
Modification of Breakfast Fat Composition Can Modulate Cytokine and Other Inflammatory Mediators in Women: A Randomized Crossover Trial



Abstract
:1. Introduction
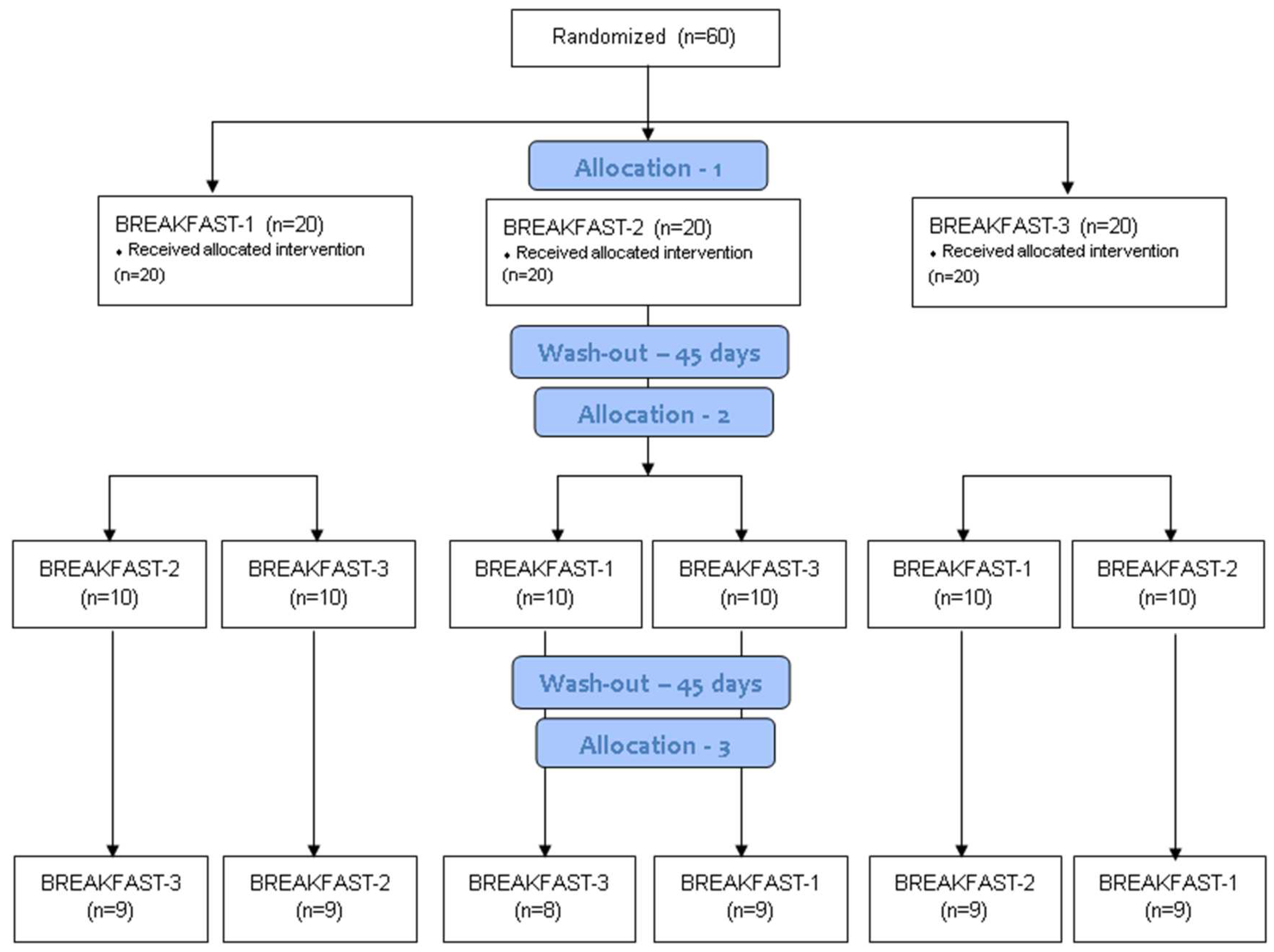
2. Materials and Methods
2.1. Design
2.2. Participants
2.3. Intervention
2.4. Measurements
2.5. Statistical Analysis
3. Results
3.1. Baseline Characteristics of the Participants
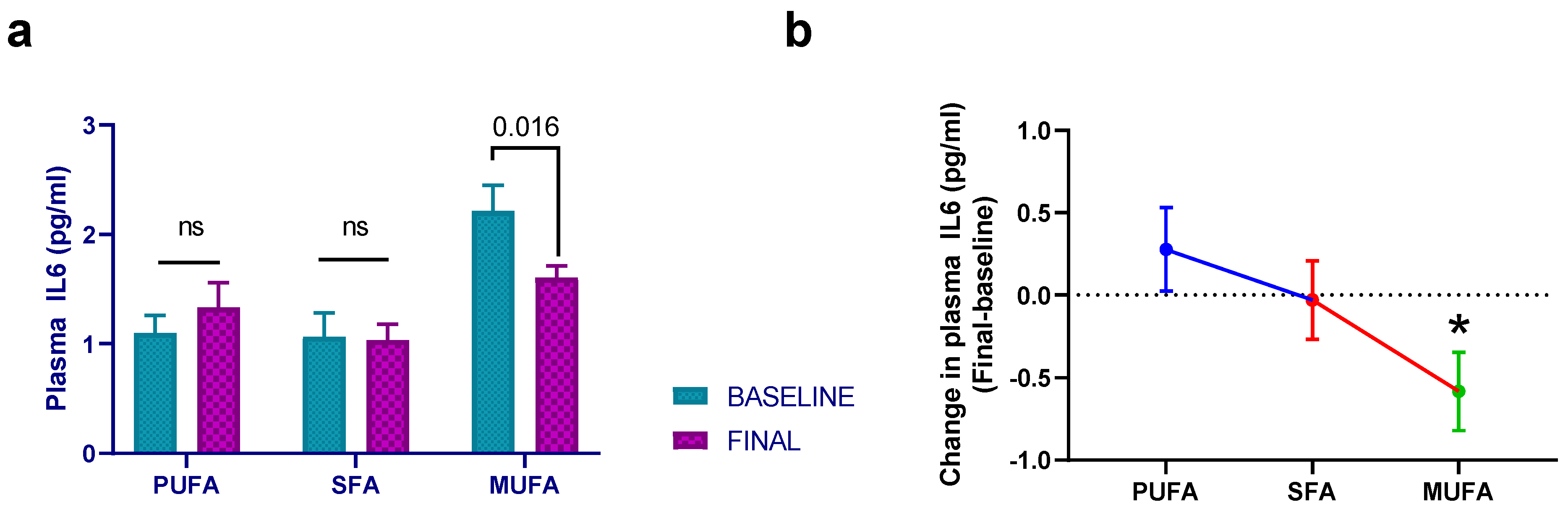
3.2. Effect of Breakfast Fat Composition on Interleukin Synthesis
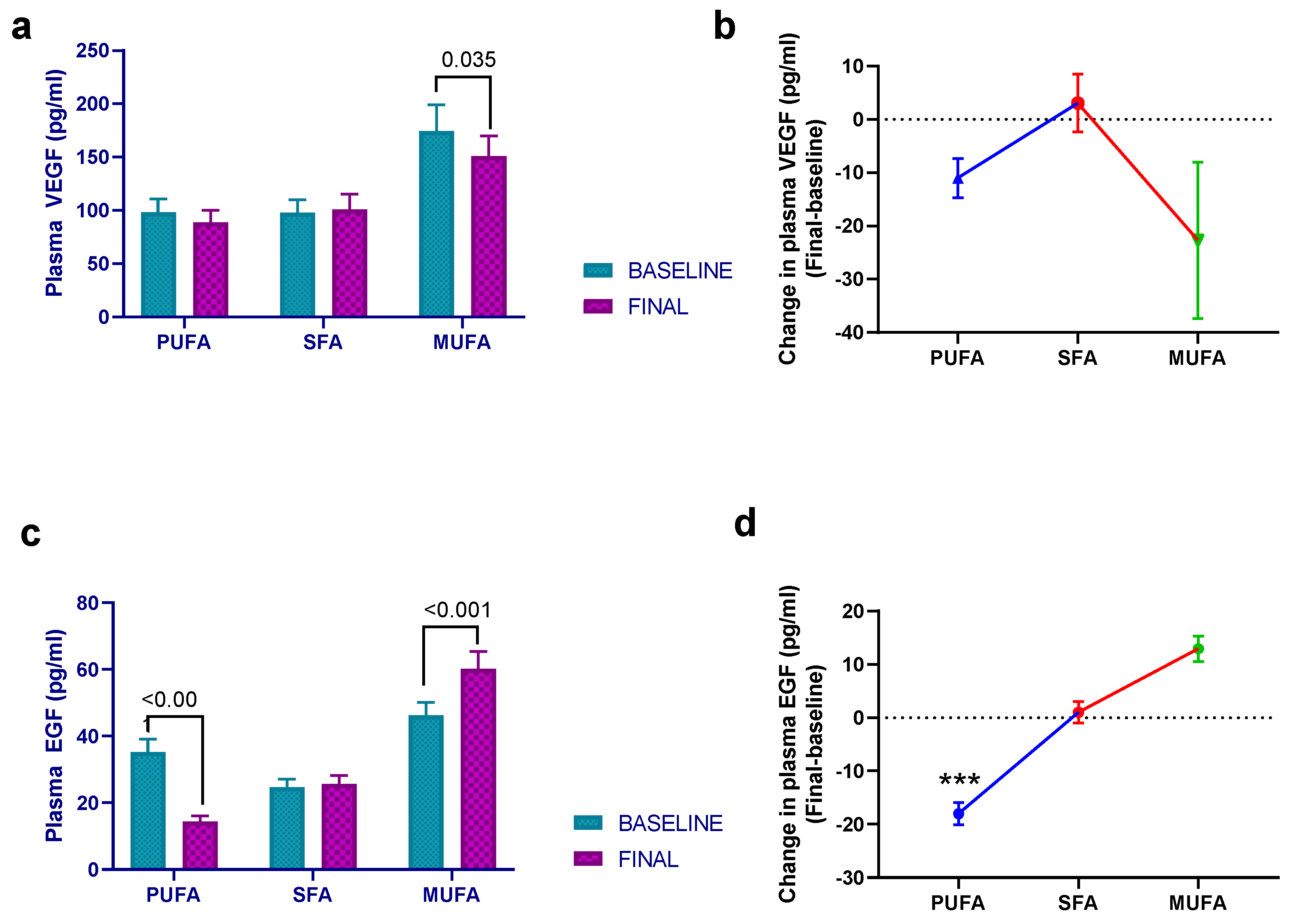
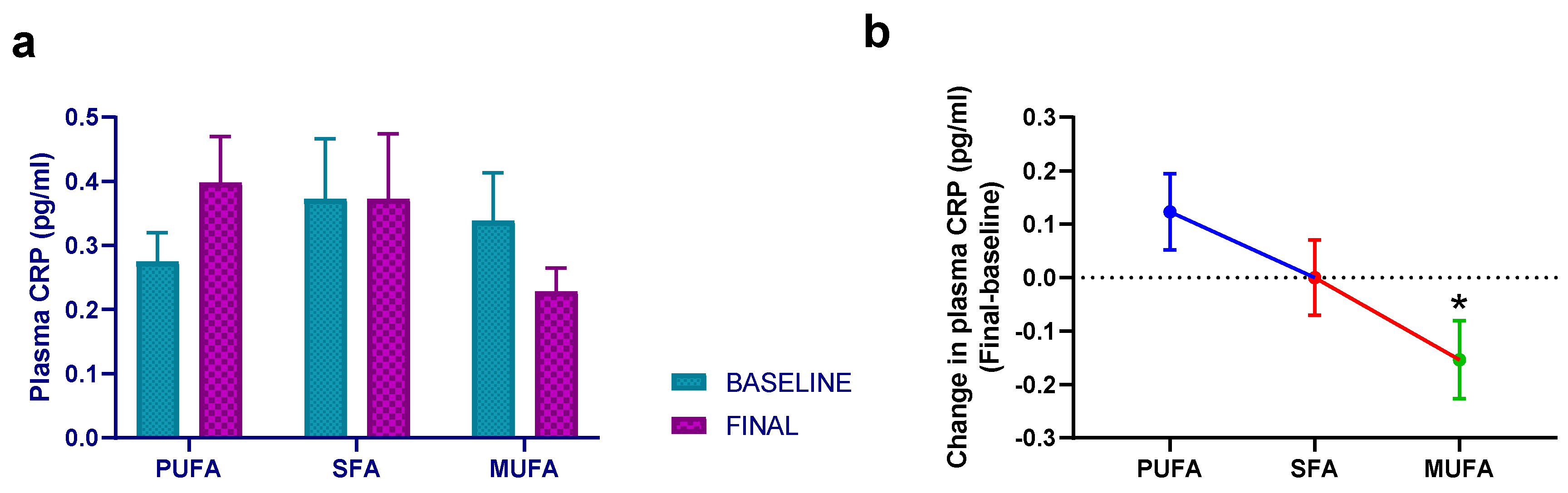
3.3. Effect of Breakfast Fat Composition on Other Cytokines and Inflammation Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franks, P.W. Obesity, Inflammatory Markers and Cardiovascular Disease: Distinguishing Causality from Confounding. J. Hum. Hypertens. 2006, 20, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a Link between Obesity, Metabolic Syndrome and Type 2 Diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory Mechanisms Linking Obesity and Metabolic Disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-Grade Inflammation, Diet Composition and Health: Current Research Evidence and Its Translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Durkin, L.A.; Childs, C.E.; Calder, P.C. Omega-3 Polyunsaturated Fatty Acids and the Intestinal Epithelium—A Review. Foods 2021, 10, 199. [Google Scholar] [CrossRef] [PubMed]
- Cantero, I.; Abete, I.; Babio, N.; Arós, F.; Corella, D.; Estruch, R.; Fitó, M.; Hebert, J.R.; Martínez-González, M.Á.; Pintó, X.; et al. Dietary Inflammatory Index and Liver Status in Subjects with Different Adiposity Levels within the PREDIMED Trial. Clin. Nutr. 2018, 37, 1736–1743. [Google Scholar] [CrossRef]
- Estruch, R.; Martínez-González, M.A.; Corella, D.; Salas-Salvadó, J.; Ruiz-Gutiérrez, V.; Covas, M.I.; Fiol, M.; Gómez-Gracia, E.; López-Sabater, M.C.; Vinyoles, E.; et al. Effects of a Mediterranean-Style Diet on Cardiovascular Risk Factors: A Randomized Trial. Ann. Intern. Med. 2006, 145, 1–11. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; Simonetta, I.; Daidone, M.; Mogavero, A.; Ortello, A.; Pinto, A. Metabolic and Vascular Effect of the Mediterranean Diet. Int. J. Mol. Sci. 2019, 20, 4716. [Google Scholar] [CrossRef]
- Zhou, H.; Urso, C.J.; Jadeja, V. Saturated Fatty Acids in Obesity-Associated Inflammation. J. Inflamm. Res. 2020, 13, 1–14. [Google Scholar] [CrossRef]
- Masson, C.J.; Mensink, R.P. Exchanging Saturated Fatty Acids for (n-6) Polyunsaturated Fatty Acids in a Mixed Meal May Decrease Postprandial Lipemia and Markers of Inflammation and Endothelial Activity in Overweight Men. J. Nutr. 2011, 141, 816–821. [Google Scholar] [CrossRef]
- Tortosa-Caparrós, E.; Navas-Carrillo, D.; Marín, F.; Orenes-Piñero, E. Anti-Inflammatory Effects of Omega 3 and Omega 6 Polyunsaturated Fatty Acids in Cardiovascular Disease and Metabolic Syndrome. Crit. Rev. Food Sci. Nutr. 2017, 57, 3421–3429. [Google Scholar] [CrossRef] [PubMed]
- Galindo Muñoz, J.S.; Gómez Gallego, M.; Díaz Soler, I.; Barberá Ortega, M.C.; Martínez Cáceres, C.M.; Hernández Morante, J.J. Effect of a Chronotype-Adjusted Diet on Weight Loss Effectiveness: A Randomized Clinical Trial. Clin. Nutr. 2020, 39, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Patterson, R.E.; Ang, A.; Emond, J.A.; Shetty, N.; Arab, L. Timing of Energy Intake during the Day Is Associated with the Risk of Obesity in Adults. J. Hum. Nutr. Diet. 2014, 27, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Gomez-Abellan, P.; Alburquerque-Bejar, J.J.; Lee, Y.-C.; Ordovas, J.M.; Scheer, F.A.J.L.; Gómez-Abellán, P.; Alburquerque-Béjar, J.J.; Lee, Y.-C.; Ordovás, J.M.; et al. Timing of Food Intake Predicts Weight Loss Effectiveness. Int. J. Obes. 2013, 37, 604–611. [Google Scholar] [CrossRef]
- van der Merwe, C.; Münch, M.; Kruger, R. Chronotype Differences in Body Composition, Dietary Intake and Eating Behavior Outcomes: A Scoping Systematic Review. Adv. Nutr. 2022, 13, 2357. [Google Scholar] [CrossRef]
- Yong, Y.N.; Dong, J.; Pakkiri, L.S.; Henry, C.J.; Haldar, S.; Drum, C.L. Chronometabolism: The Timing of the Consumption of Meals Has a Greater Influence Than Glycemic Index (GI) on the Postprandial Metabolome. Metabolites 2023, 13, 490. [Google Scholar] [CrossRef]
- Shimizu, H.; Hanzawa, F.; Kim, D.; Sun, S.; Laurent, T.; Umeki, M.; Ikeda, S.; Mochizuki, S.; Oda, H. Delayed First Active-Phase Meal, a Breakfast-Skipping Model, Led to Increased Body Weight and Shifted the Circadian Oscillation of the Hepatic Clock and Lipid Metabolism-Related Genes in Rats Fed a High-Fat Diet. PLoS ONE 2018, 13, e0206669. [Google Scholar] [CrossRef]
- Jakubowicz, D.; Barnea, M.; Wainstein, J.; Froy, O. High Caloric Intake at Breakfast vs. Dinner Differentially Influences Weight Loss of Overweight and Obese Women. Obesity 2013, 21, 2504–2512. [Google Scholar] [CrossRef]
- Konishi, T.; Takahashi, Y.; Shiina, Y.; Oike, H.; Oishi, K. Time-of-Day Effects of Consumption of Fish Oil-Enriched Sausages on Serum Lipid Parameters and Fatty Acid Composition in Normolipidemic Adults: A Randomized, Double-Blind, Placebo-Controlled, and Parallel-Group Pilot Study. Nutrition 2021, 90, 111247. [Google Scholar] [CrossRef]
- Shinto, T.; Makino, S.; Tahara, Y.; Nitta, L.; Kuwahara, M.; Tada, A.; Abe, N.; Michie, M.; Shibata, S. Relationship Between Protein Intake in Each Traditional Meal and Physical Activity: Cross-Sectional Study. JMIR Public Health Surveill. 2022, 8, e35898. [Google Scholar] [CrossRef]
- Truman, S.C.; Wirth, M.D.; Arp Adams, S.; Turner-McGrievy, G.M.; Reiss, K.E.; Hébert, J.R. Meal Timing, Distribution of Macronutrients, and Inflammation among African-American Women: A Cross-Sectional Study. Chronobiol. Int. 2022, 39, 976–983. [Google Scholar] [CrossRef]
- Lasserre, M.; Mendy, F.; Spielmann, D.; Jacotot, B. Effects of Different Dietary Intake of Essential Fatty Acids on C20:3ω6 and C20:4ω6 Serum Levels in Human Adults. Lipids 1985, 20, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Mayr, S.; Buchner, A.; Erdfelder, E.; Faul, F. A Short Tutorial of GPower. Tutor. Quant. Methods Psychol. 2007, 3, 51–59. [Google Scholar] [CrossRef]
- Delgado-Alarcón, J.M.; Morante, J.J.H.; Aviles, F.V.; Albaladejo-Otón, M.D.; Morillas-Ruíz, J.M. Effect of the Fat Eaten at Breakfast on Lipid Metabolism: A Crossover Trial in Women with Cardiovascular Risk. Nutrients 2020, 12, 1695. [Google Scholar] [CrossRef] [PubMed]
- Han, S.N.; Leka, L.S.; Lichtenstein, A.H.; Ausman, L.M.; Schaefer, E.J.; Meydani, S.N. Effect of Hydrogenated and Saturated, Relative to Polyunsaturated, Fat on Immune and Inflammatory Responses of Adults with Moderate Hypercholesterolemia. J. Lipid Res. 2002, 43, 445–452. [Google Scholar] [CrossRef]
- Cruz-Teno, C.; Pérez-Martínez, P.; Delgado-Lista, J.; Yubero-Serrano, E.M.; García-Ríos, A.; Marín, C.; Gómez, P.; Jiménez-Gómez, Y.; Camargo, A.; Rodríguez-Cantalejo, F.; et al. Dietary Fat Modifies the Postprandial Inflammatory State in Subjects with Metabolic Syndrome: The LIPGENE Study. Mol. Nutr. Food Res. 2012, 56, 854–865. [Google Scholar] [CrossRef]
- Poppitt, S.D.; Keogh, G.F.; Lithander, F.E.; Wang, Y.; Mulvey, T.B.; Chan, Y.K.; McArdle, B.H.; Cooper, G.J.S. Postprandial Response of Adiponectin, Interleukin-6, Tumor Necrosis Factor-α, and C-Reactive Protein to a High-Fat Dietary Load. Nutrition 2008, 24, 322–329. [Google Scholar] [CrossRef]
- Schubert, R.; Kitz, R.; Beermann, C.; Rose, M.A.; Baer, P.C.; Zielen, S.; Boehles, H. Influence of Low-Dose Polyunsaturated Fatty Acids Supplementation on the Inflammatory Response of Healthy Adults. Nutrition 2007, 23, 724–730. [Google Scholar] [CrossRef]
- Schüler, R.; Seebeck, N.; Osterhoff, M.A.; Witte, V.; Flöel, A.; Busjahn, A.; Jais, A.; Brüning, J.C.; Frahnow, T.; Kabisch, S.; et al. VEGF and GLUT1 Are Highly Heritable, Inversely Correlated and Affected by Dietary Fat Intake: Consequences for Cognitive Function in Humans. Mol. Metab. 2018, 11, 129–136. [Google Scholar] [CrossRef]
- Ridker, P.M.; Rifai, N.; Stampfer, M.J.; Hennekens, C.H. Plasma Concentration of Interleukin-6 and the Risk of Future Myocardial Infarction among Apparently Healthy Men. Circulation 2000, 101, 1767–1772. [Google Scholar] [CrossRef]
- Liu, W.; Yalcinkaya, M.; Maestre, I.F.; Olszewska, M.; Ampomah, P.B.; Heimlich, J.B.; Wang, R.; Vela, P.S.; Xiao, T.; Bick, A.G.; et al. Blockade of IL-6 Signaling Alleviates Atherosclerosis in Tet2-Deficient Clonal Hematopoiesis. Nat. Cardiovasc. Res. 2023, 2, 572. [Google Scholar] [CrossRef] [PubMed]
- Kaptoge, S.; Di Angelantonio, E.; Lowe, G.; Pepys, M.B.; Thompson, S.G.; Collins, R.; Danesh, J.; Tipping, R.W.; Ford, C.E.; Pressel, S.L.; et al. C-Reactive Protein Concentration and Risk of Coronary Heart Disease, Stroke, and Mortality: An Individual Participant Meta-Analysis. Lancet 2010, 375, 132. [Google Scholar] [CrossRef] [PubMed]
- Cagnina, A.; Chabot, O.; Davin, L.; Lempereur, M.; Maréchal, P.; Oury, C.; Lancellotti, P. Atherosclerosis--an Inflammatory Disease. N. Engl. J. Med. 1999, 340, 302–309. [Google Scholar] [CrossRef]
- Estruch, R. Anti-Inflammatory Effects of the Mediterranean Diet: The Experience of the PREDIMED Study. Proc. Nutr. Soc. 2010, 69, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Dell’Agli, M.; Fagnani, R.; Mitro, N.; Scurati, S.; Masciadri, M.; Mussoni, L.; Galli, G.V.; Bosisio, E.; Crestani, M.; De Fabiani, E.; et al. Minor Components of Olive Oil Modulate Proatherogenic Adhesion Molecules Involved in Endothelial Activation. J. Agric. Food Chem. 2006, 54, 3259–3264. [Google Scholar] [CrossRef]
- Teng, K.T.; Chang, L.F.; Vethakkan, S.R.; Nesaretnam, K.; Sanders, T.A.B. Effects of Exchanging Carbohydrate or Monounsaturated Fat with Saturated Fat on Inflammatory and Thrombogenic Responses in Subjects with Abdominal Obesity: A Randomized Controlled Trial. Clin. Nutr. 2017, 36, 1250–1258. [Google Scholar] [CrossRef]
- Ryan, M.F.; Grada, C.O.; Morris, C.; Segurado, R.; Walsh, M.C.; Gibney, E.R.; Brennan, L.; Roche, H.M.; Gibney, M.J. Within-Person Variation in the Postprandial Lipemic Response of Healthy Adults. Am. J. Clin. Nutr. 2013, 97, 261–267. [Google Scholar] [CrossRef]
- Meneses, M.E.; Camargo, A.; Perez-Martinez, P.; Delgado-Lista, J.; Cruz-Teno, C.; Jimenez-Gomez, Y.; Paniagua, J.A.; Gutierrez-Mariscal, F.M.; Tinahones, F.J.; Vidal-Puig, A.; et al. Postprandial Inflammatory Response in Adipose Tissue of Patients with Metabolic Syndrome after the Intake of Different Dietary Models. Mol. Nutr. Food Res. 2011, 55, 1759–1770. [Google Scholar] [CrossRef]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef]
- Botham, K.M.; Wheeler-Jones, C.P.D. Postprandial Lipoproteins and the Molecular Regulation of Vascular Homeostasis. Prog. Lipid Res. 2013, 52, 446–464. [Google Scholar] [CrossRef]
- Ramadeen, A.; Laurent, G.; dos Santos, C.C.; Hu, X.; Connelly, K.A.; Holub, B.J.; Mangat, I.; Dorian, P. N-3 Polyunsaturated Fatty Acids Alter Expression of Fibrotic and Hypertrophic Genes in a Dog Model of Atrial Cardiomyopathy. Hear. Rhythm 2010, 7, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.W.L.; Seo, J.; Davidson, L.A.; Callaway, E.S.; Fan, Y.-Y.; Lupton, J.R.; Chapkin, R.S. N-3 PUFA Alter Caveolae Lipid Composition and Resident Protein Localization in Mouse Colon. FASEB J. 2004, 18, 1040–1042. [Google Scholar] [CrossRef]
- Hernández, E.A.; Kahl, S.; Seelig, A.; Begovatz, P.; Irmler, M.; Kupriyanova, Y.; Nowotny, B.; Nowotny, P.; Herder, C.; Barosa, C.; et al. Acute Dietary Fat Intake Initiates Alterations in Energy Metabolism and Insulin Resistance. J. Clin. Invest. 2017, 127, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.; Seyedsadjadi, N.; Grant, R. Saturated Fatty Acid Intake Is Associated With Increased Inflammation, Conversion of Kynurenine to Tryptophan, and Delta-9 Desaturase Activity in Healthy Humans. Int. J. Tryptophan Res. 2020, 13. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics (n = 53) | |
|---|---|
| Age (years) | 63 ± 3 (58–68) |
| Weight (kg) | 64.6 ± 9.0 (62.1–67.2) |
| BMI | 27.77 ± 4.01 (26.63–28.91) |
| SBP (mmHg) | 130 ± 22 (124–136) |
| DBP (mmHg) | 72 ± 9 (69–75) |
| Hypertension History (n; %) | 16; 31.4% |
| Diabetes History (n; %) | 7; 13.7% |
| Hypercholesterolemia History (n; %) | 19; 37.3% |
| Family History of CVD (n; %) | 17; 33.3% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Alarcón, J.M.; Hernández Morante, J.J.; Morillas-Ruiz, J.M. Modification of Breakfast Fat Composition Can Modulate Cytokine and Other Inflammatory Mediators in Women: A Randomized Crossover Trial. Nutrients 2023, 15, 3711. https://doi.org/10.3390/nu15173711
Delgado-Alarcón JM, Hernández Morante JJ, Morillas-Ruiz JM. Modification of Breakfast Fat Composition Can Modulate Cytokine and Other Inflammatory Mediators in Women: A Randomized Crossover Trial. Nutrients. 2023; 15(17):3711. https://doi.org/10.3390/nu15173711
Chicago/Turabian StyleDelgado-Alarcón, Jessica M., Juan José Hernández Morante, and Juana M. Morillas-Ruiz. 2023. "Modification of Breakfast Fat Composition Can Modulate Cytokine and Other Inflammatory Mediators in Women: A Randomized Crossover Trial" Nutrients 15, no. 17: 3711. https://doi.org/10.3390/nu15173711
APA StyleDelgado-Alarcón, J. M., Hernández Morante, J. J., & Morillas-Ruiz, J. M. (2023). Modification of Breakfast Fat Composition Can Modulate Cytokine and Other Inflammatory Mediators in Women: A Randomized Crossover Trial. Nutrients, 15(17), 3711. https://doi.org/10.3390/nu15173711