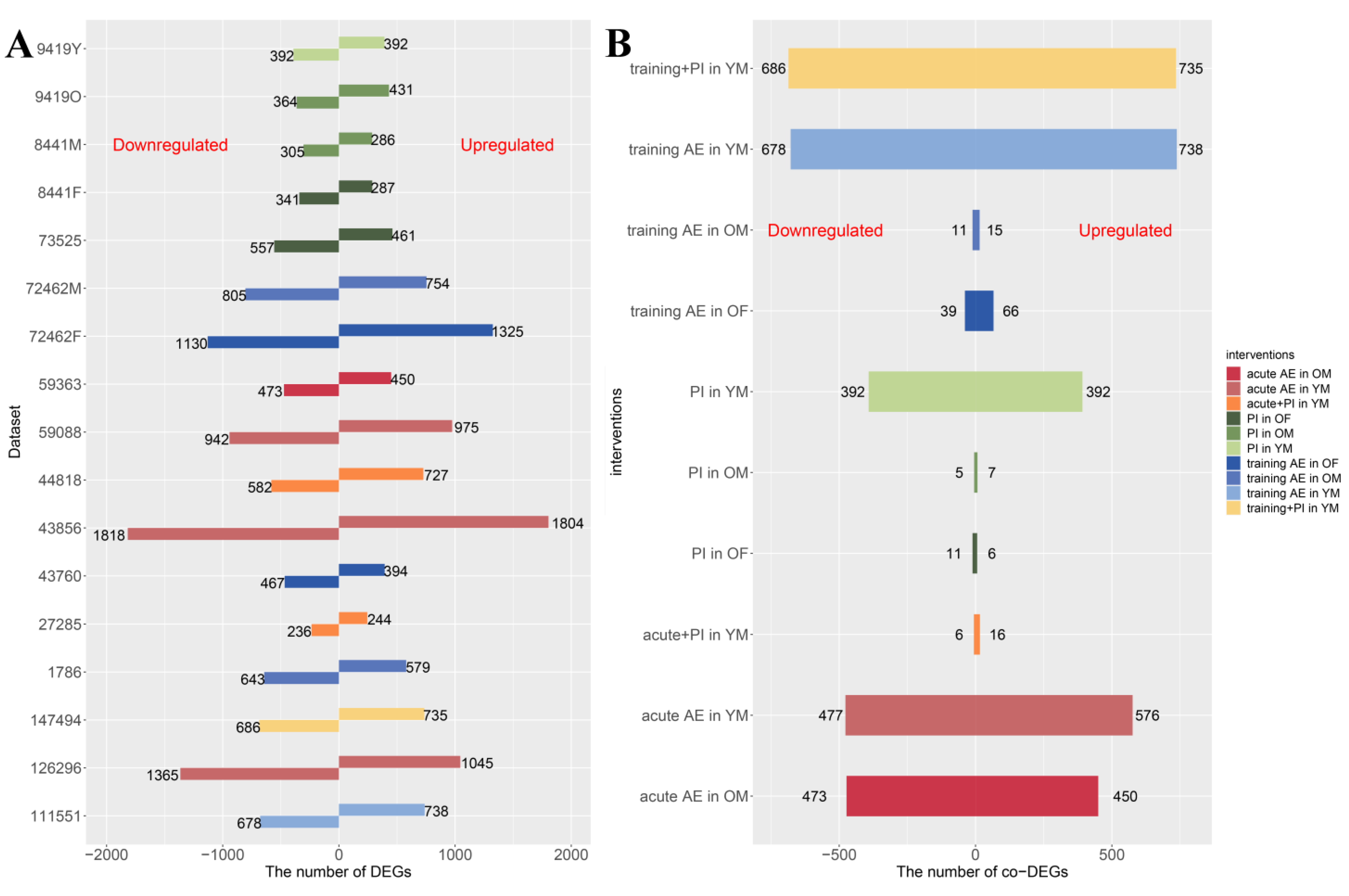
Figure 2.
The GO enrichment analysis of co-DEGs from varied interventions in the younger male. (A) acute AE; (B) training AE; (C) PI; (D) acute AE combined with PI; (E) training AE combined with PI. Note: GO, Gene Ontology; co-DEGs, co-differentially expressed genes; AE, aerobic exercise; PI, protein intake.
Figure 2.
The GO enrichment analysis of co-DEGs from varied interventions in the younger male. (A) acute AE; (B) training AE; (C) PI; (D) acute AE combined with PI; (E) training AE combined with PI. Note: GO, Gene Ontology; co-DEGs, co-differentially expressed genes; AE, aerobic exercise; PI, protein intake.

Figure 3.
The KEGG enrichment analysis of co-DEGs from varied interventions in the younger male. (A) acute AE; (B) training AE; (C) PI; (D) acute AE combined with PI; (E) training AE combined with PI. Note: KEGG, Kyoto Encyclopedia of Genes and Genomes; co-DEGs, co-differentially expressed genes; AE, aerobic exercise; PI, protein intake.
Figure 3.
The KEGG enrichment analysis of co-DEGs from varied interventions in the younger male. (A) acute AE; (B) training AE; (C) PI; (D) acute AE combined with PI; (E) training AE combined with PI. Note: KEGG, Kyoto Encyclopedia of Genes and Genomes; co-DEGs, co-differentially expressed genes; AE, aerobic exercise; PI, protein intake.
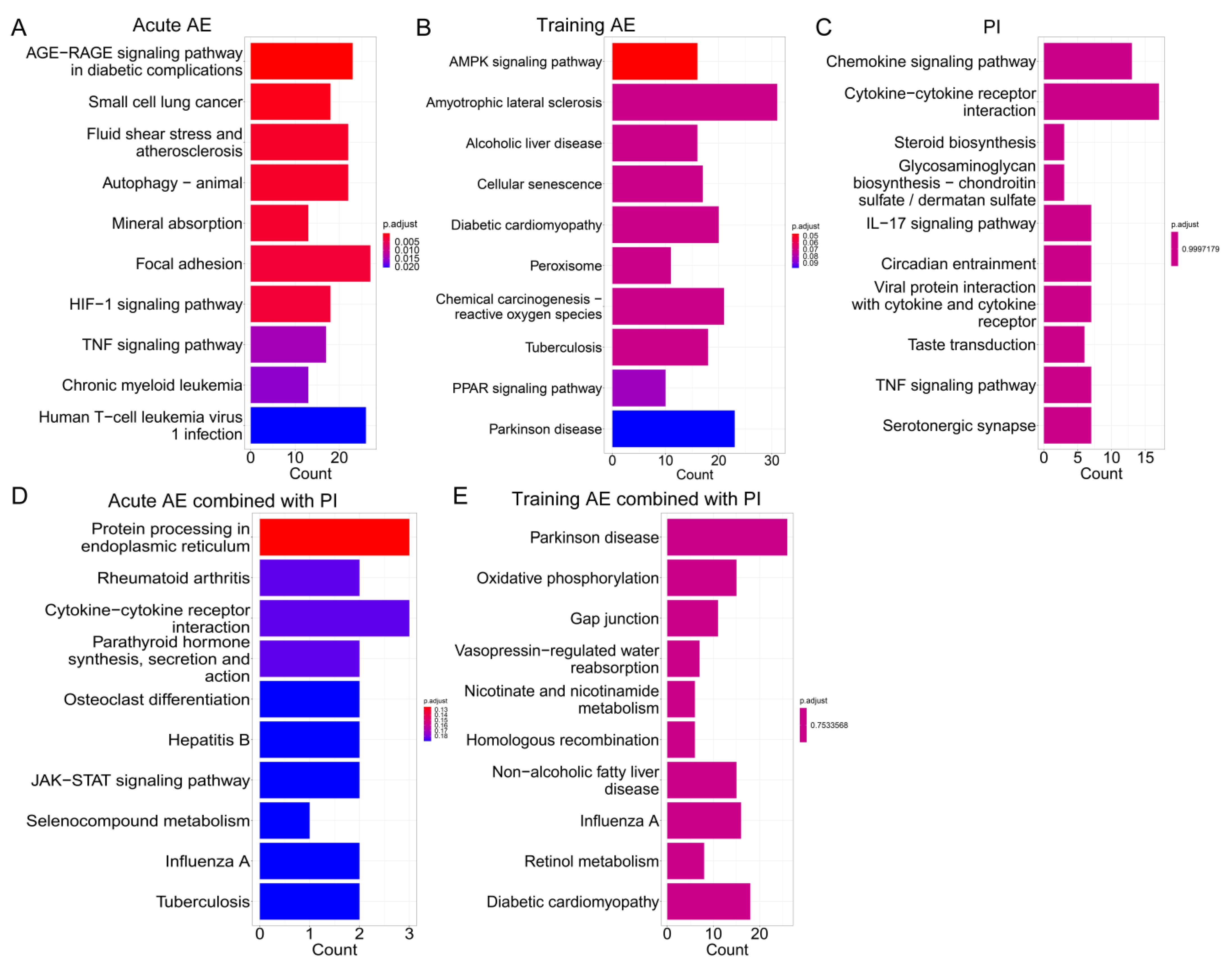
Figure 4.
The gene sets related GO terms from GSEA in each dataset of AE in the younger male. (A) GSE43856—Acute AE; (B) GSE59088—acute AE; (C) GSE126296—acute AE; (D) GSE111551—training AE. Note: GO, Gene Ontology; GSEA, gene set enrichment analysis; AE, aerobic exercise; NES, normalized enrichment score.
Figure 4.
The gene sets related GO terms from GSEA in each dataset of AE in the younger male. (A) GSE43856—Acute AE; (B) GSE59088—acute AE; (C) GSE126296—acute AE; (D) GSE111551—training AE. Note: GO, Gene Ontology; GSEA, gene set enrichment analysis; AE, aerobic exercise; NES, normalized enrichment score.
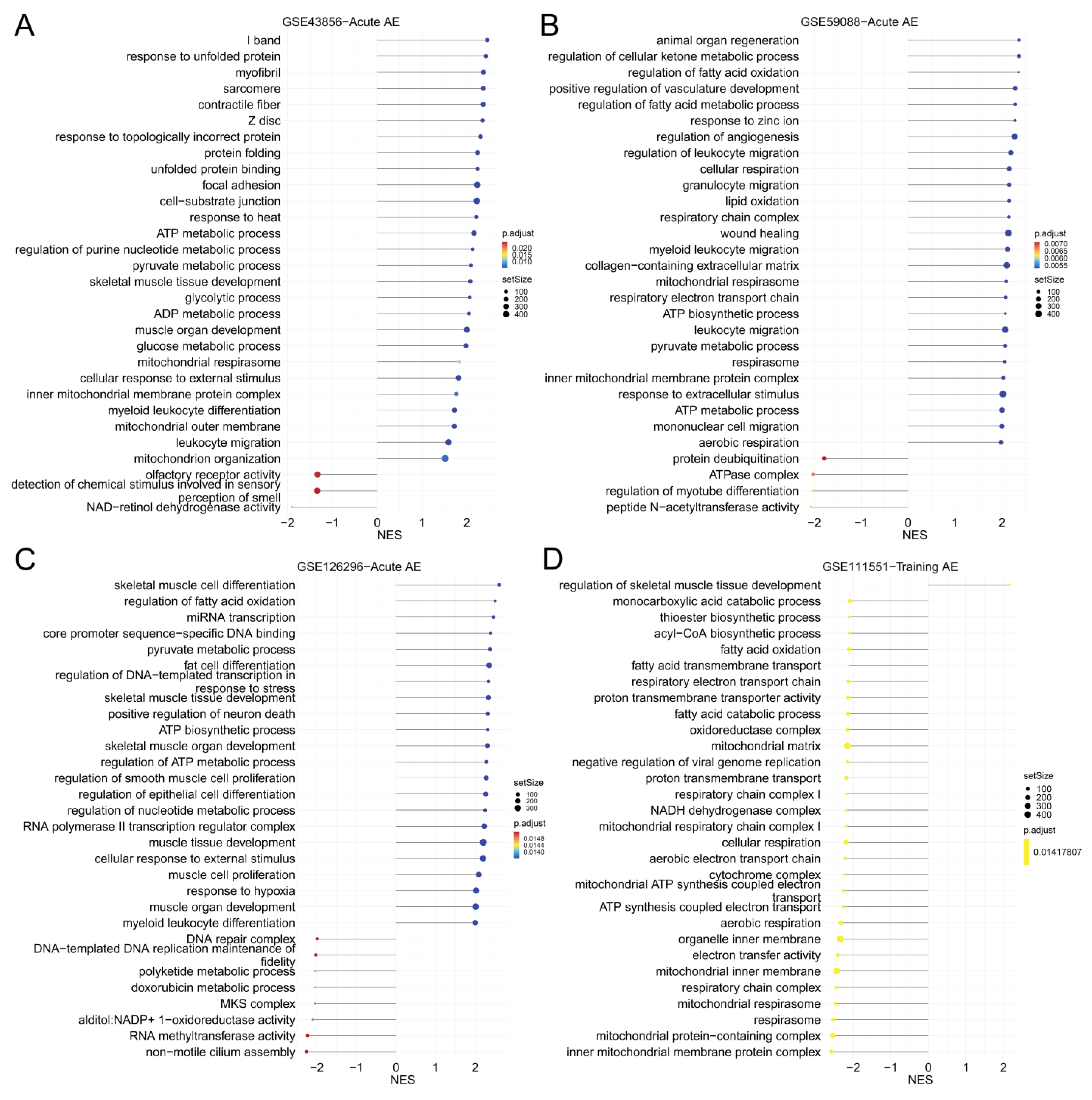
Figure 5.
The top 5 KEGG pathway-related gene sets from GSEA in each dataset of AE in the younger male. (A) GSE43856—acute AE; (B) GSE59088—acute AE; (C) GSE126296—acute AE; (D) GSE111551—training AE. Note: KEGG, Kyoto Encyclopedia of Genes and Genomes; GSEA, gene set enrichment analysis; AE, aerobic exercise.
Figure 5.
The top 5 KEGG pathway-related gene sets from GSEA in each dataset of AE in the younger male. (A) GSE43856—acute AE; (B) GSE59088—acute AE; (C) GSE126296—acute AE; (D) GSE111551—training AE. Note: KEGG, Kyoto Encyclopedia of Genes and Genomes; GSEA, gene set enrichment analysis; AE, aerobic exercise.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The characteristics of the studies included in the transcriptome analysis.
| Datasets | Groups | Intervention Details | Age (Year) | Sex (n) | Muscle Biopsy | Platform |
|---|---|---|---|---|---|---|
| Acute aerobic exercise (n = 28, included 28 males) | ||||||
| GSE43856 Neubauer O 2013, Austria [50] | Pre- and post-exercise | 1 h of cycling followed by 1 h of running. | 25 ± 4.1 | M (8) | NR (3 h, 48 h, 96 h) | GPL10558; Illumina Human HT-12 V4.0 expression beadchip |
| GSE59088 Kristian Vissing 2014, Denmark [51] | Endurance group and control pre-, 2.5 h, 5 h | 2 h of bicycle exercise at 60% post-training VO2 peak. | 23.4 ± 0.8 | M (6) | Vastus lateralis biopsy | GPL6244; [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| GSE126296 Rundqvist HC 2019, Sweden [52] | Pre- and post-exercise | Three all-out cycle sprints interspersed by 1/3-h recovery. | 26 ± 4 | M (7) | Vastus lateralis biopsy (7/3 h after the third sprint) | GPL6244; [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| GSE59363 Hansen JS 2015, Germany [53] | Pre- and post-exercise | A 1-h bicycle ergometer exercise | 57.3 ± 1.7 | M (7) | NR (0 h, 1 h, 3 h) | GPL6244; [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| Training aerobic exercise (n = 45, included 26 males and 19 females) | ||||||
| GSE111551 Michelle Silva, 2018, Brazil | Pre- and post-training | 18 weeks of running (3 times/week; 1 h every time). | Two groups group1, 26 ± 4; group2, 27 ± 3 | M (13) | NR | GPL17586; [HTA-2_0] Affymetrix Human Transcriptome Array 2.0 [transcript (gene) version] |
| GSE72462 Böhm A, 2016, Germany [54] | Pre- and post-training | The training program lasted 8 weeks and consisted of three supervised exercise sessions per week. Each training session consisted of 0.5 h of bicycle ergometer exercise and 0.5 h walking on a treadmill. | 46.4 ± 11 | M (7) F (13) | Muscle biopsies were taken from the lateral portion of the vastus lateralis of the quadriceps femoris after local anesthesia | GPL17586; [HTA-2_0] Affymetrix Human Transcriptome Array 2.0 [transcript (gene) version] |
| GSE1786 Radom-Aizik S, 2005, USA [55] | Pre- and post-training | 6 healthy sedentary men were trained on a cycle ergometer 3/wk for 12 wk at 80% of the predetermined maximal heart rate. | 68 ± 2.7 | M (6) | Vastus Lateralis muscle | GPL96; [HG-U133A] Affymetrix Human Genome U133A Array |
| GSE43760 Poelkens F, 2013, The Netherlands [56] | Pre- and post-training | 6 months guided endurance exercise training program | 49 ± 11 | F (6) | Vastus lateralis muscle | GPL6244; [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| Protein intake (n = 48, included 27 males and 21 females) | ||||||
| GSE8441 Thalacker-Mercer AE, 2007, USA [57] | Adequate protein vs. inadequate protein group | 11 subjects consumed 1.2 g/kg/d protein for a week first, then consumed 0.5 g/kg/d protein for the second week. | 55–80 (67 ± 7) | M (5) F (6) | Vastus lateralis biopsy | GPL96; [HG-U133A] Affymetrix Human Genome U133A Array |
| GSE9419 Thalacker-Mercer AE, 2008, USA [58] | Three groups with protein intakes of 0.50 g/kg/d, 0.75 g/kg/d, and 1.00 g/kg/d. | 22 healthy young and old males treated with protein intakes of 0.50 g/kg/d, 0.75 g/kg/d, and 1.00 g/kg/d. | Young 21–43 Elderly 63–79 | M (22) | Vastus lateralis biopsy | GPL570; [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array |
| GSE73525 Smith GI, 2016, Japan [59] | High-protein group vs. low-protein group | 9 subjects in a high protein (1.2 g/kg/d) group and 6 subjects in a normal protein (0.8 g/kg/d) group | 50–65 (58 ± 1) | F (15) | NR | GPL6244; [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| Acute aerobic exercise + protein intake (n = 20, included 20 males) | ||||||
| GSE27285 Rowlands DS, 2011, Switzerland [60] | Acute aerobic exercise with or without protein intake (iso-calorie) | Group1: 1 h intense cycling + 0.2/1.2/0.4 g/kg (fat/carbohydrate/protein). Group2: 1 h intense cycling + 0.2/1.6 g/kg (fat/carbohydrate). | 32.8 ± 6.4 | M (8) | Quadriceps at rest, at 3 h and at 48 h after exercise training | GPL6104; Illumina humanRef-8 v2.0 expression beadchip |
| GSE44818 Rowlands DS, 2015, Switzerland [61] | High-intensity cycling without protein intake; high-intensity cycling + 70 g protein; high-intensity cycling + 23 g protein | 5/3 h of high-intensity cycling with 70/15/180/30 g (protein/leucine/carbohydrate/fat); or with 23/5/180/30 g; or with 0/0/274/30 g | Mean age 30 years | M (12) | Vastus lateralis biopsy | GPL6947; Illumina HumanHT-12 V3.0 expression beadchip |
| Training aerobic exercise + protein intake (n = 40, included 40 males) | ||||||
| GSE147494 Knuiman P, 2020, UK [62] | Protein-treated group vs. control group | 12 weeks endurance training program with 30 g protein drink; or with 30 g carbohydrate drink (isocaloric) | 18–30 Protein group 21.5; Controls group 22.5 | M (40) | Vastus lateralis biopsy | GPL28236; [HuGene-2_1-st] Affymetrix Human Gene 2.1 ST Array |
Notes: M, male; F, female; VO2, oxygen uptake per unit of time; GSE, array accession from the Gene Expression Omnibus; NR, not reported, GPL, Gene Platform. VO2 peak: peak oxygen uptake per unit of time.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zeng, X.; Li, L.; Xia, Z.; Zou, L.; Kwok, T.; Su, Y. Transcriptomic Analysis of Human Skeletal Muscle in Response to Aerobic Exercise and Protein Intake. Nutrients 2023, 15, 3485. https://doi.org/10.3390/nu15153485
AMA Style
Zeng X, Li L, Xia Z, Zou L, Kwok T, Su Y. Transcriptomic Analysis of Human Skeletal Muscle in Response to Aerobic Exercise and Protein Intake. Nutrients. 2023; 15(15):3485. https://doi.org/10.3390/nu15153485
Chicago/Turabian StyleZeng, Xueqing, Linghong Li, Zhilin Xia, Lianhong Zou, Timothy Kwok, and Yi Su. 2023. "Transcriptomic Analysis of Human Skeletal Muscle in Response to Aerobic Exercise and Protein Intake" Nutrients 15, no. 15: 3485. https://doi.org/10.3390/nu15153485
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.